自噬核心蛋白ATG101對米色脂肪細(xì)胞分化和產(chǎn)熱的影響
2020-07-23 03:26:58馬靜遠(yuǎn)李少博楊迪白寧寧米日阿依阿里木江楊穎韓峻峰
江蘇大學(xué)學(xué)報(醫(yī)學(xué)版) 2020年4期
馬靜遠(yuǎn), 李少博, 楊迪, 白寧寧, 米日阿依·阿里木江, 楊穎, 韓峻峰
(上海交通大學(xué)附屬第六人民醫(yī)院內(nèi)分泌代謝科, 上海 200233)
脂肪組織是調(diào)節(jié)機(jī)體能量穩(wěn)態(tài)的關(guān)鍵器官。當(dāng)其功能失調(diào),無法適當(dāng)擴(kuò)展以存儲多余的能量時,會導(dǎo)致胰島素抵抗以及異位脂肪沉積[1]。哺乳動物體內(nèi)主要存在白色脂肪細(xì)胞和棕色脂肪細(xì)胞。其中,白色脂肪細(xì)胞主要存儲多余的三酰甘油,而棕色脂肪細(xì)胞則有助于能量利用和產(chǎn)熱[2-4]。近年來研究發(fā)現(xiàn)小鼠皮下白色脂肪組織中還存在第三類脂肪細(xì)胞—米色脂肪細(xì)胞:在基礎(chǔ)條件下分子和形態(tài)特征與白色脂肪細(xì)胞相似,但是當(dāng)暴露于低溫或受到β-腎上腺素刺激時,可以表達(dá)棕色脂肪細(xì)胞的標(biāo)志性基因[5-7],如解偶聯(lián)蛋白1(uncoupling protein 1,Ucp1),PR結(jié)構(gòu)域蛋白16(PR domain-containing 16,Prdm16)等。米色脂肪細(xì)胞的基本特征是可以響應(yīng)外部刺激而對產(chǎn)熱基因程序產(chǎn)生動態(tài)調(diào)節(jié)。通過遺傳和藥理學(xué)方法選擇性激活米色脂肪細(xì)胞的產(chǎn)熱作用,可以緩解肥胖和胰島素抵抗[8-9]。研究表明自噬誘導(dǎo)的線粒體更新對于體內(nèi)米色脂肪細(xì)胞的功能維持和能量消耗至關(guān)重要[10-11]。
自噬相關(guān)基因101(autophagy related gene 101,ATG101)于2009年被兩個獨立的課題組分別發(fā)現(xiàn)并命名[12-13],在哺乳動物肝臟、脂肪、心臟、腦、腸等組織和器官中廣泛表達(dá)。編碼人類ATG101的基因定位于染色體12q13.13,cDNA全長7 573 bp,編碼蛋白含218個氨基酸。ATG101的氨基酸鏈N端有一HORMA結(jié)構(gòu)域,可以與自噬相關(guān)基因13(autophagy related gene 13,ATG13)的C端HORMA結(jié)構(gòu)域形成穩(wěn)定的異二聚體[14-15]。ATG101和ATG13的穩(wěn)定結(jié)合有助于維持unc-51樣激酶1(unc-51 like kinase 1,ULK1)自噬起始復(fù)合體的穩(wěn)定性,該復(fù)合物在自噬的啟動階段可以響應(yīng)營養(yǎng)變化,并將其他下游ATG蛋白募集到自噬體結(jié)構(gòu)中[16-18],因此,ATG101對自噬過程的正常進(jìn)行十分關(guān)鍵。自噬對脂肪組織中營養(yǎng)代謝變化高度敏感,而ATG101在脂肪細(xì)胞中的作用尚未得到充分探索。本研究以小鼠的血管基質(zhì)細(xì)胞(stromal vascular fraction,SVF)為研究模型,通過慢病毒介導(dǎo)shRNA敲減Atg101基因,在細(xì)胞水平上觀察ATG101在米色脂肪細(xì)胞增殖、分化中的作用,并探究其對米色脂肪細(xì)胞產(chǎn)熱功能的影響,以期為深入研究ATG101在脂肪組織重塑中的作用提供理論線索。
1 材料與方法
1.1 主要材料
12只6周齡雄性C57BL/6J小鼠(上海斯萊克實驗動物有限公司);DMEM、胎牛血清(美國Gibco公司);3-異丁基-1-甲基黃嘌呤、地塞米松、羅格列酮、三碘甲狀腺原氨酸、膠原酶(美國Sigma公司);胰島素(常規(guī)優(yōu)泌林,美國Lilly公司);Trizol試劑(美國Invitrogen公司);RIPA裂解液、蛋白酶抑制劑PMSF和BCA蛋白定量試劑盒(上海碧云天公司);油紅O儲液(美國Sigma公司);引物(上海生物工程有限公司);逆轉(zhuǎn)錄和SYBR Premix ExTaq(南京Vazyme公司);兔源性ATG101單克隆抗體,鼠源性β-微管蛋白單克隆抗體(美國Cell Signaling Technology公司);兔源性UCP1多克隆抗體(美國Abcam公司);鼠源性GAPDH多克隆抗體(上海康城生物公司)。
1.2 方法
1.2.1 SVF向米色脂肪細(xì)胞誘導(dǎo)分化 6只6周齡C57BL/6J雄性小鼠過量麻醉處死,取皮下脂肪,緩沖液反復(fù)沖洗后,剪至肉糜狀,膠原酶37℃振蕩消化30 min,冰上放置20 min后2 400 r/min離心10 min。棄去上清液,加入DMEM后再次離心去除上清液,所得沉淀即為SVF,加入DMEM吹打均勻后鋪板。
用含10%胎牛血清的DMEM培養(yǎng)SVF細(xì)胞,細(xì)胞生長至接觸抑制后,加入誘導(dǎo)液A(10%胎牛血清的DMEM含0.5 mmol/L 3-異丁基-1-甲基黃嘌呤、1.7 μmol/L胰島素、1 μmol/L地塞米松、10 μmol/L羅格列酮、10 nmol/L三碘甲狀腺原氨酸),隔天更換誘導(dǎo)液B(10%胎牛血清的DMEM含1.7 μmol/L胰島素、10 μmol/L羅格列酮、10 nmol/L三碘甲狀腺原氨酸),之后繼續(xù)使用誘導(dǎo)液B培養(yǎng),直至出現(xiàn)大量脂滴。
1.2.2 慢病毒侵染敲減Atg101 含有shATG101載體的慢病毒購自上海吉凱基因。使用感染復(fù)數(shù)(MOI)35侵染SVF,陰性對照病毒作為對照組。侵染48 h后在熒光顯微鏡下觀察綠色熒光蛋白陽性的細(xì)胞數(shù)量來評估感染效率,通過實時定量PCR和蛋白質(zhì)印跡評估Atg101的敲減效率。確定敲減成功后,進(jìn)行細(xì)胞傳代并誘導(dǎo)分化為成熟米色脂肪細(xì)胞,用于之后的實驗。
1.2.3 實時定量PCR測定米色脂肪細(xì)胞成脂和產(chǎn)熱標(biāo)志基因mRNA表達(dá) 分別收取誘導(dǎo)分化不同時間的細(xì)胞,使用Trizol試劑抽提RNA,經(jīng)逆轉(zhuǎn)錄成為cDNA,并由PCR試劑盒進(jìn)行擴(kuò)增。引物序列如下:Atg101上游5′-ATGAACTGTCGATCAGAAGTGC-3′,下游5′-CCTATGGAGTACGTGCCCT-3′;Ucp1上游5′-AGGCTTCCAGTACCATTAGGT-3′,下游5′-CTGAGTGAGGCAAAGCTGATTT-3′;脂肪酸結(jié)合蛋白4(Fabp4)上游5′-AAGGTGAAGAGCATCATAACCCT-3′,下游5′-TCACGCCTTTCATAACACATTCC-3′;Prdm16上游5′-CAGCACGGTGAAGCCATTC-3′,下游5′-GCGTGCATCCGCTTGTG-3′;葡萄糖轉(zhuǎn)運體4(Glut4)上游5′-GTGACTGGAACACTGGTCCTA-3′,下游5′-CCAGCCACGTTGCATTGTAG-3′;誘導(dǎo)細(xì)胞死亡DNA片斷化因子α樣效應(yīng)因子(Cidea)上游5′-TGACATTCATGGGATTGCAGAC-3′,下游5′-CATGGTTTGAAACTCGAAAAGGG-3′;36B4上游5′-AAGCGCGTCCTGGCATTGTCT-3′,下游5′-CCGCAGGGGCAGCAGTGGT-3′。PCR反應(yīng)體系10 μL:cDNA 1 μL,上下游引物各0.4 μL,SBYR Premix ExTaq5 μL,去離子水3.2 μL。PCR反應(yīng)條件:95 ℃預(yù)變性30 s,95 ℃變性5 s,60 ℃退火30 s,72 ℃延伸15 s,共45個循環(huán)。以36B4作為內(nèi)參,計算目的基因mRNA的相對拷貝數(shù)2-△△Ct。
1.2.4 蛋白質(zhì)印跡法檢測ATG101、UCP1蛋白的表達(dá) 細(xì)胞總蛋白采用RIPA裂解液提取,經(jīng)BCA蛋白定量試劑盒測蛋白濃度,最后95℃加熱變性。取10 μg蛋白樣本進(jìn)行SDS-PAGE,將蛋白濕轉(zhuǎn)至硝酸纖維素膜,用5%脫脂牛奶(TBST配制)封閉1 h,一抗稀釋比例均為1 ∶ 1 000,4 ℃孵育過夜。次日TBST洗3次,每次10 min,辣根過氧化物酶標(biāo)記的抗兔(鼠)二抗按1 ∶ 2 000比例稀釋,室溫孵育1 h后TBST洗3次,每次10 min。最后用Image Quant凝膠圖像分析儀成像。
1.2.5 油紅O脂滴染色 敲減Atg101后分化第6天的米色脂肪細(xì)胞棄掉培養(yǎng)基后,用預(yù)冷的PBS沖洗細(xì)胞2次,然后用4%多聚甲醛固定15 min,用PBS洗滌2次后,稀釋好的油紅O工作液室溫孵育20 min,用雙蒸水沖洗3次,然后直接用顯微鏡(Nikon)觀察。
1.3 統(tǒng)計學(xué)處理
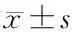
2 結(jié)果
2.1 ATG101的表達(dá)隨米色脂肪細(xì)胞分化逐漸增加
在SVF細(xì)胞誘導(dǎo)分化為成熟米色脂肪細(xì)胞的過程中,實時定量PCR結(jié)果顯示,Atg101mRNA表達(dá)水平從分化第2天起逐漸增加,并在分化第8天達(dá)到最高,與分化第0天相比,升高了約6倍(圖1A)。分化過程中細(xì)胞裂解物的蛋白質(zhì)印跡結(jié)果表明,ATG101的蛋白表達(dá)水平在分化過程中逐漸增加(圖1B)。
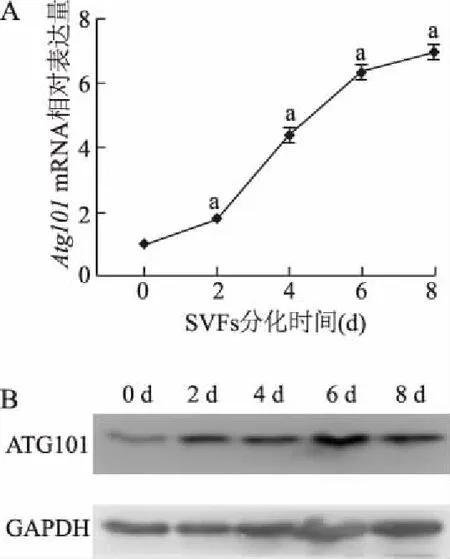
A:米色脂肪細(xì)胞分化過程中Atg101 mRNA表達(dá);B:米色脂肪細(xì)胞分化過程中ATG101蛋白的表達(dá)。a:P<0.01,與0 d比較
2.2 Atg101敲減抑制米色脂肪細(xì)胞的成脂分化
慢病毒介導(dǎo)的shRNA在米色脂肪細(xì)胞分化過程中有效地降低了ATG101的表達(dá),見圖2。
分化第6天的米色脂肪細(xì)胞油紅O染色顯示,ATG101下調(diào)后導(dǎo)致分化的米色脂肪細(xì)胞中脂質(zhì)積累減少(圖3A)。在分化第6天,實時定量PCR的結(jié)果表明,成熟米色脂肪細(xì)胞標(biāo)志基因Fabp4的表達(dá)降低至對照組的0.57,Cidea降低至0.46,而Glut4降低至0.68(P<0.01和P<0.05,圖3B)。因此,ATG101在細(xì)胞水平上參與前脂肪細(xì)胞向成熟米色脂肪細(xì)胞的分化成脂過程。

A:蛋白質(zhì)印跡; B:實時定量PCR;C:Atg101敲減后米色脂肪細(xì)胞分化第6天的綠色熒光(×100倍)。
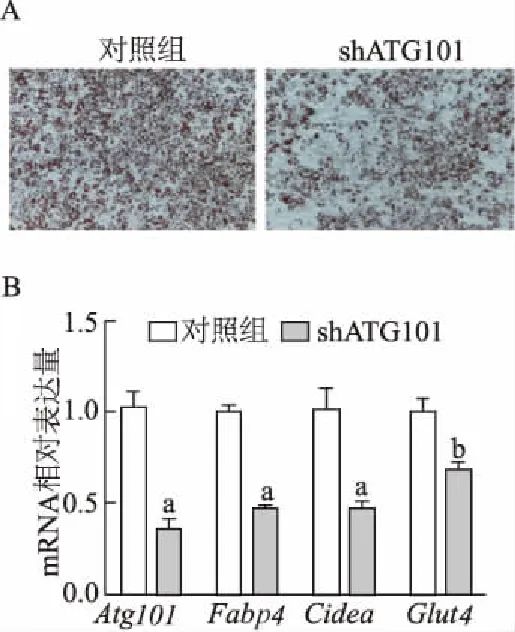
A:Atg101敲減后米色脂肪細(xì)胞分化第6天的油紅O染色(×100倍);B:Atg101敲減后成熟米色脂肪細(xì)胞標(biāo)志基因的mRNA表達(dá)。a:P<0.01,b:P<0.05,與對照組比較
2.3 Atg101敲減抑制脂肪細(xì)胞產(chǎn)熱過程
實時定量PCR結(jié)果顯示,Atg101下調(diào)后在米色脂肪細(xì)胞分化早期Prdm16的表達(dá)降低,成熟棕色脂肪細(xì)胞標(biāo)志基因Cidea和Ucp1顯著下調(diào)。在敲減Atg101后,UCP1蛋白水平也明顯下調(diào),與mRNA變化一致,見圖4。上述結(jié)果初步表明,下調(diào)Atg101可以部分影響小鼠皮下脂肪組織中米色脂肪細(xì)胞的產(chǎn)熱過程。
3 討論
自噬作為真核細(xì)胞中主要的降解途徑,可以清除多余和受損的細(xì)胞器,回收利用細(xì)胞質(zhì),以適應(yīng)營養(yǎng)條件的變化并參與維持細(xì)胞穩(wěn)態(tài)[19-20]。之前關(guān)于自噬的研究主要集中在神經(jīng)退行性疾病和腫瘤等方面,自噬與能量代謝的關(guān)系鮮有涉及。近來有研究指出脂肪分化過程可能是代表自噬活性極高的生理過程之一。在脂肪細(xì)胞誘導(dǎo)分化后的第6天,電子顯微鏡下的超微結(jié)構(gòu)顯示,分化細(xì)胞的胞質(zhì)體積中有5%以上是自噬體[21]。自噬與脂肪細(xì)胞分化和功能密切相關(guān),Singh等[22]發(fā)現(xiàn)在3T3-L1前脂肪細(xì)胞敲除自噬相關(guān)基因Atg7后,脂質(zhì)合成減少、自噬功能抑制、脂肪細(xì)胞分化相關(guān)蛋白表達(dá)減少;而Baerga等[23]通過敲除自噬相關(guān)基因Atg5后得到了類似的結(jié)果,提示自噬參與脂肪細(xì)胞的正常分化過程。因此,自噬在脂肪細(xì)胞的分化、脂質(zhì)代謝等方面發(fā)揮著重要作用。本研究結(jié)果表明ATG101在米色脂肪細(xì)胞分化期間表達(dá)增加,在SVF細(xì)胞敲減Atg101并誘導(dǎo)分化后,米色脂肪細(xì)胞的分化明顯受損,脂質(zhì)沉積減少。其確切機(jī)制尚不清楚,推測可能是Atg101的缺失導(dǎo)致了ULK1自噬起始復(fù)合物形成抑制,自噬活動清除多余或受損細(xì)胞器的功能部分受損,從而影響了脂肪細(xì)胞分化過程。
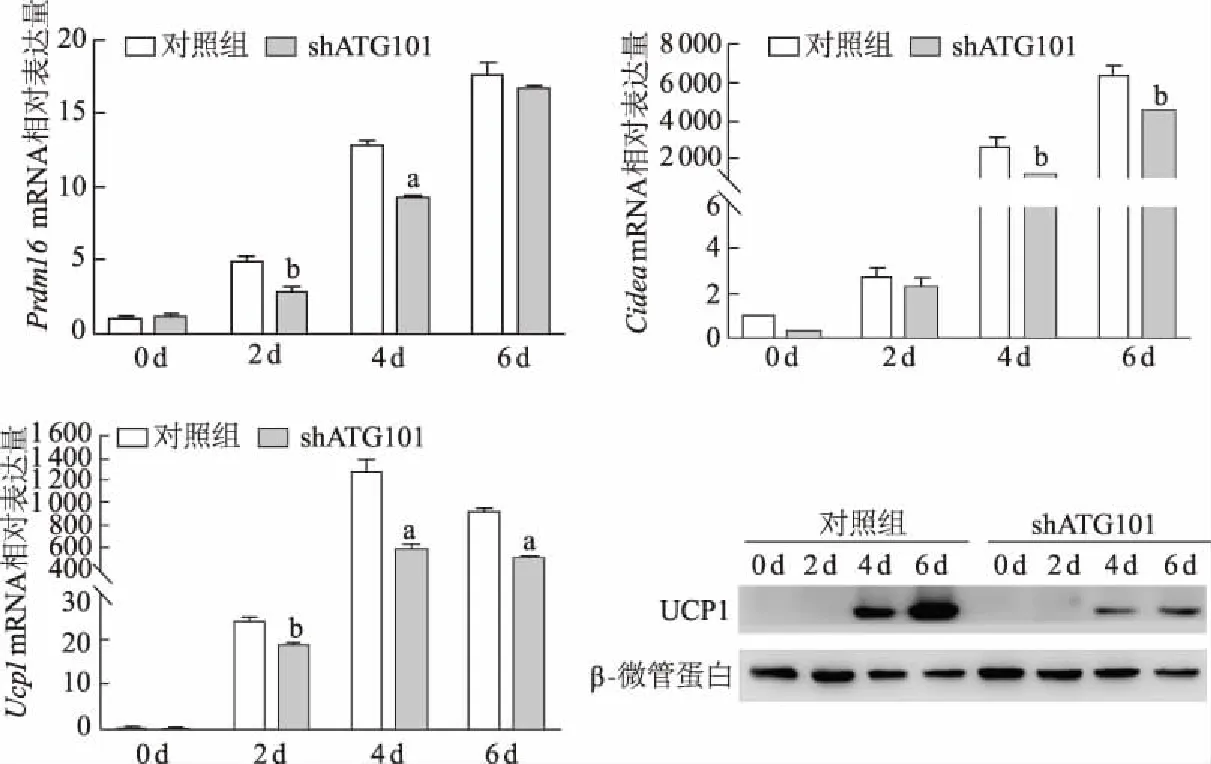
a:P<0.01,b:P<0.05,與對照組比較
白色脂肪細(xì)胞肥大、增生和(或)米色、棕色脂肪細(xì)胞萎縮、發(fā)育不全會導(dǎo)致脂肪因子分泌、游離脂肪酸釋放、脂質(zhì)代謝和產(chǎn)熱功能受損,均與肥胖發(fā)生發(fā)展密切相關(guān)。最近有研究在肥胖和糖尿病患者白色脂肪細(xì)胞中觀察到過度自噬,機(jī)體高血糖狀態(tài)會引起脂肪組織自噬增加[24];自噬激活后會吞噬線粒體,米色或棕色脂肪細(xì)胞則轉(zhuǎn)化為白色脂肪細(xì)胞[11]。通過脂肪組織特異性敲除Atg5或Atg7后,可有效改善小鼠肥胖表型[23,25],這說明機(jī)體可以通過調(diào)節(jié)自噬活性以適應(yīng)不同代謝狀況,來確保脂肪組織代謝和功能平衡,從而預(yù)防糖尿病和肥胖癥等代謝性疾病的發(fā)生。通過白色脂肪細(xì)胞棕色化增加能量消耗治療肥胖及其并發(fā)癥成為新的研究熱點[26-27]。Prdm16作為米色脂肪細(xì)胞重要的轉(zhuǎn)錄因子,可激活棕色脂肪特異性基因,是維持棕色脂肪組織活性和白色脂肪組織中米色脂肪細(xì)胞生成所必需的[28];Ucp1在產(chǎn)生ATP的氧化代謝解偶聯(lián)中發(fā)揮關(guān)鍵作用[29],Cidea則可以促進(jìn)脂肪細(xì)胞脂解和產(chǎn)熱,這些都是米色脂肪細(xì)胞產(chǎn)熱的標(biāo)志性基因。本研究中ATG101在米色脂肪細(xì)胞分化過程中的表達(dá)增加,通過shRNA體外敲減Atg101后,Ucp1、Prdm16和Cidea水平顯著下調(diào),提示米色脂肪細(xì)胞的產(chǎn)熱過程受到抑制。表明ATG101的表達(dá)變化與米色脂肪細(xì)胞的產(chǎn)熱過程密切相關(guān)。
總之,本研究結(jié)果表明ATG101在米色脂肪成脂分化和產(chǎn)熱過程中可能發(fā)揮重要作用,這為剖析自噬在脂肪重塑過程中的作用提供了新證據(jù)。但未來還需要進(jìn)行在體實驗以深入探討ATG101對于維持機(jī)體能量穩(wěn)態(tài)和參與肥胖發(fā)生等病理過程的調(diào)節(jié)作用。ATG101可能參與脂肪組織的重塑并調(diào)節(jié)機(jī)體代謝,解析其表達(dá)調(diào)控網(wǎng)絡(luò)有望為研發(fā)肥胖癥防治藥靶開啟新路徑。
