環(huán)境劑量磷酸三(1,3-二氯異丙基)酯多代暴露對(duì)斑馬魚(yú)子代仔魚(yú)的神經(jīng)發(fā)育毒性
2020-07-02 14:30:14丁希勝馬徐發(fā)余麗琴
生態(tài)毒理學(xué)報(bào) 2020年2期
關(guān)鍵詞:劑量
丁希勝,馬徐發(fā),2,#,余麗琴,2,*
1. 華中農(nóng)業(yè)大學(xué)水產(chǎn)學(xué)院,武漢 430070 2. 湖北省池塘養(yǎng)殖工程實(shí)驗(yàn)室,武漢 430070
有機(jī)磷阻燃劑被廣泛添加在電子設(shè)備、紡織品和家具等日常用品中,用于避免易燃物被引燃和延遲火災(zāi)的蔓延[1-2]。作為主要的有機(jī)磷阻燃劑,磷酸三(1,3-二氯異丙基)酯(TDCIPP)通常添加在家具、兒童泡沫玩具和汽車(chē)裝潢物中[3]。作為一種新型的環(huán)境污染物,在地表水、自來(lái)水、室內(nèi)空氣和水生生物體中,甚至人體代謝物中都有一定濃度的TDCIPP被檢出[4-5]。據(jù)報(bào)道,國(guó)內(nèi)松花江水樣中TDCIPP濃度范圍是2.5~40 ng·L-1[6-7]。在我國(guó)青島、廈門(mén)和連云港附近水域采集的水樣中,TDCIPP的最高濃度達(dá)377 ng·L-1[8]。我國(guó)南京市10個(gè)自來(lái)水廠(chǎng)水樣中TDCIPP的平均含量為8.5 ng·L-1[9]。在我國(guó)珠江三角洲區(qū)域生活的淡水魚(yú)肌肉中TDCIPP的蓄積量達(dá)到251 μg·kg-1[10]。
近年來(lái),大量的離體和活體實(shí)驗(yàn)研究都表明了TDCIPP具有神經(jīng)毒性。例如,離體實(shí)驗(yàn)結(jié)果表明,TDCIPP暴露降低了細(xì)胞的活力,提高了多種類(lèi)型神經(jīng)細(xì)胞的凋亡率[11-13],并干擾大鼠腎上腺嗜鉻細(xì)胞瘤細(xì)胞(PC12)的細(xì)胞分化和細(xì)胞遷移[13]。一項(xiàng)活體研究表明,TDCIPP暴露導(dǎo)致稀有鮈鯽(Gobiocyprisrarus)成魚(yú)體內(nèi)的神經(jīng)營(yíng)養(yǎng)因子及其受體的基因表達(dá)顯著性降低[14]。在斑馬魚(yú)中,TDCIPP暴露可降低神經(jīng)遞質(zhì)(例如血清素和多巴胺)的水平,下調(diào)成魚(yú)和母體暴露后產(chǎn)下的子代仔魚(yú)(受精后5 d,5-day post-fertilization,5 dpf)中神經(jīng)發(fā)育相關(guān)基因的表達(dá),包括髓鞘堿性蛋白(myelin basic protein,mbp)和突觸素Ⅱ(synapsin Ⅱa,syn2a)[15-16]。早期生命階段的研究表明,TDCIPP急性暴露會(huì)引起魚(yú)類(lèi)顯著的行為改變,特別是仔魚(yú)游泳活動(dòng)[17-21],推測(cè)魚(yú)類(lèi)游泳行為的變化可能是TDCIPP暴露的敏感終點(diǎn)。但是,這些研究均是基于較高劑量的暴露實(shí)驗(yàn),有關(guān)環(huán)境劑量TDCIPP對(duì)魚(yú)類(lèi)游泳行為的影響尚未知。此外,生物體在自然生態(tài)系統(tǒng)中往往長(zhǎng)達(dá)多代都暴露于環(huán)境污染物中。所以,有必要研究環(huán)境劑量TDCIPP多代暴露對(duì)斑馬魚(yú)子代仔魚(yú)神經(jīng)發(fā)育的影響。
在斑馬魚(yú)中,一些與胚胎/仔魚(yú)神經(jīng)發(fā)育相關(guān)的候選基因可以作為評(píng)價(jià)化合物神經(jīng)發(fā)育毒性的生物標(biāo)志物[22]。其中,神經(jīng)元特異性RNA結(jié)合蛋白(ELAV like neuron-specific RNA binding protein 3,elavl3)和神經(jīng)原素1 (neurogenin-related gene 1,ngn1)是神經(jīng)元發(fā)育的標(biāo)志基因[23-24]。α1微管蛋白(α1-tubulin)基因在神經(jīng)元軸突和樹(shù)突的發(fā)育和再生中起著非常重要的作用[25],生長(zhǎng)相關(guān)蛋白(growth associated protein 43,gap43)基因在神經(jīng)元中軸突再生和發(fā)育時(shí)高表達(dá)[26],神經(jīng)生長(zhǎng)因子(netrins)基因可以刺激斑馬魚(yú)胚胎中神經(jīng)元軸突的生長(zhǎng)[27],zn5是第二運(yùn)動(dòng)神經(jīng)元軸突的標(biāo)志基因[28-29]。音猬基因(shha)的功能主要是連接視網(wǎng)膜神經(jīng)調(diào)節(jié)細(xì)胞與脊髓軸突間的軸突索[30-31]。膠質(zhì)纖維酸性蛋白(glial fibrillary acidic protein, GFAP)是一種中間絲蛋白,在中樞神經(jīng)系統(tǒng)的星形膠質(zhì)細(xì)胞和放射狀神經(jīng)膠質(zhì)細(xì)胞中高度表達(dá),被作為星形膠質(zhì)細(xì)胞的標(biāo)志物[32]。髓鞘堿性蛋白(myelin basic protein, MBP)是斑馬魚(yú)正在發(fā)育的中樞神經(jīng)系統(tǒng)中軸突的髓鞘形成所必需的[33]。
本文以斑馬魚(yú)為研究模型,將3代斑馬魚(yú)暴露于環(huán)境劑量的TDCIPP,評(píng)價(jià)每一代的子代仔魚(yú)生長(zhǎng)發(fā)育以及運(yùn)動(dòng)行為所受到的影響,并考察神經(jīng)發(fā)育標(biāo)志基因的表達(dá)變化,分析其與行為變化的相關(guān)性,探討影響行為的可能原因,評(píng)價(jià)TDCIPP多代暴露對(duì)其子代仔魚(yú)神經(jīng)發(fā)育的毒性效應(yīng),研究結(jié)果對(duì)全面評(píng)價(jià)TDCIPP的環(huán)境和人類(lèi)健康風(fēng)險(xiǎn)具有重要的參考意義。
1 材料與方法(Materials and methods)
1.1 實(shí)驗(yàn)試劑
磷酸三(1,3-二氯異丙基)酯(TDCIPP)純度>95%,購(gòu)自東京化成工業(yè)株式會(huì)社(東京,日本)。用于儲(chǔ)存和稀釋TDCIPP的二甲基亞砜(DMSO)純度>99%,間氨基苯甲酸乙酯甲磺酸鹽(MS-222)純度98%,二者均購(gòu)自Sigma-Aldrich公司(密西根州圣路易斯,美國(guó))。Trizol試劑、PrimeScript?RT reagent試劑盒和SYBR?Real-Time PCR Master購(gòu)自TaKaRa公司(大連,中國(guó))。氯仿、異戊醇和無(wú)水乙醇均為國(guó)產(chǎn)分析純。
1.2 斑馬魚(yú)飼養(yǎng)及TDCIPP暴露實(shí)驗(yàn)
實(shí)驗(yàn)所用AB純系斑馬魚(yú)的養(yǎng)殖以及暴露實(shí)驗(yàn)根據(jù)Yu等[34]提供的實(shí)驗(yàn)方案實(shí)施。斑馬魚(yú)胚胎(野生型AB品系)購(gòu)于中國(guó)科學(xué)院水生生物研究所。挑選發(fā)育正常的囊胚期胚胎(受精后2 h,2 hpf),隨機(jī)分配至含有0、3、30和300 ng·L-1TDCIPP暴露液的培養(yǎng)皿中,每個(gè)皿中放置100顆胚胎,每個(gè)濃度設(shè)置3個(gè)平行,所有暴露組DMSO濃度為0.001% (V/V)。將培養(yǎng)皿置于光照培養(yǎng)箱中培養(yǎng),5 d后將孵化出的仔魚(yú)從培養(yǎng)皿中轉(zhuǎn)移到魚(yú)房中25 L的魚(yú)缸內(nèi),每隔2天全部更換一次暴露液。控制水溫穩(wěn)定在(28±0.5) ℃,光照控制為14 h光照∶10 h黑暗,每日投喂2次豐年蟲(chóng)。實(shí)驗(yàn)中所投喂的豐年蟲(chóng)均于實(shí)驗(yàn)室中孵化,豐年蟲(chóng)卵從天津豐年水產(chǎn)養(yǎng)殖有限公司購(gòu)得。從F0胚胎開(kāi)始暴露,持續(xù)暴露3代(F0代、F1代和F2代),每一代暴露120 d,然后以1∶1的比例將雄魚(yú)和雌魚(yú)混在一起產(chǎn)卵,收集每一代的子代(F1代、F2代和F3代),于暴露液中持續(xù)培養(yǎng)至5 dpf,統(tǒng)計(jì)F1代、F2代和F3代胚胎/仔魚(yú)3 dpf的孵化率、5 dpf的存活率、5 dpf的畸形率和5 dpf的體長(zhǎng)。
1.3 斑馬魚(yú)仔魚(yú)運(yùn)動(dòng)行為分析
斑馬魚(yú)仔魚(yú)的游泳行為的檢測(cè)參照筆者之前的實(shí)驗(yàn)方法[20]。簡(jiǎn)單的說(shuō),針對(duì)每一代斑馬魚(yú)仔魚(yú)(F1代、F2代和F3代),每個(gè)濃度每個(gè)平行缸取8尾仔魚(yú)進(jìn)行游泳行為測(cè)定,采用ZebraLab行為圖像監(jiān)測(cè)器中Video-Track系統(tǒng)(View Point Life Sciences,蒙特利爾,加拿大)中的攝像頭記錄仔魚(yú)運(yùn)動(dòng)軌跡,從而分析獲得相關(guān)行為數(shù)據(jù),定量分析5 dpf仔魚(yú)的游泳行為。測(cè)定之前,先將斑馬魚(yú)放入裝有TDCIPP暴露液的12孔板中適應(yīng)10 min,調(diào)節(jié)水溫保持在(28±0.5) ℃,每個(gè)孔中一條仔魚(yú);然后進(jìn)行40 min的光暗交替(5 min光照—5 min黑暗—5 min光照—5 min黑暗)刺激,每30秒采集一次仔魚(yú)運(yùn)動(dòng)頻率、行進(jìn)距離和運(yùn)動(dòng)持續(xù)時(shí)間,試驗(yàn)中保持安靜的環(huán)境。記錄的數(shù)據(jù)用在線(xiàn)Open Office.Org 2.4軟件分析。
1.4 實(shí)時(shí)熒光定量PCR (qRT-PCR)
斑馬魚(yú)仔魚(yú)的神經(jīng)發(fā)育相關(guān)基因的檢測(cè)參照筆者之前的實(shí)驗(yàn)方法[35]。對(duì)F1代、F2代和F3代120 hpf的斑馬魚(yú)仔魚(yú)進(jìn)行神經(jīng)相關(guān)基因表達(dá)分析。每個(gè)平行缸隨機(jī)取50尾仔魚(yú)(n=3)置于Trizol中,根據(jù)筆者之前的方法[33]進(jìn)行總RNA的提取,采用分光光度計(jì)檢測(cè)260/280比值來(lái)確定cDNA的合成。使用在線(xiàn)引物設(shè)計(jì)軟件Primer 3 software (http://bioinfo.ut.ee/primer3-0.4.0/primer3/)進(jìn)行設(shè)計(jì),引物序列如表1所示。
1.5 數(shù)據(jù)分析方法
實(shí)驗(yàn)數(shù)據(jù)采用SPSS 13.0軟件(SPSS, Chicago, IL, USA)處理分析。首先采用Kolmogorov-Smirnov方法對(duì)數(shù)據(jù)進(jìn)行正態(tài)分布檢驗(yàn),對(duì)不符合正態(tài)分布的數(shù)據(jù)進(jìn)行轉(zhuǎn)換使其符合正態(tài)分布。然后采用Levene’檢驗(yàn)方法進(jìn)行數(shù)據(jù)的方差齊性檢驗(yàn)。暴露組和對(duì)照組采用單因素方差分析方法(one-way analysis of variance, ANOVA)中Tukey’s多重比較法進(jìn)行數(shù)據(jù)的差異性檢驗(yàn)。結(jié)果表示為3次獨(dú)立重復(fù)試驗(yàn)數(shù)據(jù)的算術(shù)平均值±標(biāo)準(zhǔn)差(mean±SD)。使用Spearman相關(guān)分析來(lái)檢驗(yàn)仔魚(yú)在光暗刺激下的平均游泳速度和神經(jīng)發(fā)育相關(guān)基因表達(dá)之間的相關(guān)性,P<0.05為顯著性差異。
2 結(jié)果(Results)
2.1 TDCIPP多代暴露對(duì)各自子代5 dpf仔魚(yú)存活率和孵化率的影響
如表2所示,與對(duì)照組相比,經(jīng)環(huán)境劑量的TDCIPP (300 ng·L-1)暴露后,F(xiàn)0代斑馬魚(yú)的子代F1代的孵化率顯著性降低(P<0.05),5 dpf子代的存活率也顯著性下降(P<0.01)(表2),但其畸形率和體長(zhǎng)無(wú)顯著性變化。隨著暴露代數(shù)的增加,與對(duì)照組相比,F(xiàn)2代和F3代仔魚(yú)的生長(zhǎng)、存活率以及畸形率均無(wú)顯著性差異(表2)。
2.2 TDCIPP多代暴露對(duì)各自子代5 dpf仔魚(yú)神經(jīng)行為的影響
如圖1所示,在光暗周期刺激下,與對(duì)照組相比,在初始黑暗狀態(tài)下300 ng·L-1TDCIPP暴露組中F0代斑馬魚(yú)120 d后所產(chǎn)子代F1代仔魚(yú)的游泳速度顯著性降低(P<0.05);在處于光照環(huán)境中時(shí),3和300 ng·L-1TDCIPP暴露組仔魚(yú)的游泳速度均顯著性下降(P<0.05、P<0.05);當(dāng)環(huán)境再次處于黑暗周期時(shí),最高劑量(300 ng·L-1)暴露導(dǎo)致子代仔魚(yú)的游泳速度顯著性下降(P<0.05)。

表1 研究中的目的基因與引物序列Table 1 Genes and primers used in this study
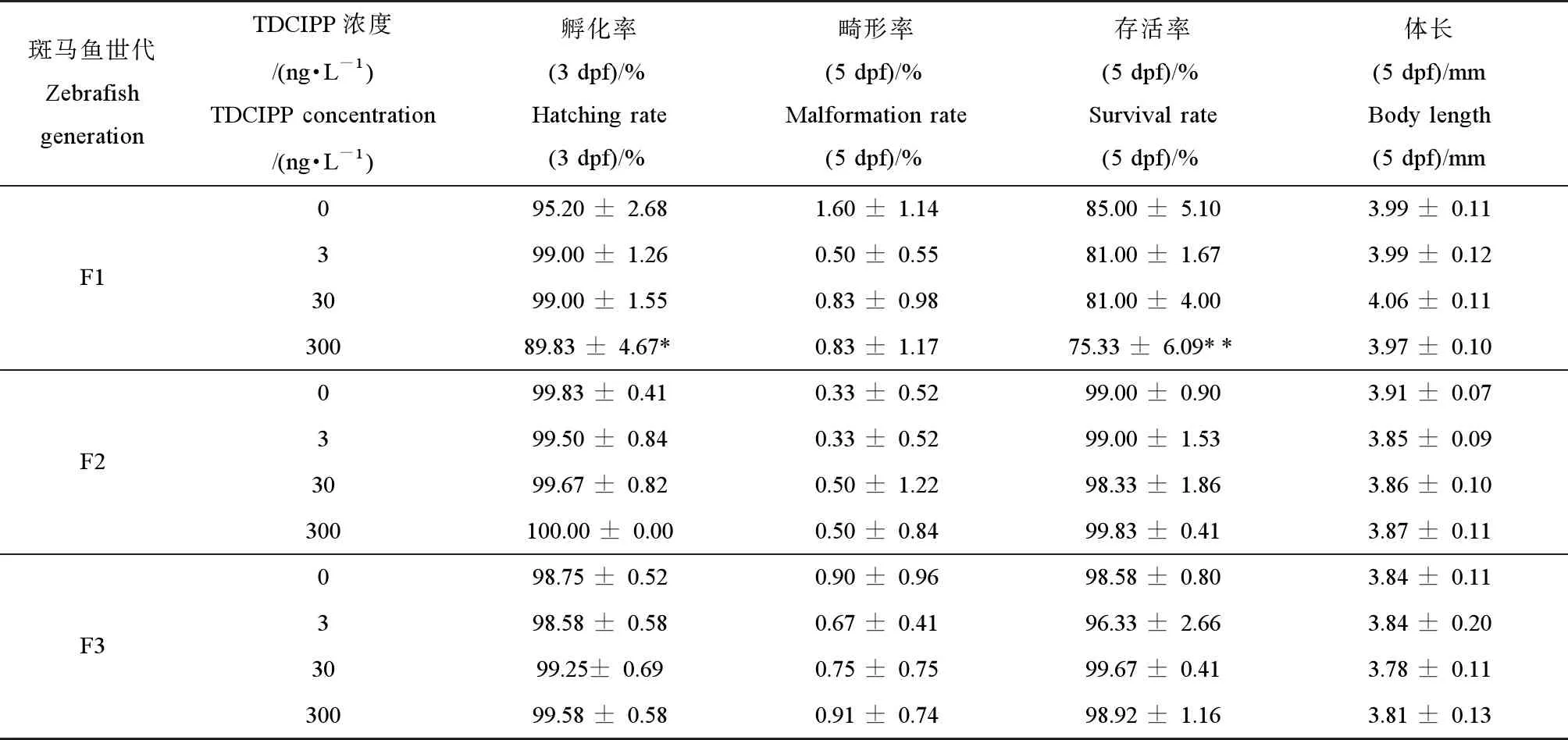
表2 環(huán)境劑量磷酸三(1,3-二氯異丙基)酯(TDCIPP)多代暴露對(duì)F1代、F2代和F3代斑馬魚(yú)胚胎/仔魚(yú)的孵化率、畸形率、成活率和體長(zhǎng)的影響Table 2 Effects of multigenerational exposure to tris (1,3-dichloro-2-propyl) phosphate (TDCIPP) at environmental concertation on the hatching rate, survival rate, malformation rate and body length of F1, F2 or F3 embryos/larvae
注:數(shù)據(jù)表示為平均值±標(biāo)準(zhǔn)差(mean±SD),每個(gè)濃度3個(gè)平行樣,每個(gè)平行樣50條胚胎或仔魚(yú)。*P<0.05、**P<0.01表示暴露組與溶劑對(duì)照組相比具有顯著差異。
Note: The data are expressed as mean ±SD of three replicates (50 embryos/larvae per replicate); *P<0.05, **P<0.01 indicate significant difference detected between solvent control and TDCIPP exposure group.
如圖2所示,與對(duì)照組相比,300 ng·L-1TDCIPP暴露組中F1代120 d后所產(chǎn)子代F2代的游泳速度在黑暗刺激下顯著性降低(P<0.05),在光照周期下無(wú)顯著性差異。
如圖3所示,與對(duì)照組相比,3、30和300 ng·L-1TDCIPP暴露組中F2代120 d后所產(chǎn)子代F3代的游泳速度在黑暗和光照刺激下均無(wú)顯著性變化。
2.3 TDCIPP多代暴露對(duì)各自子代5 dpf仔魚(yú)神經(jīng)發(fā)育相關(guān)基因表達(dá)的影響
檢測(cè)了F1代、F2代和F3代5 dpf仔魚(yú)在TDCIPP暴露下神經(jīng)細(xì)胞生長(zhǎng)發(fā)育及分化相關(guān)的幾種基因的相對(duì)表達(dá)量(圖4)。如圖4(a),在F1代中,與對(duì)照組相比,300 ng·L-1TDCIPP暴露導(dǎo)致神經(jīng)元標(biāo)志基因ngn1表達(dá)量顯著性升高(P<0.01),3和300 ng·L-1TDCIPP暴露導(dǎo)致神經(jīng)元軸突再生和發(fā)育相關(guān)的基因α1-tubulin顯著性上調(diào)(P<0.001、P<0.01),30 ng·L-1TDCIPP暴露導(dǎo)致刺激軸突生長(zhǎng)的netrin1b基因顯著性上調(diào)(P<0.05),3和300 ng·L-1TDCIPP暴露導(dǎo)致運(yùn)動(dòng)神經(jīng)元軸突標(biāo)志基因zn5顯著性上調(diào)(P<0.05,P<0.05),但星形膠質(zhì)細(xì)胞的標(biāo)志基因gfap在300 ng·L-1TDCIPP暴露下顯著性下調(diào)(P<0.01)。
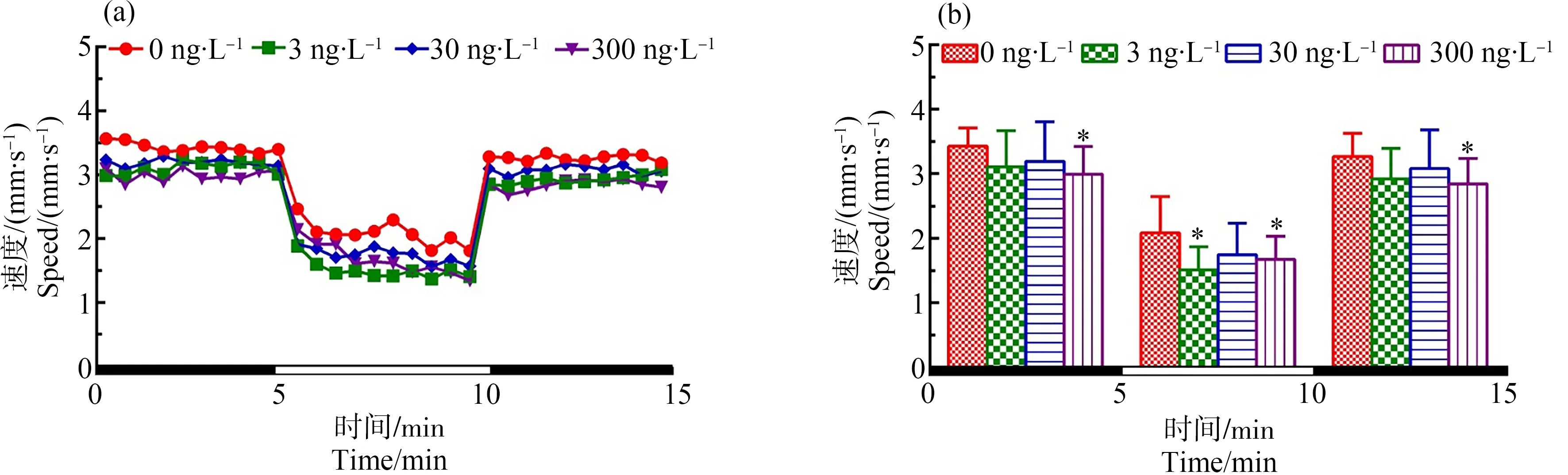
圖1 F0代斑馬魚(yú)暴露于TDCIPP 120 d后對(duì)所產(chǎn)F1代5 dpf仔魚(yú)運(yùn)動(dòng)行為的影響注:(a)光暗周期刺激下斑馬魚(yú)仔魚(yú)的運(yùn)動(dòng)模式,(b)光暗周期刺激下的斑馬魚(yú)仔魚(yú)在各階段的平均游泳速率;數(shù)據(jù)表示為平均值±標(biāo)準(zhǔn)差(mean±SD),每個(gè)濃度3個(gè)平行樣,每個(gè)平行樣8條仔魚(yú);* P<0.05表示暴露組與溶劑對(duì)照組相比具有顯著差異。Fig. 1 Locomotor behavior of the 5 dpf larvae of F1 after F0 parental zebrafish exposed to TDCIPP for 120 dNote: (a) locomotor patterns in response to an alternating light change; (b) the average swimming speed of 5-min intervals for each light state (light or dark); the data are expressed as mean±SD of three replicates (8 larvae per replicate); * P<0.05 indicates significant difference detected between solvent control and TDCIPP exposure group.
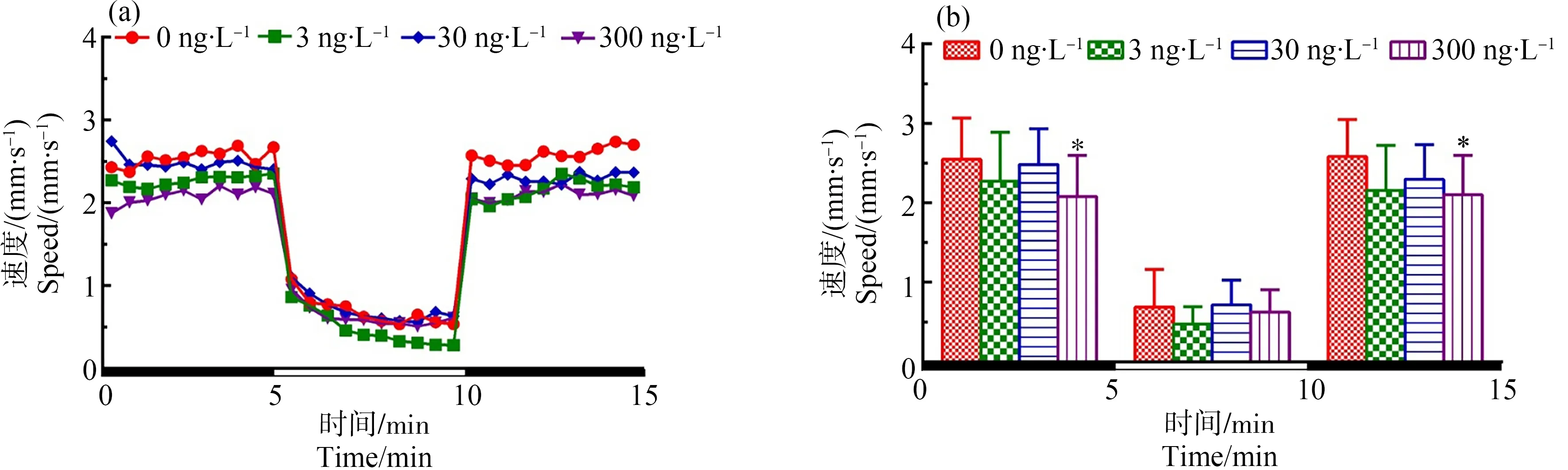
圖2 F1代斑馬魚(yú)暴露于TDCIPP 120 d后對(duì)所產(chǎn)F2代5 dpf仔魚(yú)運(yùn)動(dòng)行為的影響注:(a)光暗周期刺激下斑馬魚(yú)仔魚(yú)的運(yùn)動(dòng)模式,(b)光暗周期刺激下的斑馬魚(yú)仔魚(yú)在各階段的平均游泳速率;數(shù)據(jù)表示為平均值±標(biāo)準(zhǔn)差(mean±SD),每個(gè)濃度3個(gè)平行樣,每個(gè)平行樣8條仔魚(yú);* P<0.05表示暴露組與溶劑對(duì)照組相比具有顯著差異。Fig. 2 Locomotor behavior of the 5 dpf larvae of F2 after F1 parental zebrafish exposed to TDCIPP for 120 dNote: (a) locomotor patterns in response to an alternating light change; (b) the average swimming speed of 5-min intervals for each light state (light or dark); the data are expressed as mean±SD of three replicates (8 larvae per replicate); * P<0.05 indicates significant difference detected between solvent control and TDCIPP exposure group.
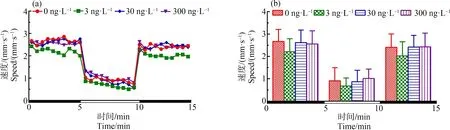
圖3 F2代斑馬魚(yú)暴露于TDCIPP 120 d后對(duì)所產(chǎn)F3代5 dpf仔魚(yú)運(yùn)動(dòng)行為的影響注:(a)光暗周期刺激下斑馬魚(yú)仔魚(yú)的運(yùn)動(dòng)模式,(b)光暗周期刺激下的斑馬魚(yú)仔魚(yú)在各階段的平均游泳速率;數(shù)據(jù)表示為平均值±標(biāo)準(zhǔn)差(mean±SD),每個(gè)濃度3個(gè)平行樣,每個(gè)平行樣8條仔魚(yú)。Fig. 3 Locomotor behavior of the 5 dpf larvae of F3 after F2 parental zebrafish exposed to TDCIPP for 120 dNote: (a) locomotor patterns in response to an alternating light change; (b) the average swimming speed of 5-min intervals for each light state (light or dark); the data are expressed as mean±SD of three replicates (8 larvae per replicate).
如圖4(b)所示,在F2代中,與對(duì)照組相比,最高劑量TDCIPP (300 ng·L-1)暴露導(dǎo)致神經(jīng)元標(biāo)志基因elavl3顯著性下調(diào)(P<0.05),與神經(jīng)元軸突生長(zhǎng)和再生相關(guān)的標(biāo)記基因gap43顯著性下調(diào)(P<0.05),30和300 ng·L-1TDCIPP暴露導(dǎo)致shha基因顯著性下調(diào)(P<0.05、P<0.05),3和300 ng·L-1TDCIPP暴露導(dǎo)致gfap基因表達(dá)量顯著性下調(diào)(P<0.05、P<0.05)。但對(duì)其他基因的表達(dá)均無(wú)顯著性影響。
如圖4(c)所示,在F3代中,與對(duì)照組比較,300 ng·L-1TDCIPP暴露導(dǎo)致gap43表達(dá)量顯著性下降(P<0.05),3和300 ng·L-1TDCIPP暴露導(dǎo)致gfap基因表達(dá)量顯著性下降(P<0.05、P<0.05),但對(duì)其他基因無(wú)顯著性影響。
2.4 相關(guān)分析
F1代、F2代和F3代5 dpf仔魚(yú)神經(jīng)發(fā)育相關(guān)基因表達(dá)的變化與各自游泳速度變化之間的相關(guān)性分析如表3所示。由Spearman相關(guān)性分析可知,F(xiàn)1代5 dpf仔魚(yú)在光暗刺激下的游泳速度與神經(jīng)元標(biāo)記基因ngn1、神經(jīng)元軸突生長(zhǎng)密切相關(guān)的基因α1-tubulin和運(yùn)動(dòng)神經(jīng)元標(biāo)志基因zn5的表達(dá)顯著負(fù)相關(guān)。而F2代和F3代仔魚(yú)在光暗刺激下的游泳速度與神經(jīng)發(fā)育相關(guān)基因的表達(dá)都無(wú)顯著相關(guān)性。

表3 F1代、F2代和F3代5 dpf仔魚(yú)在光暗刺激下的平均游泳速度與神經(jīng)發(fā)育相關(guān)基因表達(dá)的Spearman rank相關(guān)系數(shù)Table 3 Spearman rank correlation coefficients between the average swimming speed of 5-min intervals for each light state (light or dark) and the expressions of genes related to neural development in the 5 dpf zebrafish larvae of F1, F2, F3
注:*0.05水平的顯著相關(guān)(雙尾),** 0.01水平的顯著相關(guān)(雙尾)。
Note: * correlation is significant at the 0.05 level (two-tailed); ** correlation is significant at the 0.01 level (two-tailed).
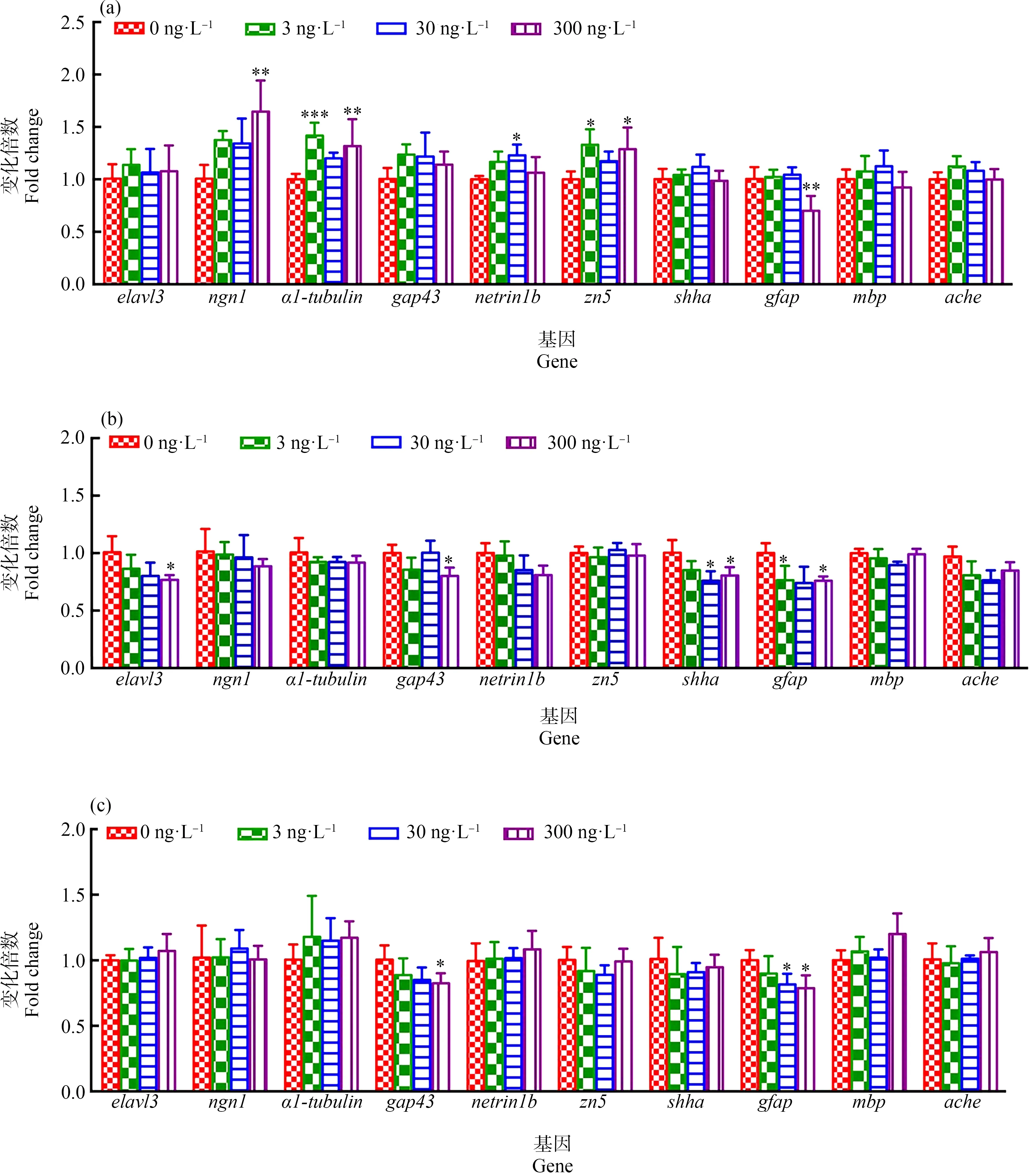
圖4 環(huán)境劑量TDCIPP多代暴露對(duì)斑馬魚(yú)F1代(a)、F2代(b)和F3代(c)的5 dpf仔魚(yú)神經(jīng)發(fā)育相關(guān)基因的影響注:數(shù)據(jù)表示為平均值±標(biāo)準(zhǔn)差(mean±SD),每個(gè)濃度3個(gè)平行樣,每個(gè)平行樣50條仔魚(yú);*P<0.05、** P<0.01和*** P<0.001表示暴露組與溶劑對(duì)照組相比具有顯著差異。Fig. 4 Expressions of genes related to neural development in 5 dpf zebrafish larvae (F1 generation (a), F2 generation (b), F3 generation (c)) under multigenerational exposure to environmental concentration of TDCIPPNote: The data are expressed as mean±SD of three replicates (50 larvae per replicate); * P<0.05, ** P<0.01 and *** P<0.001 indicate significant difference detected between solvent control and TDCIPP exposure group.
3 討論(Discussion)
TDCIPP作為一種新型污染物,經(jīng)常在自然水體中檢測(cè)到(<μg·L-1)。筆者所在團(tuán)隊(duì)前期的研究結(jié)果表明,環(huán)境劑量的TDCIPP暴露,會(huì)導(dǎo)致TDCIPP在斑馬魚(yú)母體內(nèi)蓄積[36-37],并會(huì)傳遞到子代魚(yú)卵中[36]。此外,斑馬魚(yú)持續(xù)暴露于環(huán)境劑量的TDCIPP 2代后,會(huì)在第2代斑馬魚(yú)成魚(yú)體內(nèi)檢測(cè)到其蓄積[38]。目前,這些蓄積的TDCIPP是否會(huì)對(duì)斑馬魚(yú)子代持續(xù)產(chǎn)生不利影響還是未知的。早期發(fā)育階段對(duì)化合物暴露更為敏感,尤其是大腦發(fā)育階段[39],斑馬魚(yú)早期發(fā)育階段神經(jīng)系統(tǒng)中專(zhuān)一表達(dá)的一些基因已證實(shí)可以作為發(fā)育神經(jīng)毒性的生物標(biāo)志[22]。此外,斑馬魚(yú)的游泳行為是在早期發(fā)育后期出現(xiàn)的一個(gè)較復(fù)雜的行為,需要化學(xué)遞質(zhì)傳遞和后腦神經(jīng)系統(tǒng)的輸出[40],仔魚(yú)的運(yùn)動(dòng)行為也可用于指示神經(jīng)毒性效應(yīng)[41]。本實(shí)驗(yàn)采用環(huán)境劑量TDCIPP對(duì)斑馬魚(yú)進(jìn)行3代暴露實(shí)驗(yàn),探究其對(duì)子代斑馬魚(yú)仔魚(yú)生長(zhǎng)發(fā)育和神經(jīng)發(fā)育的影響,以此來(lái)評(píng)估TDCIPP的生態(tài)毒性風(fēng)險(xiǎn)效應(yīng)。
F0代暴露于環(huán)境劑量TDCIPP (300 ng·L-1) 120 d后所產(chǎn)子代F1代仔魚(yú)的孵化率和成活率均顯著性降低。這與筆者所在團(tuán)隊(duì)前期的研究結(jié)果是類(lèi)似的,斑馬魚(yú)暴露于5 000 ng·L-1TDCIPP 240 d導(dǎo)致子代仔魚(yú)的存活率下降、體長(zhǎng)減小和生長(zhǎng)發(fā)育受阻[36]。但在后面2代的持續(xù)暴露中,F(xiàn)2代和F3代仔魚(yú)的孵化率、存活率、畸形率以及體長(zhǎng)均未受到顯著性影響,這些結(jié)果表明,隨著暴露代數(shù)的增加,TDCIPP對(duì)子代斑馬魚(yú)的發(fā)育毒性減弱。
環(huán)境劑量TDCIPP多代暴露對(duì)子代斑馬魚(yú)仔魚(yú)F1代和F2代的游泳行為具有顯著抑制作用,但是對(duì)F3代的游泳行為無(wú)顯著性影響。目前,有關(guān)環(huán)境劑量TDCIPP多代暴露對(duì)生物體影響的研究非常少,而其對(duì)神經(jīng)行為的影響更是鮮有報(bào)道。在筆者的實(shí)驗(yàn)中,300 ng·L-1TDCIPP暴露導(dǎo)致F1代仔魚(yú)在黑暗刺激下的游泳速度均顯著性下降,暴露于3和300 ng·L-1TDCIPP的F1代仔魚(yú)在光照刺激下的行為也顯著性下降。類(lèi)似的結(jié)果在之前的研究中也有報(bào)道,Wang等[16]將斑馬魚(yú)暴露于100 μg·L-1TDCIPP 3個(gè)月后,成魚(yú)的運(yùn)動(dòng)行為未受到顯著性影響,但所產(chǎn)子代仔魚(yú)的平均游泳速率在光照和黑暗下均顯著性下降。此外,筆者所在團(tuán)隊(duì)前期的實(shí)驗(yàn)結(jié)果表明,較高劑量TDCIPP (>300 μg·L-1)短期暴露會(huì)導(dǎo)致斑馬魚(yú)仔魚(yú)的游泳行為在最初黑暗適應(yīng)期的活動(dòng)有所增加,而在光照時(shí)反應(yīng)有所減弱[20],而濃度>100 μg·L-1的TDCIPP暴露不會(huì)影響仔魚(yú)的游泳行為[16,20]。在13 μg·L-1的TDCIPP暴露下,仔魚(yú)在最初黑暗適應(yīng)期的活動(dòng)有所增加,而逃逸反應(yīng)有所減弱[42]。而有研究發(fā)現(xiàn),暴露于較高濃度(1.353和2.413 mg·L-1;3和6 μmol·L-1)TDCIPP下,在光照和黑暗2種條件下斑馬魚(yú)仔魚(yú)的游泳速度均顯著性降低[17,21]。這些研究結(jié)果表明,TDCIPP暴露對(duì)斑馬魚(yú)仔魚(yú)的游泳行為具有顯著的干擾效應(yīng),且母體TDCIPP暴露后的F1代仔魚(yú)比F0代成魚(yú)和仔魚(yú)都要敏感。對(duì)F2代仔魚(yú),300 ng·L-1TDCIPP暴露導(dǎo)致在黑暗下的斑馬魚(yú)仔魚(yú)游泳速率顯著性下降,但在光照刺激下其運(yùn)動(dòng)速度無(wú)顯著性變化。但到F3代仔魚(yú)時(shí),TDCIPP的持續(xù)暴露未引起斑馬魚(yú)的游泳行為的顯著性變化。結(jié)果表明,環(huán)境劑量的TDCIPP暴露會(huì)導(dǎo)致F1代仔魚(yú)的神經(jīng)發(fā)育毒性,但是TDCIPP持續(xù)多代暴露并未導(dǎo)致毒性的增強(qiáng),反而隨著暴露代數(shù)的增加,斑馬魚(yú)仔魚(yú)對(duì)TDCIPP的敏感性呈現(xiàn)降低的趨勢(shì)。
有研究表明,斑馬魚(yú)運(yùn)動(dòng)行為的改變與運(yùn)動(dòng)神經(jīng)元的發(fā)育受影響相關(guān)[43-44]。為探究TDCIPP可能的神經(jīng)發(fā)育毒性機(jī)制,分析了仔魚(yú)神經(jīng)相關(guān)基因的轉(zhuǎn)錄水平,并將其與運(yùn)動(dòng)行為進(jìn)行了相關(guān)性分析。在F1代仔魚(yú)中,神經(jīng)元標(biāo)記基因ngn1表達(dá)量顯著性升高,與神經(jīng)元軸突生長(zhǎng)密切相關(guān)的基因α1-tubulin、netrin1b和zn5顯著性上調(diào),這些基因表達(dá)的上調(diào)可能是對(duì)神經(jīng)元軸突損傷的一個(gè)補(bǔ)償性反饋。在之前的研究中,α1-tubulin表達(dá)量的上調(diào)被認(rèn)為是對(duì)化學(xué)品暴露后斑馬魚(yú)仔魚(yú)軸突生長(zhǎng)受到的抑制作用的補(bǔ)償性反饋(如得克隆(DP)和2,2’,4,4’-四溴聯(lián)苯醚(BDE-47)暴露實(shí)驗(yàn))[43-44]。實(shí)際上,在筆者所在團(tuán)隊(duì)的前期實(shí)驗(yàn)中也發(fā)現(xiàn),TDCIPP急性暴露會(huì)導(dǎo)致運(yùn)動(dòng)神經(jīng)元軸突的損傷,并伴隨著軸突生長(zhǎng)相關(guān)基因α1-tubulin、netrin表達(dá)量的上調(diào)[20]。環(huán)境劑量TDCIPP暴露主要影響了F1代神經(jīng)元和軸突生長(zhǎng)相關(guān)基因的表達(dá),而對(duì)神經(jīng)遞質(zhì)乙酰膽堿酶活基因(acetylcholinesterase,ache)、髓磷脂堿性蛋白基因(myelin basic protein,mbp)等均無(wú)顯著性影響,相關(guān)性分析顯示,游泳速度的抑制與ngn1、α1-tubulin、gap43和zn5這4個(gè)基因的表達(dá)顯著負(fù)相關(guān),這些結(jié)果表明,TDCIPP可能主要通過(guò)影響神經(jīng)元軸突的生長(zhǎng)來(lái)導(dǎo)致游泳行為受阻,這與前期的實(shí)驗(yàn)結(jié)果類(lèi)似[20]。在F2代斑馬魚(yú)仔魚(yú)中,TDCIPP暴露導(dǎo)致神經(jīng)元標(biāo)志基因elavl3(編碼HuC)表達(dá)量顯著性下調(diào),表明神經(jīng)發(fā)育受到了損傷。生長(zhǎng)相關(guān)蛋白gap43基因的表達(dá)量也顯著性降低,gap43常常用作神經(jīng)損傷后再生時(shí)重新誘導(dǎo)軸突生長(zhǎng)的標(biāo)志[20],該基因的下調(diào)表明受損神經(jīng)的再生受到了影響。此外,星形膠質(zhì)細(xì)胞標(biāo)記基因gfap以及連接視網(wǎng)膜和脊髓間的軸突索基因shha均顯著性下降。這些基因表達(dá)的下調(diào)表明TDCIPP暴露可能對(duì)神經(jīng)元、神經(jīng)膠質(zhì)細(xì)胞的數(shù)量和再生均產(chǎn)生了影響,表現(xiàn)出明顯的神經(jīng)發(fā)育毒性。然而,相關(guān)性分析表明,F(xiàn)2代仔魚(yú)游泳速度的降低與這些基因表達(dá)的下降無(wú)顯著相關(guān)性,提示TDCIPP可能不是通過(guò)影響神經(jīng)細(xì)胞的數(shù)量而影響?hù)~(yú)的游泳行為。在F3代中,僅gap43和gfap這2個(gè)基因的表達(dá)量受到TDCIPP暴露的抑制,表明隨著暴露代數(shù)的增加,TDCIPP的神經(jīng)發(fā)育毒性在減弱,這與游泳行為以及生長(zhǎng)發(fā)育的結(jié)果是一致的。
綜上所述,環(huán)境劑量TDCIPP暴露對(duì)斑馬魚(yú)子代仔魚(yú)具有神經(jīng)發(fā)育毒性,但是隨著暴露代數(shù)的增加TDCIPP的毒性效應(yīng)呈現(xiàn)減弱的趨勢(shì)。TDCIPP暴露導(dǎo)致F0代所產(chǎn)F1代仔魚(yú)的生長(zhǎng)發(fā)育以及運(yùn)動(dòng)行為受到顯著的抑制,其對(duì)運(yùn)動(dòng)速度的影響可能主要源于神經(jīng)元軸突的生長(zhǎng)受到了影響,表現(xiàn)為運(yùn)動(dòng)行為抑制與神經(jīng)軸突生長(zhǎng)相關(guān)基因(ngn1、α1-tubulin和zn5)的表達(dá)量顯著負(fù)相關(guān)。繼續(xù)暴露后,F(xiàn)2代仔魚(yú)的生長(zhǎng)發(fā)育不再受到顯著性影響,運(yùn)動(dòng)行為僅在最高劑量組黑暗處理下受到抑制,神經(jīng)發(fā)育相關(guān)基因(elavl3、gap43、gfap和shha)表達(dá)量顯著下降,但運(yùn)動(dòng)的抑制與基因表達(dá)無(wú)顯著相關(guān)性。暴露至F3代仔魚(yú),其生長(zhǎng)發(fā)育和神經(jīng)行為均不再受影響,僅gap43和gfap這2個(gè)基因表達(dá)量下調(diào)。表明隨著暴露代數(shù)的增加,TDCIPP的神經(jīng)發(fā)育毒性效應(yīng)降低。筆者首次研究了生物體多代處于環(huán)境劑量TDCIPP暴露后的毒性效應(yīng),為T(mén)DCIPP的生態(tài)風(fēng)險(xiǎn)評(píng)價(jià)提供了理論依據(jù)。在進(jìn)一步的研究中,將結(jié)合化學(xué)分析和分子生物學(xué)手段,開(kāi)展深層次的研究,力圖從化合物在生物體內(nèi)的蓄積、代謝以及生物體的解毒代謝等多方面來(lái)揭示TDCIPP多代持續(xù)暴露后毒性減弱的機(jī)制。
猜你喜歡
全科護(hù)理(2022年10期)2022-12-26 21:19:15
中國(guó)合理用藥探索(2022年1期)2022-11-26 00:22:32
今日農(nóng)業(yè)(2022年4期)2022-11-16 19:42:02
鄉(xiāng)村科技(2021年33期)2021-03-16 02:26:54
國(guó)際放射醫(yī)學(xué)核醫(yī)學(xué)雜志(2021年10期)2021-02-28 08:41:58
藥學(xué)與臨床研究(2015年4期)2015-06-05 11:35:54
衛(wèi)生職業(yè)教育(2014年24期)2014-05-20 09:05:38
同位素(2014年2期)2014-04-16 04:57:20
中國(guó)合理用藥探索(2014年11期)2014-03-11 20:30:20
