不同林齡油松根際土壤微生物群落多樣性及其影響因子
2020-07-08 00:39:28冀玉良
水土保持研究 2020年4期
丁 麗, 冀玉良, 李 懿
(1.包頭鐵道職業技術學院 建筑工程系, 內蒙古 包頭014060; 2.中國科學院 成都生物研究所, 成都 610041)
作為地球化學循環過程中的重要參與者,土壤微生物參與土壤中有機質分解、元素循環和土壤礦化等化學活動,能夠促進土壤物質循環、穩定與保持土壤的生產力和健康[1-2]。土壤生態系統如果想使功能、完整性和可持續性得到維持,必須要保持土壤微生物的多樣性。多年的研究發現,微生物的多樣性是增強微生物功能和生態系統穩定性的重要因素[3-5]。作為直接受植物根系和分泌物影響的土壤區域,根際土壤為土壤微生物與植物相互作用提供了場所[4],使微生物與根系之間保持了適應性協同進化和互惠的關系[6-8]。在植物根系活動的影響下,根際土壤的養分含量及其他土壤理化性質發生了變化,進一步使根際微生物群落的組成發生改變,非根際與根際土微生物群落組成和多樣性也出現了很大的差異。植物生長需要根際微生物的參與和作用,土壤營養物質能夠被促植物生長細菌(PGPR)的代謝活動進行分解、轉化,成為植物可吸收利用的形態后被吸收,有效地促進了植物的生長,而一些細菌分泌的激素能夠提高植物的適應性[9-10]。植物的種類、健康狀況、生長階段等因素會影響根際土壤微生物的群落結構。另外,如土壤營養狀況、pH值、溫度和水分等土壤理化性質會對微生物群落產生較大影響[11]。
植被生長除了需要一定的水分和養分之外,還需要合適的、有酶活性及微生物活動的土壤環境,能夠促進土壤活性及肥力。很多學者從土壤及微生物的關系角度,探究植被生長發育過程,發現微生物活動及其肥力的保持等規律,明確植被、土壤及微生物之間的關系[12-13],為實踐提供理論指導。但是,目前的研究中,研究岷江流域的根際土壤較少。因為該地區的土質較差,貧瘠沒有肥力,影響了植被的生長,造成水土流失較為嚴重,為保持該區域的水土[14-15],一般通過人工育林的方式進行鞏固,就對樹種的要求較高,否則人工林的生長發育會受到影響。油松(Pinustabulaefrmis)是一種松科常綠喬木,大多分布在我國的亞熱帶區域,具有良好的生長特性和環境適應性,作為人工育林樹種入選,表現優異,能夠有效地調節森林生態平衡,促進退耕還林工作的發展。油松的生長發育期較長,中間會出現林地肥力下降等問題[16-17]。這時,根際是油松與土壤進行能量交換與吸收的轉換器,生物活性較強,發揮了巨大的調節作用,通過分泌不同的有機質,平衡土壤中的營養元素,改善土壤結構,根際土壤特性變化比較明顯[18-19]。在此基礎上,本研究采用Illumina-MiSeq高通量測序技術,對不同林齡油松非根際與根際土壤中真菌18SrDNA基因V4區片段和細菌的16SrDNA基因V3—V4區片段做測序,發現和挖掘不同林齡油松非根際與根際土壤微生物群落多樣性及其與土壤環境因子之間的關系,為做好以油松為代表的人工育林工作提供參考和借鑒。
1 材料與方法
1.1 研究區概況
本試驗選定區域地處岷山南麓、長江流域最大的支流——岷江流域,具有非常復雜的地形地貌,擁有的干流超過735 km,并因地勢高低形成3 km左右的落差,上中游分界處為樂山、都江堰,上游區域的峽谷很多,地形復雜多變,并未形成大量成片的耕地,居住的人口非常少;中下游區域地勢比較平緩,大多是丘陵平原,利于人類居住,分布有大量的生活區和農業種植區。受經緯度影響,該區域為亞寒熱帶氣候,上游區域氣溫低于中下游氣溫,據多年氣象資料記載,該區域內夏季比較炎熱,最高溫接近39℃,冬季氣溫較低,最低溫為-4℃左右,出現在1月份,所以年均氣溫為9℃。該區域的無霜期接近半年,全年日照時長為2 100 h,雨季分明,土壤為紫色土。
1.2 試驗方法
2018年8月選取岷江上游的不同林齡的油松林(幼齡、中齡、成熟齡、過熟齡),每種類型油松林設置3個長、寬均為50 m的樣地,定期在每個樣地標記3株油松采集根際和非根際土壤樣品,其中根際土采用抖落法,即抖落大塊土壤,以根圍5 mm內的土壤作為根際土,根圍外5 mm土為非根際土壤,總共采集土壤樣品24份(根際土12份,非根際土12份);其中每一份根際土和非根際土分為3小份,一份用液氮保存,帶回實驗室后-80℃保存測定微生物群落結構;另外一部分車載冰箱4℃保存測定土壤微生物量碳氮和酶活性;第三部分常溫下風干處理用以測定土壤養分。
1.3 土壤養分和酶活性
采用滴定法、平板梯度稀釋法和比色法分別測定土壤脫氫酶、過氧化氫酶及脲酶,對有機碳、pH值、有效磷和有效氮等土壤養分[20],借助于《土壤農化分析》進行測定。
1.4 土壤總DNA提取
利用Fast DNA spin kit for soil 試劑盒(MP BIO,Inc.,Irvine,CA,USA)進行土壤DNA的提取,然后采用1%的瓊脂糖凝膠電泳,NanoDrop ND-1000微光分光光度計測定濃度。
1.5 土壤微生物基因組DNA的提取
往2 ml無菌離心管中滴入1 ml根際土壤溶液,于4℃,10 000 g離心30 s,棄上清,作為根際土壤微生物基因組DNA提取之用;往2 ml無菌離心管中滴入大約0.2 g非根際土壤(濕重),用無菌藥匙量取,作為提取基因組DNA之用。提取基因組DNA時使用OMEGA試劑盒E.Z.N.ATM Mag-Bind Soil DNA Kit試劑盒(OMEGA),檢測DNA的長度和完整性時使用1%瓊脂糖凝膠,檢測DNA的濃度和純度時使用NanoDrop。
1.6 DNA擴增及測序
對細菌16S rDNA V3—V4區進行擴增時,使用兩輪法。第一輪擴增,上游引物341F:CCCTACACGACG-ACGATCTTACAGATAGGGGGCGCAG,下游引物805R:GACTGGAGTTCCTTGGCACCCGAGAATTCCAG-ATACGGGTATCTAATCC,利用融合了barcode序列的通用引物進行;真菌18S rDNA擴增,第一輪擴增,上下游引物序列為NS1:CCTACACGACGATCCTATGCTTAACGTG,Fungi:GAAGTCCCTTATACGGGGATTTACCG-TAAACGT,利用融合了barcode 序列的通用引物進行。反應體系30 μl,包含各1 μl引物(10 μmol/L),15 μl 2×Taq master Mix(Thermo),模板DNA 20 ng。擴增條件為:94℃ 30 s,94℃ 3 min,65℃ 30 s,45℃ 20 s,擴增5個循環;94℃ 20 s,72℃ 30 s,55℃ 20 s,擴增20個循環,72℃延伸5 min。第二輪擴增,以第一輪的PCR產物為模板,使用Illumina 橋式PCR兼容引物。反應體系同上。擴增條件:95℃ 15 s,95℃ 30 s,72℃ 30 s,55℃ 15 s,擴增5個循環,72℃延伸5 min。PCR產物經磁珠法回收并定量。等量混合每個重復個體的PCR產物,使每份混合樣品中都包含全部重復個體,將混合樣品送至生工(上海)生物工程有限公司,使用Illumina-MiSeq平臺進行高通量測序。
1.7 數據分析
去除引物接頭序列和各低質量堿基(pH值red Quality Score=20)后,對測序原始數據進行拼接,去除長度短于200 bp的序列,非特異性擴增序列及嵌合體,就會得到每個樣本的有效序列數據劃定閾值為97%,針對18S和16S序列進行操作分類單元(operational taxonomic unit,OTU)的劃分。進行主坐標分析(PCoA)時,以樣本OTUs組成之間的weighted unifrac距離矩陣為依據,尋找微生物群落之間的相似性;對土壤養分和微生物群落之間進行冗余分析(Reundancy analysis,RDA)時,采用Canoco 4.5軟件。
用SPSS 18.0和Excel 2007進行方差檢驗和數據統計,為檢驗各指標之間的相關性,可以通過單因素方差分析(One-way ANOVA),平均值±標準誤差表示(Mean±SE)和Pearson相關系數法實現。
2 結果與分析
2.1 不同林齡油松根際與非根際土壤養分
從表1中看出,不同林齡油松樣地,土壤中根際土壤pH值低于非根際土壤,說明由于根在土壤中長期生長,過程中不斷向根際分泌出有機化合物,油松林的土壤pH值處于弱酸性,屬較強酸性土壤,隨著林齡的增大,pH值也逐漸增加。土壤中根際和非根際有機碳含量隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,其中幼林、中林和成熟林根際有機碳含量顯著高于非根際(p<0.05),過熟林根際有機碳含量與非根際差異并不顯著(p>0.05);根際土壤有機碳含量在幼林、中林、成熟林和過熟林分別高于非根際15.98%,17.63%,10.81%,15.31%。土壤中根際和非根際全氮含量隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,其中幼林、中林和成熟林根際全氮含量顯著高于非根際(p<0.05),過熟林根際全氮含量與非根際差異并不顯著(p>0.05);根際全氮含量在幼林、中林、成熟林和過熟林分別高于非根際13.95%,12.39%,11.57%,3.33%。土壤中根際和非根際有效磷含量隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,其中幼林、中林和成熟林根際有效磷含量顯著高于非根際(p<0.05),過熟林根際有效磷含量與非根際差異并不顯著(p>0.05);根際有效磷含量在幼林、中林、成熟林和過熟林分別高于非根際16.98%,10.90%,24.28%,1.81%。土壤中根際和非根際有效鉀含量隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,其中幼林、中林和成熟林根際有效鉀含量顯著高于非根際(p<0.05),過熟林根際有效鉀含量與非根際差異并不顯著(p>0.05);根際有效鉀含量在幼林、中林、成熟林和過熟林分別高于非根際20.23%,12.59%,10.14%,6.37%。

表1 不同林齡油松根際與非根際土壤養分
注:*表示根際與非根際差異顯著(p<0.05),下表同。
2.2 不同林齡油松根際與非根際土壤酶活性
由表2可知,不同林齡油松表現出各種酶類的活性,其根際土壤高于非根際土壤。土壤中根際和非根際蔗糖酶活性隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,其中幼林、中林和成熟林根際蔗糖酶活性顯著高于非根際(p<0.05),過熟林根際蔗糖酶活性與非根際差異并不顯著(p>0.05);根際蔗糖酶活性在幼林、中林、成熟林和過熟林分別高于非根際28.00%,33.73%,27.66%,12.48%。土壤中根際和非根際堿性磷酸酶活性隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,其中幼林、中林和成熟林根際堿性磷酸酶活性顯著高于非根際(p<0.05),過熟林根際堿性磷酸酶活性與非根際差異并不顯著(p>0.05);根際堿性磷酸酶活性在幼林、中林、成熟林和過熟林分別高于非根際23.53%,12.62%,39.44%,4.69%。土壤中根際和非根際脲酶活性隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,其中幼林、中林和成熟林根際脲酶活性顯著高于非根際(p<0.05),過熟林根際脲酶活性與非根際差異并不顯著(p>0.05);根際脲酶活性在幼林、中林、成熟林和過熟林分別高于非根際20.97%,41.83%,16.43%,8.47%。土壤中根際和非根際過氧化氫酶活性隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,其中幼林、中林和成熟林根際過氧化氫酶活性顯著高于非根際(p<0.05),過熟林根際過氧化氫酶活性與非根際差異并不顯著(p>0.05);根際過氧化氫酶活性在幼林、中林、成熟林和過熟林分別高于非根際27.57%,14.73%,9.60%,0.31%。

表2 不同林齡油松根際與非根際土壤酶活性 mg/g
2.3 不同林齡油松土壤微生物群落多樣性
對于土壤細菌多樣性,由表3可知,油松根際和非根際土壤細菌覆蓋度、豐富度指數、Shannon-Wiener指數、均勻度指數、ACE、Chao 1和Simpson指數隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,過熟林有所降低。差異性檢驗表明,油松根際土壤細菌豐富度指數、Shannon-Wiener指數、ACE、Chao 1指數均顯著高于非根際(p<0.05);而根際土壤細菌覆蓋度、均勻度和Simpson指數與非根際差異并不顯著(p>0.05)。
對于土壤真菌多樣性,由表4可知,油松根際和非根際土壤真菌覆蓋度、豐富度指數、Shannon-Wiener指數、均勻度指數、ACE、Chao 1和Simpson指數隨著油松林齡的增長表現出先增加后降低趨勢,在成熟林階段達到最大值,過熟林有所降低。差異性檢驗表明,油松根際土壤真菌豐富度指數、Shannon-Wiener 指數、ACE、Chao 1指數均顯著高于非根際(p<0.05);而根際土壤細菌覆蓋度、均勻度和Simpson指數與非根際差異并不顯著(p>0.05)。

表3 油松根際與非根際土壤細菌多樣性
注:同列不同小寫字母表示不同林齡差異顯著(p<0.05),下表同。

表4 油松根際與非根際土壤真菌多樣性
2.4 不同林齡油松土壤微生物群落組成
由圖1可知,不同林齡油松根際土壤細菌群落中,優勢類群主要是變形菌門(36.29%)、酸桿菌門(28.92%)和芽單胞菌門(7.63%),還包括浮霉菌門(5.19%)、放線菌門(4.36%)、擬桿菌門(0.57%)、后壁菌門(4.19%)、綠彎菌門(5.23%)、疣微菌門(4.07%)。非根際土壤細菌群落中,優勢類群主要是變形菌門(46.23%)、酸桿菌門(21.47%)和芽單胞菌門(11.53%)。其中根際土壤細菌酸桿菌門相對豐度高于非根際,變形菌門相對豐度低于非根際。
油松根際土壤真菌群落中,優勢類群主要是子囊菌門(56.39%)和擔子菌門(24.13%),還包括結合菌門(5.02%)、壺菌門(0.24%)、新麗鞭毛菌門(6.57%)、球囊菌門(2.01%)、芽枝菌門(2.19%)。非根際土壤真菌群落中,優勢類群主要是子囊菌門(48.23%)和擔子菌門(29.56%),其中根際土壤細菌酸桿菌門相對豐度高于非根際,變形菌門相對豐度低于非根際。
主坐標分析(PCoA)分析表明:油松土壤根際和非根際細菌群落具有很好的相似性,并且根際和非根際細菌群落產生明顯的分離效應;油松土壤根際和非根際真菌群落具有很好的相似性,并且根際和非根際真菌群落產生明顯的分離效應(圖2)。
2.5 土壤養分、酶活性與微生物群落結構的相關性
相關性分析表明(表5):油松土壤細菌Coverage,ACE與土壤養分均沒有顯著的相關性(p>0.05);土壤pH值與土壤細菌多樣性呈負相關,土壤有效磷、有效鉀和蔗糖酶活性均沒有顯著的相關性(p>0.05)。土壤有機碳與細菌Richness,Shannon呈極顯著的正相關(p<0.01),與細菌Evenness,Chao 1和Simpson呈顯著的正相關(p<0.05);土壤全氮與細菌Richness,Shannon呈極顯著的正相關(p<0.01),與細菌Chao 1和Simpson呈顯著的正相關(p<0.05);堿性磷酸酶活性與Shannon,Chao 1和Simpson呈極顯著的正相關(p<0.01),與Evenness呈顯著的正相關(p<0.05);脲酶活性與Shannon,Evenness和Chao 1呈極顯著的正相關(p<0.01),與Simpson呈顯著的正相關(p<0.05);過氧化氫酶活性與Shannon,Evenness,Chao 1 Simpson呈極顯著的正相關(p<0.01)。
油松土壤真菌Coverage,ACE與土壤養分均沒有顯著的相關性(p>0.05);土壤pH值與土壤真菌多樣性呈負相關,土壤有效磷、有效鉀和蔗糖酶活性均沒有顯著的相關性(p>0.05)。土壤有機碳與真菌Shannon,Evenness,Chao 1呈顯著的正相關(p<0.05);土壤全氮與Shannon呈極顯著的正相關(p<0.01),與Richness,Chao 1和Simpson呈顯著的正相關(p<0.05);堿性磷酸酶活性與Shannon,Evenness,Chao 1和Simpson呈顯著的正相關(p<0.05);脲酶活性與Shannon,Evenness,Chao 1和Simpson呈顯著的正相關(p<0.05);過氧化氫酶活性與Shannon,Evenness,Chao 1和Simpson呈顯著的正相關(p<0.05)。

圖1 油松根際和非根際土壤微生物群落組成
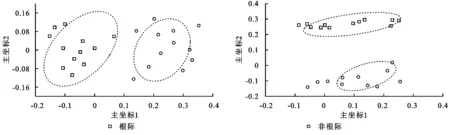
圖2 基于Weighted unifrac 距離的土壤微生物群落的主坐標分析(PCoA)
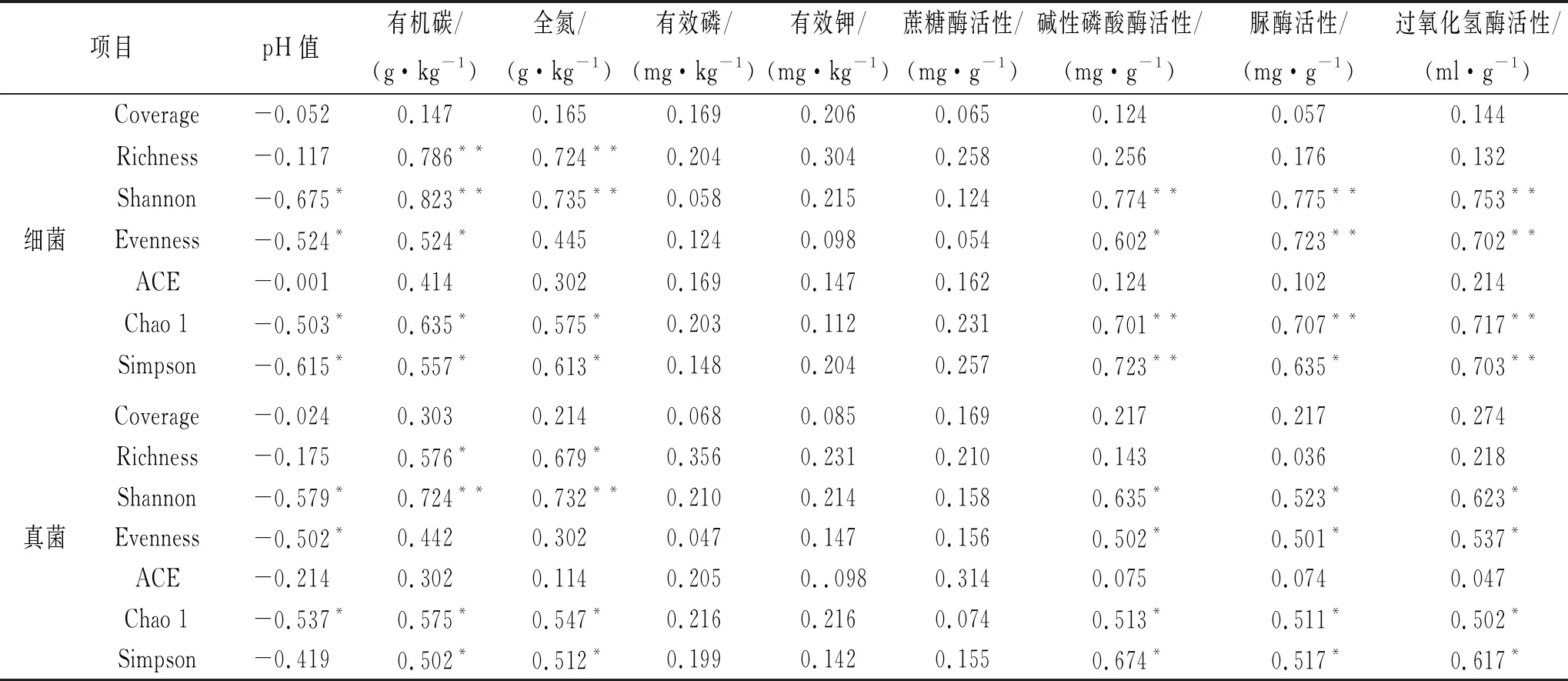
表5 土壤養分、酶活性與土壤微生物群落多樣性相關性
注:*,**分別表示在0.05,0.01水平上差異顯著。
2.6 土壤微生物群落的RDA分析
蒙特卡羅置換檢驗結果顯示,第一典范軸p<0.05(F=36.59),所有典范軸的p值為0.913,表明該排序模型的解釋變量(即土壤環境因子)與響應變量(即土壤細菌和真菌群落結構)的變化關系較為顯著。油松根際土壤細菌和真菌群落與土壤環境因子的冗余分析結果見表6。對于土壤細菌群落,RDA前兩個排序軸的特征值分別為0.652 4,0.211 7,分別解釋了65.24%,21.17%的細菌多樣性變化。本文所選的9個土壤環境因子共解釋了86%的總特征值,對油松根際土壤細菌群落有顯著影響。其中對土壤細菌多樣性影響較大的有SOC和STN,而pH值對土壤細菌多樣性影響為負。對于土壤真菌群落,RDA前兩個排序軸的特征值分別為0.606 8,0.235 6,分別解釋了60.68%,23.56%的真菌多樣性變化。本文所選的9個土壤環境因子共解釋了86%的總特征值,對油松根際土壤真菌多樣性有顯著影響。其中對土壤真菌多樣性影響較大的有SOC和STN,而pH值對土壤真菌多樣性影響為負(圖3)。

表6 冗余分析結果
3 討論與結論
水分及養分是大多數植被所賴以生存的生長發育物質,能夠保證其獲得能量和轉換,并影響最終的存活。在這個階段中,微生物的活動至關重要,承擔了陸地有機質分解者的角色,凋落物等腐殖質通過微生物作用得到分解或降解,轉化成植被可以吸收的能量和養分,從而促進養分的礦化及分解活動。微生物是土壤生態的重要組成部分,其數量及新陳代謝的變化能夠直接作用于土壤,使其結構發生變化,并在一定程度上體現了土壤的肥力及活性。眾所周知,林木生長階段存在不同現象,同樣的,微生物多樣性也會變化[21-22]。一般而言,油松非根際(p<0.05)土壤pH值明顯高于根際,非根際(p<0.05)土壤的全氮、有機碳、有效磷和有效氮均低于根際,非根際和根際土壤中的全氮、有機碳、有效磷和有效氮含量的變化區間一直,即伴隨著油松林的增長,呈現先增加后降低的趨勢,生長至成熟林時,處于最高值。非根際土壤中微生物多樣性和酶活性(堿性磷酸酶活性、脲酶活性、蔗糖酶活性、過氧化氫酶活性)明顯低于根際,兩者的土壤酶活性變化曲線一致,隨著油松林齡的增長,呈現先增加后降低狀況,并在成熟林階段處于最大值,過熟林時開始降低,這反映了根際聚集效應,表明土壤中的有效鉀、有效磷含量較高。但是對于幼齡油松來說,其根系并未發育完善,根產物較少,所以主要通過腐殖質分解獲得有機質;對于中齡林和成熟林來說,可以通過根產物收獲大量的有機質。隨著油松林齡的增加,土壤酶活性會呈現“低—高—低”的狀態,先降后升,于中齡時達到最高點后再開始下降。研究顯示,根際酶活性遠遠高于非根際酶活性[23-24],油松根系分泌的酶進入土壤會形成極強的酶活性。在微生物作用下,根際的作用范圍縮小,微生物分布數量及養分在外界環境變化的情況下,會改變內部結構,直接影響酶的分泌,導致不同土壤區域的酶活性出現較大差異。對于根際或非根際土壤來說,在林齡不斷增加的前提下,過氧化氫酶、脫氫酶、脲酶的活性的變化特點為先降后升,并在一定情景下呈現下降趨勢,即“低—高—低”的狀態。這說明,隨著樹齡的變化,油松對土壤的作用會發生變化和差異,中齡的油松根系活力更強,能夠通過產生更多的分泌物,加速有機質降解,提高微生物活動效率,提高能量轉化速度。
土壤和植被具有密不可分的關系,前者為后者生存提供載體和場所,后者能反過來影響和制約前者,土壤中的養分能夠促進植被的生長發育,微生物則通過降解凋落物等腐殖質,將其轉化為能夠得到利用吸收的養分[16-18],能夠幫助土壤保持有效的肥力。另外,在這個階段中,酶所起的作用也不容忽視,能夠促進有機質分解,加速土壤的生化反應,提高土壤活性。經研究可知,油松土壤非根際和根際真菌群落和細菌的相似性很高,這兩種細菌群落會產生較顯著的分離效應。據分析相關資料可知,油松土壤的細菌和真菌多樣性與有效鉀、有效磷和蔗糖酶活性不存在明顯的相關性(p>0.05),油松土壤真菌Coverage,ACE和細菌與土壤養分不存在明顯的相關性(p>0.05),微生物的生長特點,決定了過高的土壤pH值對微生物生長和新陳代謝不利,養分及酶活性則對微生物分布多樣性具有有利影響,這導致了微生物分布在不同的土壤中會出現較大的差別。在不同的發育階段,油松對土壤的養分需求會發生變化,也反映在微生物的變化和根際土壤變化方面,凸顯了根際的作用。整體來看,根際土壤中分布的微生物數量明顯多于非根際土壤。但是油松的生長發育會受到很多因素的影響,根際及非根際土壤還會受到季節更迭的影響,所以這些也需要學界在今后加強具有針對性的研究[25-27]。
土壤微生物群落多樣性和組成,受到土壤理化性質和不同生境之間的差異影響,在不同的生境中,根際微生物群落存在很大差別。另外,油松土壤的非根際和根際真菌群落及細菌具有較強的一致性,非根際和根際細菌群落存在顯著的分離效應。經分析發現:油松土壤細菌和真菌多樣性與土壤pH值存在負比例關系(p>0.05),油松土壤真菌Coverage,ACE和細菌與土壤養分不存在顯著的相關性(p>0.05);土壤細菌和真菌多樣性與土壤電導率和全磷均不存在顯著的相關性(p>0.05)。冗余分析顯示:真菌82%和細菌86%的總特征值由9個土壤環境因子分別進行解釋,可知油松土壤細菌和真菌多樣性收到了土壤環境因子的影響。其中SOC和STN對土壤細菌和真菌多樣性影響較大,而土壤細菌和真菌多樣性受到pH值的影響為負。
