溶液體系中迷迭香酸與肌球蛋白的相互作用及其對(duì)蛋白理化特性的影響
2020-07-04 10:58:02陳雪珂戴宏杰朱瀚昆王洪霞張宇昊
食品科學(xué) 2020年12期
周 揚(yáng),陳雪珂,戴宏杰,余 永,朱瀚昆,王洪霞,張宇昊*
(西南大學(xué)食品科學(xué)學(xué)院,重慶 400715)
肌球蛋白是肉及肉制品中最主要的蛋白質(zhì),約占肌肉總蛋白的1/3,是蛋白功能特性的主要承擔(dān)者,對(duì)肉制品品質(zhì)起關(guān)鍵性作用[1-2]。迷迭香是肉制品加工、貯藏等過(guò)程中常見(jiàn)的調(diào)料,賦予了產(chǎn)品特殊風(fēng)味并起到抗氧化等作用,其中起主要作用的成分為多酚類化合物。迷迭香酸(rosmarinic acid,RA)是一種水溶性多酚,被鑒定為迷迭香提取物的主要成分之一,具有抗氧化、抗炎、抗血栓生成、抗菌等生物活性,廣泛應(yīng)用于食品、醫(yī)藥、化妝品等領(lǐng)域[3]。
目前,將RA或迷迭香提取物用于延緩肉類制品的脂肪氧化和蛋白質(zhì)氧化已有報(bào)道。高輝等[4]在火腿腸中添加了適量RA,有效延緩了火腿腸的脂肪氧化。張曉潔[5]將RA添加到明膠膜中用于豬肉的保鮮研究,結(jié)果表明RA可以提高明膠膜的抗氧化性,延緩臘肉的脂質(zhì)氧化。此外,也有將迷迭香提取物添加到肉類產(chǎn)品中,賈娜等[6]將迷迭香提取物添加到雞肉糜中,具有抑制雞肉糜在冷藏過(guò)程中的脂肪氧化作用,并改善了肉制品的品質(zhì)。其中,RA等多酚類物質(zhì)會(huì)與肉制品中的蛋白質(zhì)發(fā)生相互作用,進(jìn)而影響蛋白質(zhì)的結(jié)構(gòu)特征和理化特性,導(dǎo)致其功能特性發(fā)生改變。諸多研究表明,蛋白質(zhì)與多酚類化合物之間的相互作用分為可逆的非共價(jià)相互作用和不可逆的共價(jià)相互作用,但因?yàn)榈鞍踪|(zhì)的高度復(fù)雜性、多酚的多反應(yīng)活性、蛋白與多酚種類及反應(yīng)條件的不同,其相互作用呈現(xiàn)出復(fù)雜性。目前,關(guān)于肉類蛋白與RA的互作研究大部分都基于氧化條件下對(duì)于蛋白凝膠特性的影響。而在非氧化條件下,肌球蛋白與RA在溶液體系中的相互作用及其與蛋白理化特性的內(nèi)在聯(lián)系的研究較少。
本研究以肌球蛋白和RA為原料制備混合溶液,運(yùn)用熒光光譜、圓二色譜、電泳等方法表征RA與肌球蛋白的相互作用,并測(cè)定不同鹽濃度條件下肌球蛋白的構(gòu)象變化和功能特性等指標(biāo),旨在探究溶液狀態(tài)下RA與肌球蛋白的相互作用及其對(duì)肌球蛋白理化特性的影響,并闡明相關(guān)機(jī)理。
1 材料與方法
1.1 材料與試劑
新鮮豬里脊肉、大豆油 重慶市北碚區(qū)永輝超市;RA(純度97%) 上海阿拉丁試劑公司;甘氨酸生工生物工程上海有限公司;十二烷基硫酸鈉(sodium dodecyl sulfate,SDS)、過(guò)硫酸銨、β-巰基乙醇(β-mercaptoethanol,β-ME)、三羥甲基氨基甲烷(Tris,分析純)、四甲基乙二胺、考馬斯亮藍(lán)R-250(分析純) 上海佰曄生物科技中心;標(biāo)準(zhǔn)蛋白(分子質(zhì)量10~200 kDa) 加拿大Fermentas公司;三磷酸腺苷二鈉鹽、乙二醇-雙-(2-氨基乙醚)四乙酸(ethylenebis(oxyethylenenitrilo)tetraacetic acid,EGTA)、5,5-二硫代雙-(2-硝基苯甲酸)(5,5’-dithio bis-(2-nitrobenzoic acid),DTNB)、溴酚藍(lán)(bromophenol blue,BPB)、丙烯酰胺(分析純,質(zhì)量分?jǐn)?shù)30%) 北京索萊寶科技有限公司;氯化鈉成都市科龍化工試劑廠;其他試劑均為分析純。
1.2 儀器與設(shè)備
JA3003B型電子天平 上海精天電子儀器有限公司;PHS-25型數(shù)顯酸度計(jì) 杭州雷磁分析儀器廠;QL 901 Vortex型渦旋混合器 海門市其林貝爾儀器制造有限公司;XHF-D型高速分散器 寧波新芝有限公司;Heraeus Multifuge X3R型高速冷凍離心機(jī) 美國(guó)賽默飛世爾科技公司;722-P型紫外-可見(jiàn)光分光光度計(jì) 上海菁華科技儀器有限公司;F-4500型熒光分光光度計(jì) 日本日立公司;Power PacTM型基礎(chǔ)電泳儀 美國(guó)Bio-Rad公司;G:BOX EF型凝膠成像系統(tǒng) 英國(guó)Syngene公司;MOS-500型圓二色譜儀 法國(guó)Bio-Logic公司;ZEN3690型粒度儀 英國(guó)馬爾文儀器公司。
1.3 方法
1.3.1 肌球蛋白的提取
參照Liu Ru等[7]的方法略作修改。為了防止蛋白質(zhì)變性和蛋白質(zhì)水解,用于制備肌球蛋白的溶液要在4 ℃進(jìn)行保存。取豬里脊肉,切成肉糜狀,加入10 倍體積的試劑A(0.1 mol/L KCl,20 mmol/L Tris-HCl緩沖液,用鹽酸調(diào)節(jié)pH值至7.5)。用均質(zhì)機(jī)在10 000 r/min條件下將其均質(zhì)1 min。混合物在4 ℃放置15 min,5 000 r/min、4 ℃離心5 min。取沉淀,加入5 倍體積的試劑B(0.45 mol/L KCl,5 mmol/L β-ME,0.2 mol/L乙酸鎂,1 mmol/L EGTA,20 mmol/L Tris-HCl緩沖液,用馬來(lái)酸調(diào)節(jié)pH值至6.8),再加ATP,使其最終濃度為10 mmol/L,混合物在4 ℃放置90 min,10 000 r/min、4 ℃離心10 min。取上清液,加入5 倍體積的1 mmol/L KHCO3進(jìn)行緩慢稀釋,4 ℃放置20 min,10 000 r/min離心10 min(4 ℃)。取沉淀,加入2.5 倍體積的試劑C(0.5 mol/L KCl,5 mmol/L β-ME,20 mmol/L Tris-HCl緩沖液,用鹽酸調(diào)節(jié)pH值至7.5),4 ℃保存10 min,然后緩慢加入7 倍體積的1 mmol/L KHCO3,同時(shí),加入MgCl2使其最終濃度為10 mmol/L。將形成的混合物在4 ℃保存12 h,10 000 r/min、4 ℃離心10 min。取沉淀物,即為肌球蛋白,4 ℃保存,3 d內(nèi)使用,肌球蛋白含量的測(cè)定采用雙縮脲法。
1.3.2 肌球蛋白-RA混合液的制備
將提取的肌球蛋白分散于20 mmol/L Tris-HCl(pH 7.0)緩沖液中,并與RA溶液(溶于相同緩沖液)混合均勻,RA添加質(zhì)量分?jǐn)?shù)2%。向混合液中加入固體NaCl,使NaCl的最終濃度分別為0.2、0.4、0.6、0.8、1.0 mol/L,攪拌均勻,得到一系列不同鹽濃度條件下的肌球蛋白-RA混合液(myosin-rosmarinic acid,M-RA)。對(duì)照組(M)不含RA。
1.3.3 色氨酸熒光測(cè)定
采用熒光分光光度儀檢測(cè)肌球蛋白的內(nèi)源色氨酸熒光[8]。室溫下,將肌球蛋白分散于20 mmol/L Tris-HCl緩沖液(含0.6 mol/L NaCl,pH 7.0)中,將RA溶液(溶于相同緩沖液)按照不同比例混合,得到一系列混合溶液,其最終肌球蛋白質(zhì)量濃度為0.2 mg/mL,RA質(zhì)量濃度為0~0.014 mg/mL。混合均勻后測(cè)量在3 個(gè)不同溫度(298、308、318 K)條件下的肌球蛋白熒光光譜。激發(fā)波長(zhǎng)為295 nm,激發(fā)和發(fā)射狹縫寬度均設(shè)置為5 nm,掃描范圍為300~500 nm。
Stern-Volmer方程:
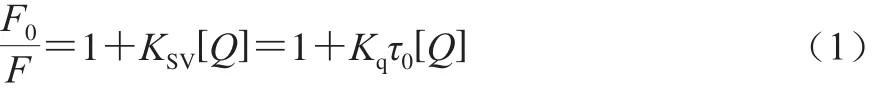
式中:F0和F分別為淬滅劑不存在和存在的熒光強(qiáng)度;KSV為速率常數(shù)/(L/mol);Kq為猝滅常數(shù)/(L/(mol·s));[Q]為淬滅劑的濃度/(mol/L);生物大分子τ0=10-8s-1。
Lineweaver-Burk靜態(tài)猝滅公式:
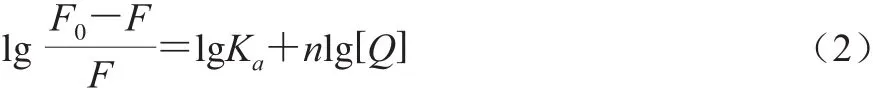
式中:Ka為表觀結(jié)合常數(shù)/(L/mol);n為結(jié)合位點(diǎn);[Q]為熒光猝滅劑的濃度/(mol/L)。
Van’t Hoff等式:

式中:K為溫度T時(shí)的結(jié)合常數(shù);R為氣體常數(shù);ΔH為焓變;ΔS為熵變;ΔG為吉布斯自由能。
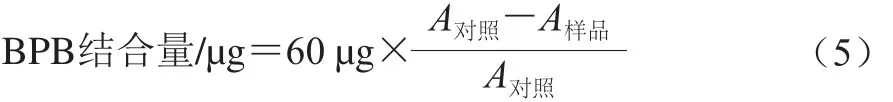
1.3.4 表面疏水性
參照Chelh等[9]的方法略作修改。將樣品調(diào)至蛋白質(zhì)量濃度為2 mg/mL,取2 mL樣液于離心管中,分別加入60 μL的1 mg/mL溴酚藍(lán)溶液,振蕩均勻,10 000 r/min離心10 min。取上清液再次離心后于595 nm波長(zhǎng)處測(cè)定吸光度。以緩沖溶液為空白對(duì)照,蛋白結(jié)合的BPB含量作為疏水性指標(biāo),計(jì)算如式(5)所示:
1.3.5 總巰基測(cè)定
參照Liu Jianhua等[10]的方法,并略作修改。將樣品調(diào)至蛋白質(zhì)量濃度為2 mg/mL,取1 mL樣液懸浮于2 mL的Tris-甘氨酸緩沖液(0.086 mol/L Tris,0.09 mol/L甘氨酸,4 mmol/L EDTA,8 mol/L尿素,pH 8.0)中,加入0.5 mL 10 mmol/L Ellman試劑(0.2 g DTNB溶解于50 mL的Tris-甘氨酸緩沖液中),混合均勻后靜置30 min,于412 nm波長(zhǎng)處測(cè)吸光度。以緩沖溶液為空白對(duì)照,計(jì)算如式(6)所示:
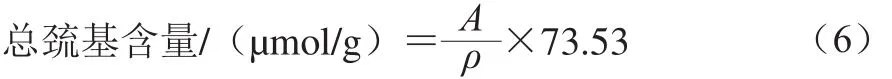
式中:A為412 nm波長(zhǎng)處的吸光度,ρ為樣品的蛋白質(zhì)量濃度/(mg/mL)。
1.3.6 電泳分析
制備12%分離膠、4%濃縮膠。將樣品調(diào)至蛋白質(zhì)量濃度為2 mg/mL,取20 μL樣品并添加5 μL樣品緩沖液(250 mmol/L Tris-HCl,pH 6.8)、0.1 g/mL十二烷基硫酸鈉、5 mg/mL溴酚藍(lán)、體積分?jǐn)?shù)50%甘油、體積分?jǐn)?shù)5% β-ME,沸水浴5 min后立即于冰水浴中冷卻,上樣量為10 μL。電流為15 mA,待溴酚藍(lán)從濃縮膠跑到分離膠中部后,電流調(diào)至25 mA,電泳結(jié)束后用考馬斯亮藍(lán)R-250染色液進(jìn)行染色,2 h后用脫色液振蕩脫色,待條帶清晰后拍攝電泳圖譜。
1.3.7 圓二色譜掃描
采用圓二色譜儀測(cè)定肌球蛋白的二級(jí)結(jié)構(gòu)。將樣品調(diào)至蛋白質(zhì)量濃度為0.1 mg/mL,圓二色譜掃描波長(zhǎng)范圍為190~250 nm,掃描速率為100 nm/min,樣品池光徑為1 mm。以緩沖液為空白對(duì)照,計(jì)算如式(7)所示:
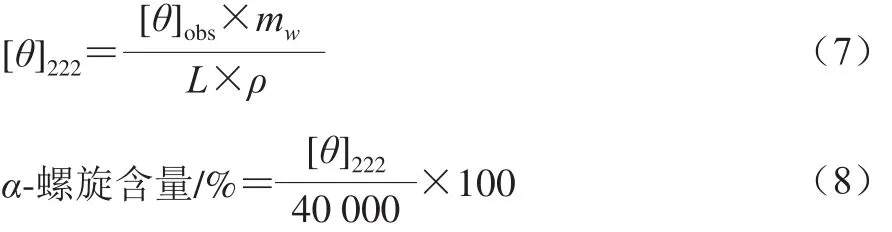
式中:[θ]222為2 2 2 n m波長(zhǎng)處的摩爾橢圓率/(deg·cm2/dmol);[θ]obs為222 nm波長(zhǎng)處的橢圓率/mdeg;mw為肌球蛋白的平均殘基分子質(zhì)量,取115 g/mol;ρ為肌球蛋白質(zhì)量濃度/(mg/mL);L為比色皿光程長(zhǎng)度/mm。
1.3.8 溶解度和濁度測(cè)定
溶解度:將樣品調(diào)至蛋白質(zhì)量濃度為2 mg/mL,4 ℃、10 000 r/min離心15 min,測(cè)定上清液中的蛋白濃度,蛋白溶解度計(jì)算如式(9)所示:

濁度:將樣品調(diào)至蛋白質(zhì)量濃度為2 mg/mL,于370 nm波長(zhǎng)處測(cè)定吸光度。
1.3.9 Zeta電位和粒徑測(cè)定
采用馬爾文粒徑儀測(cè)定肌球蛋白樣品的電位和粒徑分布。將樣品調(diào)至蛋白質(zhì)量濃度0.5 mg/mL,粒度分析儀為He-Ne光源,功率5 mW,散射角90°,平衡時(shí)間60 s。
1.3.10 乳化特性測(cè)定
采用濁度法測(cè)定肌球蛋白的乳化性質(zhì)[11]。將樣品調(diào)至蛋白質(zhì)量濃度為0.2 mg/mL,取8.0 mL置于離心管中,并加入2.0 mL大豆油,10 000 r/min均質(zhì)1 min,立即從距離心管底0.5 cm的地方取勻漿液50 μL,加入到5 mL質(zhì)量分?jǐn)?shù)為0.1%的SDS溶液中,振蕩混勻后于500 nm波長(zhǎng)處測(cè)定吸光度記為A0;靜置10 min再次在相同的位置取勻漿液50 μL,加入到5 mL相同SDS溶液中,振蕩混勻后測(cè)定吸光度記為A10。以0.1% SDS溶液為空白對(duì)照,乳化活力和乳化穩(wěn)定性的計(jì)算公式如下:
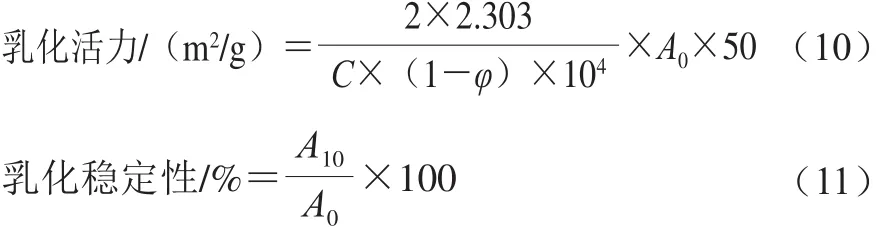
式中:A0、A10為乳狀液在第0、10分鐘的吸光度;φ為油相體積分?jǐn)?shù)(φ=0.2);C為蛋白質(zhì)量濃度/(mg/mL)。
1.4 數(shù)據(jù)分析
數(shù)據(jù)處理采用SPSS 17.0和Origin 8.0軟件,每次實(shí)驗(yàn)重復(fù)3 次。在分析過(guò)程中對(duì)數(shù)據(jù)進(jìn)行方差分析,利用Duncan多重比較進(jìn)行顯著性分析,P<0.05,差異顯著。
2 結(jié)果與分析
2.1 熒光光譜分析
2.1.1 色氨酸熒光結(jié)果
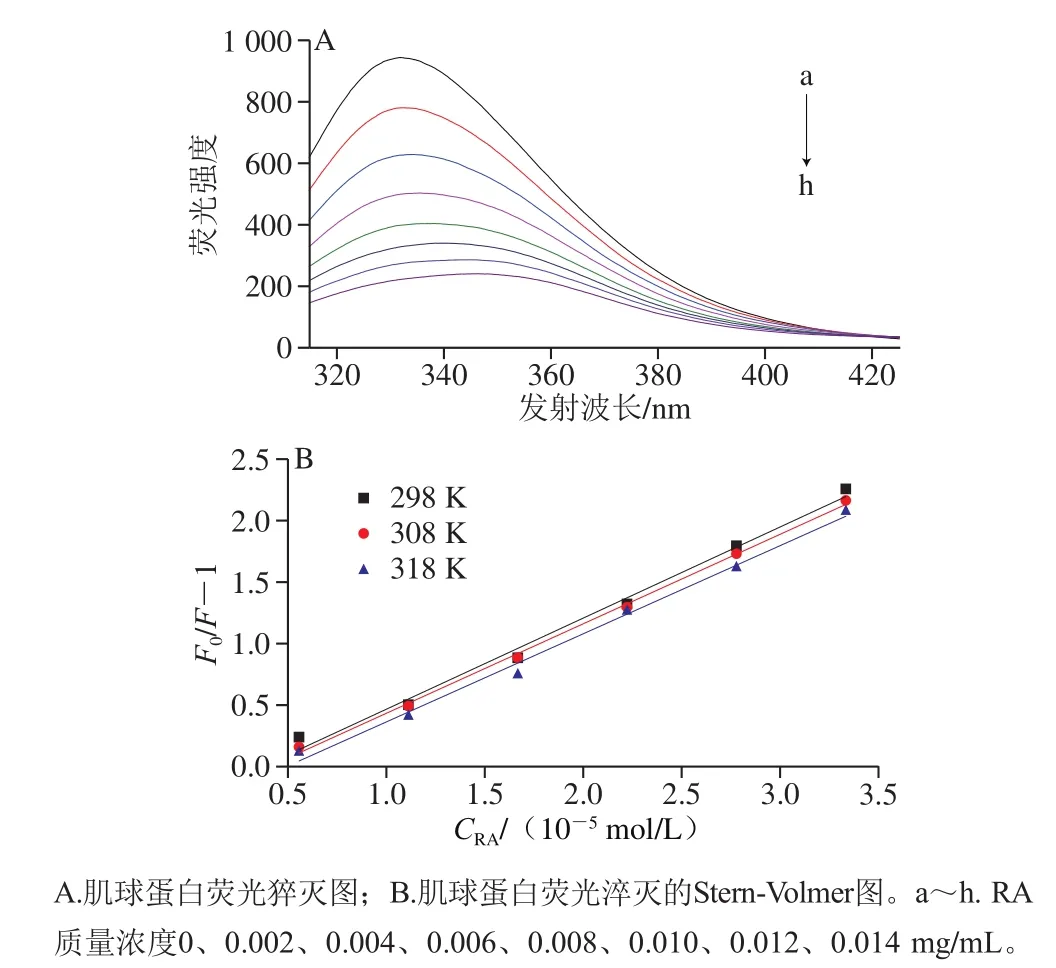
圖1 RA對(duì)肌球蛋白內(nèi)源熒光的淬滅作用Fig. 1 Effect of RA concentration on tryptophan fluorescence of myosin

表1 不同溫度下RA與肌球蛋白相互作用的猝滅常數(shù)(Kq)和速率常數(shù)(Ksv)Table 1Stern-Volmer quenching constants for M-RA system at different temperatures
在一定的激發(fā)波長(zhǎng)下,蛋白質(zhì)分子中的芳香族氨基酸(色氨酸、酪氨酸和苯丙氨酸)可以產(chǎn)生熒光,其中,色氨酸的熒光最強(qiáng)[12]。蛋白溶液中添加多酚后,可能會(huì)導(dǎo)致熒光強(qiáng)度降低,即熒光猝滅,故可以通過(guò)研究蛋白的色氨酸熒光了解蛋白與多酚之間的相互作用。如圖1A所示,當(dāng)激發(fā)波長(zhǎng)為295 nm時(shí),肌球蛋白被激發(fā)后展現(xiàn)出很強(qiáng)的熒光強(qiáng)度。隨RA添加量的增加,肌球蛋白的熒光強(qiáng)度逐漸降低,且伴隨著最大發(fā)射峰出現(xiàn)明顯的紅移現(xiàn)象(從332 nm移至344 nm),說(shuō)明RA對(duì)于肌球蛋白內(nèi)源熒光具有明顯的猝滅作用,二者可能通過(guò)相互作用促使蛋白結(jié)構(gòu)展開(kāi),進(jìn)而使色氨酸從蛋白內(nèi)部疏水性位置暴露到極性環(huán)境中[13]。
一般而言,導(dǎo)致蛋白質(zhì)熒光猝滅的方式通常可分為動(dòng)態(tài)猝滅和靜態(tài)猝滅。為進(jìn)一步闡明RA對(duì)肌球蛋白的猝滅機(jī)理,采用經(jīng)典Stern-Volmer方程對(duì)熒光數(shù)據(jù)進(jìn)行分析[14]。如圖1B、表1所示,隨溫度升高,猝滅曲線的斜率(Ksv)逐漸降低,且猝滅常數(shù)Kq遠(yuǎn)大于最大動(dòng)態(tài)猝滅常數(shù)(2.0×1010L/(mol·s))[15],說(shuō)明RA引起的肌球蛋白熒光猝滅是分子之間結(jié)合形成復(fù)合物所引起的靜態(tài)熒光猝滅,而不是由分子擴(kuò)散和碰撞所引起的動(dòng)態(tài)熒光猝滅。
2.1.2 結(jié)合情況和作用力類型判斷

表2 不同溫度下RA與肌球蛋白結(jié)合的相關(guān)常數(shù)和方程Table 2Regression equations and correlation coef ficients for RA-M system at different temperatures

表3 RA和肌球蛋白相互作用的熱力學(xué)參數(shù)Table 3 Thermodynamic parameters of interaction between RA and myosin
如表2所示,溫度對(duì)肌球蛋白與RA的結(jié)合常數(shù)影響不大,Ka值均大于104L/mol,且可形成一個(gè)結(jié)合位點(diǎn),表明RA與肌球蛋白之間結(jié)合力很好。
如表3所示,通過(guò)反應(yīng)的熱力學(xué)參數(shù)的計(jì)算可大致確定作用力的類型。肌球蛋白與RA的作用過(guò)程是一個(gè)放熱過(guò)程(ΔH<0),熵增過(guò)程(ΔS>0)及自發(fā)過(guò)程(ΔG<0),且熵增加是肌球蛋白與RA相互作用過(guò)程的主要驅(qū)動(dòng)力。在水溶液中,ΔS>0,可認(rèn)為二者之間主要的作用力為疏水作用,這是因?yàn)樗芤褐杏须x子間的靜電作用使得ΔH<0[16]。因此,肌球蛋白與RA之間以疏水相互作用為主,還包括氫鍵和范德華力等,如圖2所示。He Wenjia等[17]分析熱力學(xué)結(jié)果得到,矢車菊素-3-O-葡萄糖苷(cyanidin-3-O-glucoside,C3G)與β-乳球蛋白主要通過(guò)氫鍵和疏水相互作用結(jié)合,C3G與β-酪蛋白則主要通過(guò)氫鍵和范德華力進(jìn)行相互作用。這些不同的結(jié)果可能是由于蛋白和多酚種類及狀態(tài)不同所致。
2.2 表面疏水性分析
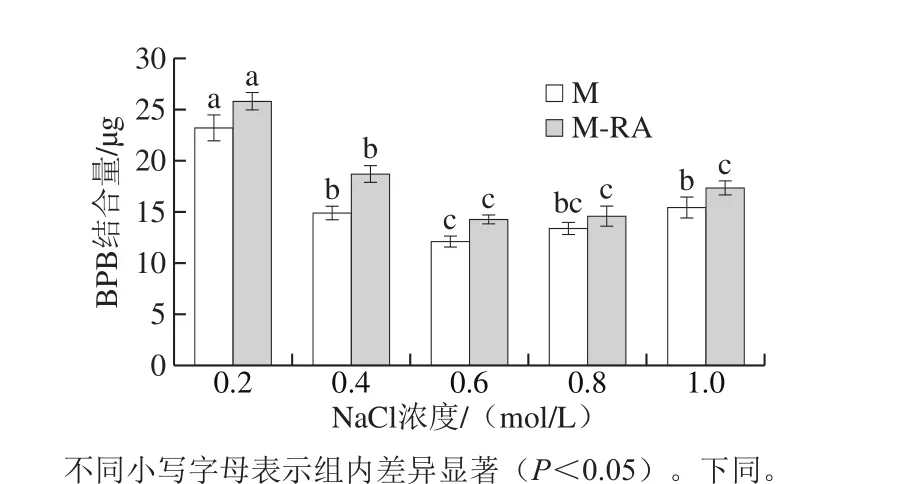
圖2 不同NaCl濃度下RA對(duì)肌球蛋白表面疏水性的影響Fig. 2 Effect of RA on surface hydrophobicity of myosin at different NaCl concentrations
如圖2所示,當(dāng)NaCl濃度從0.2 mol/L增加至0.6 mol/L時(shí),兩組疏水性均呈顯著減少的趨勢(shì)(P<0.05),這可能是因?yàn)殡SNaCl濃度升高,蛋白質(zhì)周圍的親水基團(tuán)逐漸結(jié)合水分子,允許部分疏水性氨基酸殘基包埋在蛋白質(zhì)分子內(nèi)部,引起表面疏水性下降。當(dāng)NaCl濃度從0.6 mol/L增加至1.0 mol/L時(shí),M組疏水性顯著增加(P<0.05),而M-RA組無(wú)顯著變化(P>0.05),這可能是因?yàn)榧∏虻鞍自诟啕}濃度下出現(xiàn)鹽析作用導(dǎo)致疏水作用增加,而添加RA可以抑制這種作用。總體而言,添加RA導(dǎo)致肌球蛋白疏水性升高,說(shuō)明RA可以促進(jìn)肌球蛋白結(jié)構(gòu)的展開(kāi),暴露埋藏在蛋白分子內(nèi)部的疏水性基團(tuán)。結(jié)合色氨酸熒光結(jié)果,進(jìn)一步表明添加RA可以促使蛋白結(jié)構(gòu)展開(kāi)并暴露疏水性基團(tuán),從而利于肌球蛋白與RA發(fā)生疏水相互作用。
2.3 總巰基分析
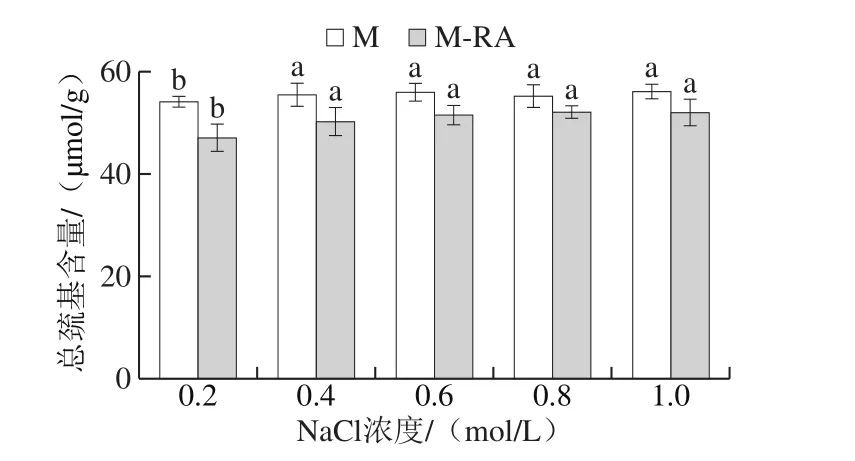
圖3 不同NaCl濃度下RA對(duì)肌球蛋白總巰基的影響Fig. 3 Effect of RA on total sulfhydryl content of myosin at different NaCl concentrations
總巰基包括埋藏在蛋白分子內(nèi)部和暴露在蛋白分子表面的巰基,暴露于分子表面的巰基極易被氧化成二硫鍵,引起蛋白質(zhì)分子間交聯(lián),進(jìn)而影響蛋白質(zhì)的功能特性[18]。如圖3所示,當(dāng)NaCl濃度從0.2 mol/L增加至0.4 mol/L時(shí),兩組總巰基含量均顯著增加(P<0.05);NaCl濃度在0.4~1.0 mol/L范圍內(nèi),兩組總巰基含量無(wú)顯著變化(P>0.05),說(shuō)明此時(shí)NaCl濃度對(duì)蛋白巰基含量無(wú)顯著性影響。總體而言,添加RA導(dǎo)致肌球蛋白總巰基含量下降,這可能是因?yàn)镽A促進(jìn)蛋白結(jié)構(gòu)展開(kāi),埋藏在蛋白分子內(nèi)部的部分巰基暴露出來(lái),進(jìn)而被氧化為二硫鍵等導(dǎo)致巰基含量降低[19]。此外,巰基的減少還可能是因?yàn)榉宇惢衔锱c蛋白巰基發(fā)生相互作用。賈娜等[20]將迷迭香提取物添加到肌原纖維蛋白中,迷迭香提取物中的多酚類化合物均含有巰基結(jié)合位點(diǎn),因而使得巰基含量下降。Tang Changbo等[21]研究發(fā)現(xiàn)高濃度的多酚能與蛋白中的巰基發(fā)生共價(jià)結(jié)合,生成巰基-醌加成物。

圖4 不同NaCl濃度下RA對(duì)肌球蛋白SDS-PAGE的影響Fig. 4 Effect of RA on SDS-PAGE pattern of myosin at different NaCl concentrations
為了進(jìn)一步驗(yàn)證溶液體系中RA與巰基是否發(fā)生了相互作用,分析RA對(duì)肌球蛋白SDS-PAGE圖譜的影響。如圖4所示,肌球蛋白由大約200 kDa的肌球蛋白重鏈(myosin heavy chain,MHC)亞基譜帶和分子質(zhì)量大約15~25 kDa的3 條肌球蛋白輕鏈(myosin light chain,MLC)亞基,以及43 kDa的肌動(dòng)蛋白雜條帶組成。在SDS-PAGE中,非還原條件下(-β-ME),隨NaCl濃度的增加,兩組蛋白分子組成無(wú)明顯區(qū)別,在濃縮膠頂部均存在高分子聚合物。在還原條件下(+β-ME),這些聚合物得到分解,說(shuō)明聚合物主要是通過(guò)二硫鍵交聯(lián)產(chǎn)生。此外,MHC帶處濃度明顯減少,說(shuō)明非還原條件下,該條帶并非單純的MHC組分,還包含了部分由二硫鍵交聯(lián)形成的聚合物。與M組相比,還原和非還原條件下M-RA組圖譜均無(wú)明顯變化,也就是說(shuō)所有聚合物是通過(guò)二硫鍵交聯(lián)而非RA導(dǎo)致的共價(jià)交聯(lián)所產(chǎn)生,說(shuō)明肌球蛋白與RA之間作用以非共價(jià)作用為主,故在電泳條件下,SDS溶液可以破壞這些弱鍵,使得圖譜沒(méi)有明顯差異。
2.4 圓二色譜分析
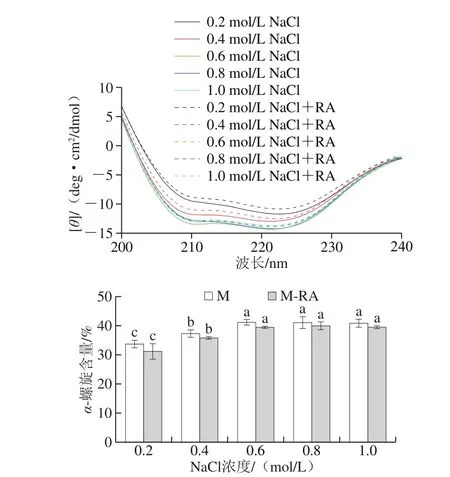
圖5 不同NaCl濃度下RA對(duì)肌球蛋白二級(jí)結(jié)構(gòu)的影響Fig. 5 Effect of RA on secondary structures of myosin at different NaCl concentrations
如圖5所示,兩組肌球蛋白在208 nm和222 nm波長(zhǎng)附近均出現(xiàn)2 個(gè)強(qiáng)負(fù)吸收峰,表明螺旋結(jié)構(gòu)的存在并作為二級(jí)結(jié)構(gòu)的主要部分,隨NaCl濃度增加,兩組α-螺旋結(jié)構(gòu)含量顯著增加(P<0.05),這可能是因?yàn)棣?螺旋結(jié)構(gòu)是由蛋白質(zhì)的羰基(—CO)和氨基(NH—)之間的氫鍵所維持的,高離子強(qiáng)度下受靜電屏蔽作用更有利于氫鍵的形成和穩(wěn)定。
總體而言,添加RA后,肌球蛋白的α-螺旋含量降低,說(shuō)明RA可以促進(jìn)肌球蛋白雙螺旋桿狀尾部結(jié)構(gòu)的解旋,導(dǎo)致肌球蛋白結(jié)構(gòu)進(jìn)一步得到展開(kāi),這與表面疏水性結(jié)果一致。此外,RA的羥基可與蛋白質(zhì)多肽鏈的羰基形成氫鍵,這可能會(huì)破壞肌球蛋白分子內(nèi)氫鍵平衡,從而破壞其穩(wěn)定性。
2.5 肌球蛋白與RA的互作機(jī)制
如圖6所示,在溶液中,肌球蛋白與RA之間主要通過(guò)疏水相互作用結(jié)合,不存在共價(jià)相互作用,添加RA可以促進(jìn)肌球蛋白結(jié)構(gòu)展開(kāi),暴露出更多的活性基團(tuán),進(jìn)一步利于肌球蛋白與RA之間發(fā)生疏水相互作用。肌球蛋白與RA之間的相互作用除了可以影響蛋白分子的結(jié)構(gòu)特征,還可能會(huì)進(jìn)一步影響其理化特性。
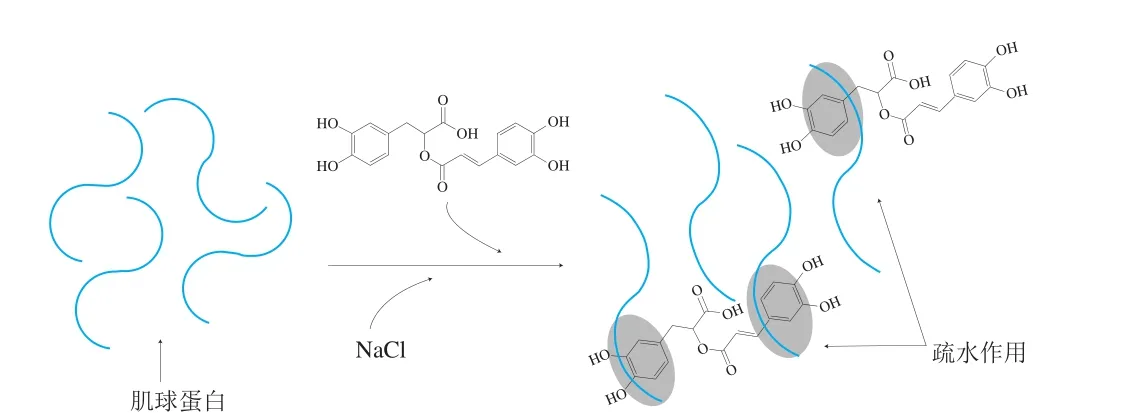
圖6 溶液中肌球蛋白與RA的互作機(jī)制Fig. 6 Mechanism underlying interaction between myosin and RA in aqueous solution
2.6 溶解度和濁度分析
如圖7所示,當(dāng)NaCl濃度從0.2 mol/L增加至0.6 mol/L時(shí),兩組的溶解度均呈顯著增加趨勢(shì)(P<0.05),而濁度呈現(xiàn)相反變化趨勢(shì),這是因?yàn)樵?.2 mol/L NaCl條件下,肌球蛋白分子以不溶性的纖絲形式存在;隨NaCl濃度增加,鹽離子逐漸破壞肌球蛋白分子間靜電相互作用,抑制絲狀體的形成,蛋白分子表現(xiàn)為可溶性單體或寡聚體[22]。當(dāng)NaCl濃度高于0.6 mol/L時(shí),M組溶解度呈現(xiàn)微弱的下降趨勢(shì),這可能是因?yàn)榉肿娱g的靜電斥力減弱允許蛋白分子相互靠近以致發(fā)生聚集,且由于疏水基團(tuán)的互作用,導(dǎo)致溶解度下降[20]。總體而言,添加RA后,肌球蛋白的溶解度降低,濁度增加,這可能是因?yàn)镽A促進(jìn)了肌球蛋白結(jié)構(gòu)展開(kāi),疏水性基團(tuán)得到暴露,并改變了蛋白分子的帶電狀況,導(dǎo)致蛋白聚集,從而改變了肌球蛋白的溶解度和濁度。
2.7 Zeta電位和粒徑分析
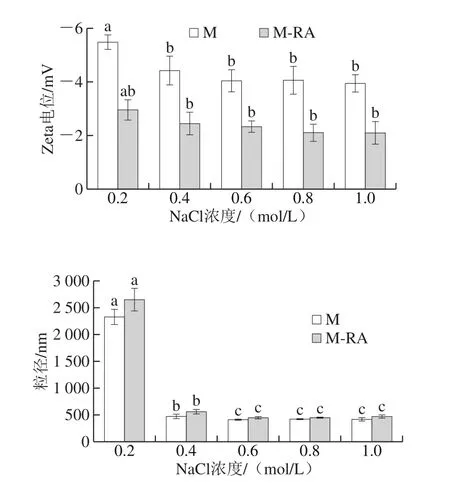
圖8 不同NaCl濃度下RA對(duì)肌球蛋白粒徑和Zeta電位的影響Fig. 8 Effects of RA on particle size and zeta potential of myosin at different NaCl concentrations
Zeta電位描述了溶液體系中顆粒之間的靜電相互作用,可以表征溶液體系的穩(wěn)定性[23-24]。如圖8所示,Zeta電位為負(fù)值,表明肌球蛋白呈負(fù)電荷。隨著NaCl濃度的增加,肌球蛋白Zeta電位的絕對(duì)值降低,這可能是由于鹽離子的引入對(duì)蛋白分子表面擴(kuò)散雙電層產(chǎn)生壓縮作用[25]。總體而言,添加RA后,Zeta電位的絕對(duì)值明顯降低,這可能是因?yàn)榧∏虻鞍着cRA結(jié)合后對(duì)電荷產(chǎn)生的的屏蔽作用,導(dǎo)致肌球蛋白表面負(fù)電荷數(shù)量減少,減弱了蛋白分子間的靜電斥力,使得蛋白分子之間傾向于相互聚集[26]。
粒徑反映了溶液體系中粒子的大小與分布,測(cè)定蛋白質(zhì)粒徑能直觀地看出蛋白質(zhì)聚集情況[27]。如圖8所示,與肌球蛋白濁度的變化結(jié)果一致,在0.2 mol/L NaCl下,肌球蛋白呈難溶性的細(xì)長(zhǎng)絲狀,蛋白分子聚集成團(tuán),故粒徑較大;而隨NaCl濃度的升高,聚集的蛋白分子經(jīng)歷先解聚再進(jìn)一步解離的過(guò)程,并以可溶性單體形式存在,粒徑顯著減小(P<0.05)。總體而言,添加RA后,蛋白粒徑增加,這可能是因?yàn)镽A促進(jìn)了肌球蛋白表面疏水性的升高,疏水作用是蛋白質(zhì)聚集現(xiàn)象的主要作用力,這種聚集在一定程度上影響了蛋白質(zhì)分子的粒徑分布。此外,肌球蛋白與RA之間的相互作用對(duì)肌球蛋白電荷的屏蔽作用降低了蛋白穩(wěn)定性,也會(huì)增大其粒徑。
2.8 乳化特性分析
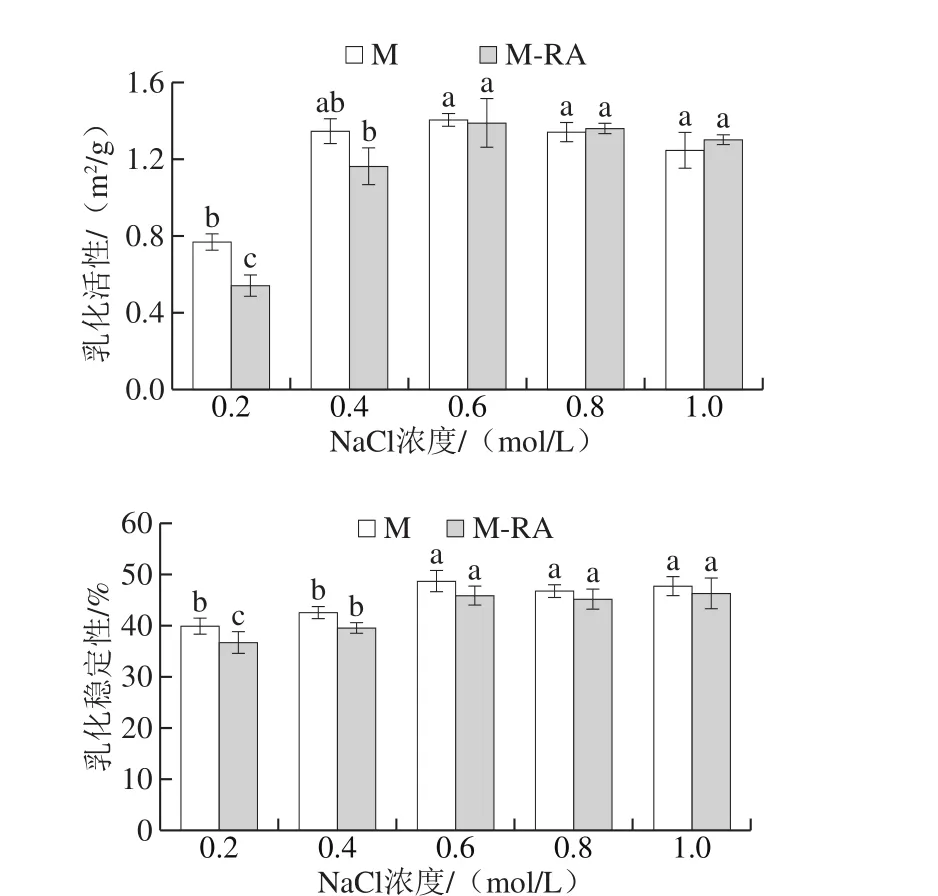
圖9 不同NaCl濃度下RA對(duì)肌球蛋白乳化活性和乳化穩(wěn)定性的影響Fig. 9 Effects of RA on EAI and ESI of myosin at different NaCl concentrations
在肉制品加工中,乳化性直接影響乳化肉制品的質(zhì)構(gòu)、黏著性、保油保水性和出品率,并對(duì)最終產(chǎn)品的品質(zhì)起決定性作用。如圖9所示,當(dāng)NaCl濃度從0.2 mol/L增加至0.6 mol/L時(shí),兩組乳化活性和乳化穩(wěn)定性均呈顯著增加趨勢(shì)(P<0.05),當(dāng)NaCl濃度高于0.6 mol/L時(shí),兩組乳化性和乳化穩(wěn)定性變化不明顯。溶解度是影響蛋白乳化性的重要因素,吳菊清等[28]研究表明蛋白乳化性與其溶解度呈正相關(guān),與表面疏水性呈負(fù)相關(guān)。隨NaCl濃度增加,肌球蛋白的溶解度逐漸增大,蛋白能迅速移到油水界面形成界面膜參與乳化,進(jìn)而提高了乳化活性;溶解度的增加還提高了分布在油水界面的蛋白質(zhì)濃度,蛋白可以更好地包裹在油滴表面,從而增強(qiáng)了乳化穩(wěn)定性。
添加RA后,0.2~0.4 mol/L NaCl條件下,乳化活性和乳化穩(wěn)定性明顯降低,這可能是因?yàn)榧∏虻鞍識(shí)A與的相互作用導(dǎo)致肌球蛋白溶解度降低,故乳化能力減弱。0.6~1.0 mol/L NaCl條件下,雖然RA降低了肌球蛋白的溶解度,但乳化性沒(méi)有明顯差別,乳化穩(wěn)定性差異也較小,這可能是因?yàn)镽A的添加改變了體系的親水親油平衡值,使蛋白更容易在油水界面展開(kāi),從而彌補(bǔ)了溶解度差異帶來(lái)的影響。曹云剛[29]研究發(fā)現(xiàn)低濃度表沒(méi)食子兒茶素沒(méi)食子酸酯(epigallocatechin gallate,EGCG)對(duì)肌原纖維蛋白乳化活性和乳化穩(wěn)定性無(wú)明顯影響,但高濃度EGCG顯著降低了肌原纖維蛋白的乳化活性和乳化穩(wěn)定性。故在實(shí)際應(yīng)用中,要考慮多酚類化合物或香辛料的種類及其使用量對(duì)產(chǎn)品功能性質(zhì)的影響[30],避免對(duì)產(chǎn)品品質(zhì)產(chǎn)生不良影響。
3 結(jié) 論
本研究分析了溶液狀態(tài)下肌球蛋白與RA之間的相互作用。色氨酸熒光結(jié)果表明,RA對(duì)肌球蛋白的內(nèi)源熒光具有較強(qiáng)的靜態(tài)猝滅作用,且RA與肌球蛋白主要通過(guò)疏水相互作用自發(fā)進(jìn)行結(jié)合。總巰基和電泳結(jié)果表明,肌球蛋白與RA之間不存在共價(jià)交聯(lián)。表面疏水性和圓二色譜結(jié)果表明,添加RA可以促進(jìn)肌球蛋白結(jié)構(gòu)展開(kāi),α-螺旋含量降低,更多活性基團(tuán)暴露,表面疏水性增加。不同NaCl條件下,添加RA會(huì)降低Zeta電位的絕對(duì)值,導(dǎo)致肌球蛋白的溶解度降低,濁度和粒徑增大。低鹽濃度下(0.2~0.4 mol/L),添加RA降低了肌球蛋白的乳化性,中高鹽濃度時(shí)(0.6~1.0 mol/L)時(shí),RA對(duì)肌球蛋白的乳化性影響較小。
