尼羅羅非魚(yú)Galectin-4基因的原核表達(dá)及誘導(dǎo)條件優(yōu)化
2020-07-24 08:23:36張志強(qiáng)劉鑫潮牛金中簡(jiǎn)紀(jì)常
廣東海洋大學(xué)學(xué)報(bào) 2020年5期
關(guān)鍵詞:優(yōu)化
張志強(qiáng),劉鑫潮,牛金中,黃 瑜,王 蓓,簡(jiǎn)紀(jì)常
尼羅羅非魚(yú)基因的原核表達(dá)及誘導(dǎo)條件優(yōu)化
張志強(qiáng),劉鑫潮,牛金中,黃 瑜,王 蓓,簡(jiǎn)紀(jì)常
(廣東海洋大學(xué)水產(chǎn)學(xué)院 // 廣東省水產(chǎn)經(jīng)濟(jì)動(dòng)物病原生物學(xué)及流行病學(xué)重點(diǎn)實(shí)驗(yàn)室 // 廣東省水產(chǎn)經(jīng)濟(jì)動(dòng)物病害控制重點(diǎn)實(shí)驗(yàn)室,廣東 湛江 524088)
【】研究尼羅羅非魚(yú)()基因(記作)的原核表達(dá),為大量獲取尼羅羅非魚(yú)Galectin-4蛋白(OnGal-4)和后續(xù)功能研究奠定基礎(chǔ)。根據(jù)NCBI上公布的尼羅羅非魚(yú)基因序列(Genbank:XM-019345120)設(shè)計(jì)引物,構(gòu)建原核表達(dá)載體,誘導(dǎo)Galectin-4重組蛋白(rOnGal-4)在BL21原核表達(dá)系統(tǒng)中表達(dá),并優(yōu)化其誘導(dǎo)條件。成功擴(kuò)增出基因,并構(gòu)建原核表達(dá)載體pGEX-4T-OnGal-4。rOnGal-4最佳誘導(dǎo)條件是溫度28 ℃,IPTG濃度0.4 mmol/L,誘導(dǎo)時(shí)間4 h。在此條件下,rOnGal-4在可溶性蛋白中大量表達(dá)。Western-blot結(jié)果顯示,rOnGal-4可與抗GST標(biāo)簽的單克隆抗體發(fā)生特異性反應(yīng)。
尼羅羅非魚(yú);Galectin-4;基因克隆;原核表達(dá);條件優(yōu)化
尼羅羅非魚(yú)()是一種重要的世界性經(jīng)濟(jì)魚(yú)類,在我國(guó)南方廣泛養(yǎng)殖[1-2]。無(wú)乳鏈球菌()為羅非魚(yú)病原體之一,常會(huì)引起羅非魚(yú)患病死亡,近年來(lái)給我國(guó)和世界羅非魚(yú)養(yǎng)殖造成巨大的經(jīng)濟(jì)損失[3-4]。為有效控制疾病的發(fā)生與傳播,了解與深入研究羅非魚(yú)自身免疫系統(tǒng)尤為重要。
半乳糖凝集素(Galectin)是一類能與β-半乳糖苷特異結(jié)合的蛋白質(zhì),有調(diào)節(jié)細(xì)胞-細(xì)胞相互作用、細(xì)胞-基質(zhì)黏附和跨膜信號(hào)轉(zhuǎn)導(dǎo)等多種生理功能[5],可識(shí)別微生物的非自身多糖,被認(rèn)為是模式識(shí)別受體(Pattern Recognition Receptor),可調(diào)節(jié)病原體相關(guān)分子模式(Pathogen-AssociatedMolecular Pattern) 激活的天然免疫過(guò)程[6]。迄今已鑒定出15種哺乳動(dòng)物Galectin[7],其中Galectin-4有促進(jìn)細(xì)胞增殖、抑制細(xì)胞凋亡以及通過(guò)穩(wěn)定黏附連接復(fù)合物而促進(jìn)細(xì)胞黏附等功能[8],還可通過(guò)單核細(xì)胞的激活和分化發(fā)揮免疫調(diào)節(jié)功能[9],在病原體識(shí)別、T細(xì)胞凋亡和NK細(xì)胞活化中發(fā)揮重要作用[10-11]。Galectin-4基因已在人類和小鼠中有相關(guān)研究[12-13],但有關(guān)硬骨魚(yú)類中Galectin-4基因的報(bào)道僅見(jiàn)于線鱧()和大菱鲆()[14-15],Galectin-4在大菱鲆腸道中的表達(dá)水平最高,而在皮膚中的表達(dá)水平最低,另外,用鰻弧菌()和海豚鏈球菌()攻毒后,其表達(dá)量在腸道中顯著下調(diào)[15]。表明Galectin-4在魚(yú)類腸道抵抗感染的免疫反應(yīng)中起著至關(guān)重要的作用,但目前對(duì)于Galectin-4在尼羅羅非魚(yú)中的研究還未見(jiàn)報(bào)道。本研究通過(guò)克隆獲得尼羅羅非魚(yú)Galectin-4基因()的開(kāi)放閱讀框(ORF)全長(zhǎng),構(gòu)建原核表達(dá)載體pGEX-4T-OnGal-4,同時(shí)對(duì)誘導(dǎo)條件進(jìn)行了優(yōu)化,為大量獲取尼羅羅非魚(yú)Galectin-4重組蛋白(rOnGal-4)和研究該蛋白在尼羅羅非魚(yú)免疫系統(tǒng)中的功能奠定基礎(chǔ)。
1 材料與方法
1.1 材料
羅非魚(yú)購(gòu)自廣東省湛江市東風(fēng)水產(chǎn)品市場(chǎng)。pMD18-T載體、rTaq酶、DNA Maker、T4連接酶、R I和I,TaKaRa公司。總RNA提取試劑盒、cDNA反轉(zhuǎn)錄試劑盒,TRANS公司。彩虹Maker和GST標(biāo)簽蛋白純化試劑盒,碧云天公司。質(zhì)粒提取試劑盒和凝膠回收試劑盒,Thermo Fisher公司。大腸桿菌DH5a和BL21感受態(tài)細(xì)胞采用超級(jí)感受態(tài)試劑盒制作。原核表達(dá)載體pGEX-4T(+)Vector由本實(shí)驗(yàn)室保存。引物合成與菌液測(cè)序由生工生物股份有限公司完成。
1.2 方法
1.2.1基因的擴(kuò)增 剖取尼羅羅非魚(yú)的頭腎,在無(wú)RNA酶的環(huán)境中提取尼羅羅非魚(yú)的總RNA。將檢測(cè)合格(瓊脂糖凝膠電泳檢測(cè)結(jié)果為條帶單一) 的尼羅羅非魚(yú)總RNA用反轉(zhuǎn)錄試劑盒按說(shuō)明書(shū)將其反轉(zhuǎn)錄成cDNA模板。根據(jù)NCBI上預(yù)測(cè)的尼羅羅非魚(yú)基因的ORF全長(zhǎng)序列設(shè)計(jì)引物,引物序列為Gal4-R:CTTTG TTGCTCCTCCTGGC(R I)、Gal4-F:TAAGAA TAGAGAAGTGGATGTAGGAGAT(I)。以尼羅羅非魚(yú)頭腎的cDNA為模板,采用20 μL體系(Extaq Mix 10 μL、滅菌水7.5 μL、Gal4-R 1 μL、Gal4-F 1 μL、cDNA 0.5 μL)進(jìn)行PCR反應(yīng)擴(kuò)增,擴(kuò)增條件:95 ℃ 5 min、95 ℃ 30 s、57 ℃ 30 s、72 ℃ 70 s,循環(huán)36次;72 ℃下終延伸10 min。擴(kuò)增得到目的條帶后,用pMD18-T載體與純化回收產(chǎn)物在16 ℃連接30 min。取50 μL大腸桿菌DH5a感受態(tài)細(xì)胞,加入4 μL連接產(chǎn)物,混勻,冰浴20 min,42 ℃水浴鍋中熱激90 s,即刻置冰上冰浴2 min;加400 μL LB液體培養(yǎng)基,于37 ℃搖床振蕩培養(yǎng)45 ~ 60 min;取50 ~100 μL涂在LA(帶有Amp+的LB)瓊脂培養(yǎng)板上,于37 ℃下倒置培養(yǎng)12 h。挑取單菌落用通用引物M13、RV進(jìn)行菌落PCR鑒定,陽(yáng)性菌落送去公司測(cè)序。
1.2.2基因原核表達(dá)載體的構(gòu)建 對(duì)測(cè)序正確的菌液和pGEX-4T(+)空載菌種提取質(zhì)粒,將質(zhì)粒在37 ℃下雙酶切30 min,純化回收酶切后的目的片段。用T4連接酶將回收的目的片段與載體片段于4 ℃下連接10 h。將連接產(chǎn)物轉(zhuǎn)入大腸桿菌DH5a感受態(tài)中,方法同1.2.1。采用pGEX4T(F)、pGEX4T(R) 進(jìn)行菌落PCR鑒定,選取陽(yáng)性克隆菌測(cè)序。對(duì)測(cè)序吻合的菌液提取質(zhì)粒,并將提取的質(zhì)粒轉(zhuǎn)入大腸桿菌BL21感受態(tài),用LA液體培養(yǎng)基在37 ℃搖床培育3 ~ 5 h。
1.2.3 rOnGal-4的誘導(dǎo)表達(dá) 待菌液的OD600值達(dá)到0.5時(shí),取部分菌液在超凈臺(tái)用體積分?jǐn)?shù)50%甘油保種,置于? 80 ℃冰箱保存,剩余部分用錐形瓶分裝為兩瓶。在其中一瓶中加入異丙基---硫代半乳糖苷(IPTG)至終濃度1 mmol/L,另一瓶不加IPTG,兩瓶菌液均置于18 ℃、120 r/min的搖床誘導(dǎo)9 h。收集誘導(dǎo)菌液和未誘導(dǎo)菌液各2 mL,以12 000 r/min離心,棄上清液,沉淀用磷酸鹽緩沖液(PBS)重懸3次,加入PBS重懸菌液50 μL,蛋白上樣緩沖液10 μL,混勻。于100 ℃水中煮沸5 min,離心,取上清液(全菌蛋白)進(jìn)行SDS-PAGE電泳檢測(cè)。
1.2.4 rOnGal-4表達(dá)條件優(yōu)化 在蛋白可成功表達(dá)的前提下,采取單一變量控制法對(duì)蛋白誘導(dǎo)表達(dá)條件進(jìn)行逐一優(yōu)化。比較rOnGal-4在IPTG濃度分別為0、0.2、0.4、0.6、0.8、1.0、1.2 mmol/L,誘導(dǎo)時(shí)間分別為0、1、2、3、4、5、6、7 h,誘導(dǎo)溫度分別為18、28、37℃下的表達(dá)情況,篩選出最佳誘導(dǎo)條件。
1.2.5 rOnGal-4的純化及Western blot鑒定 最佳誘導(dǎo)條件下誘導(dǎo)rOnGal-4,然后收集誘導(dǎo)的菌液,進(jìn)行超聲破碎,在4℃、8 000 r/min條件下離心40 min,取上清液通過(guò)GST鎳柱層析法純化蛋白。取所得蛋白兩份進(jìn)行SDS-PAGE電泳,一份染色鑒定,同時(shí)將另一份用半干法轉(zhuǎn)膜(5.5 mA 10 min)到PVDF膜上,于4 ℃下用含有50 mg/mL脫脂奶粉的TBST封閉10 h。先用鼠抗GST多克隆抗體(1∶5 000稀釋)室溫條件下孵育2 h,再用HRP標(biāo)記的羊抗鼠IgM(1∶5000稀釋)以室溫條件孵育1 h,用超敏快速Western blot檢測(cè)試劑盒顯色檢測(cè),拍照。
2 結(jié)果
2.1 OnGal-4基因的PCR擴(kuò)增和鑒定
PCR擴(kuò)增獲得一1 194 bp的條帶(圖1),條帶大小與目的片段的ORF全長(zhǎng)相符。將擴(kuò)增片段和pMD-18T載體相連,轉(zhuǎn)化,涂板,挑菌,用通用引物M13和RV鑒定單菌落,得到一1 354 bp的條帶(圖1),表明該菌落為陽(yáng)性克隆。將陽(yáng)性克隆菌落送去測(cè)序,測(cè)序結(jié)果顯示插入pMD-18T載體的片段與目的片段完全一致。
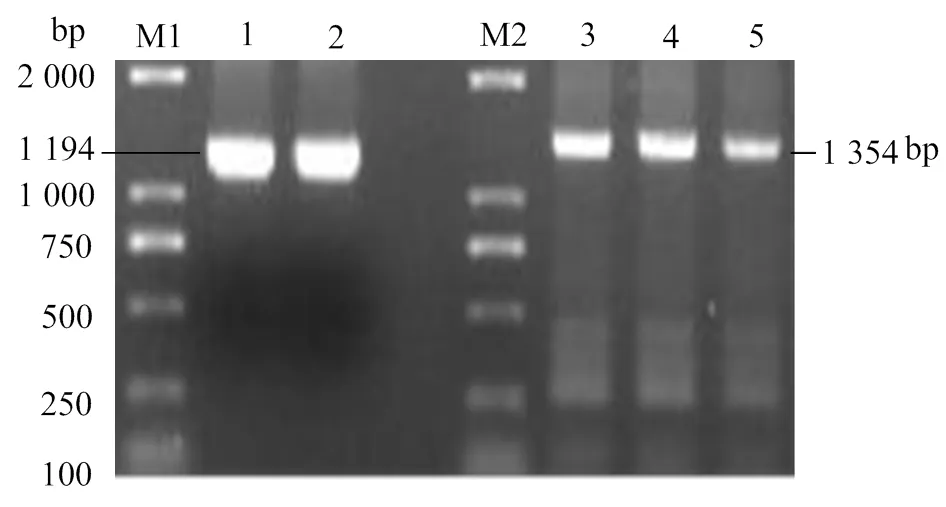
M1、M2,DL2000 DNA分子標(biāo)準(zhǔn);1、2,引物Gal4-R、Gal4-F擴(kuò)增產(chǎn)物;3、4、5,引物M13和RV擴(kuò)增產(chǎn)物
M1 and M2, DNA molecular Marker DL2000; 1 and 2, PCR amplification products by using Gal4-R and Gal4-F primers; 3, 4 and 5, PCR amplification products by using M13 and RV primers
圖1基因PCR擴(kuò)增和鑒定
Fig. 1 Cloning and identification ofgene
2.2 重組質(zhì)粒pGEX-4T-OnGal-4的酶切鑒定
重組質(zhì)粒pGEX-4T-OnGal-4雙酶切后,在1 194 bp處得一段酶切產(chǎn)物,該酶切產(chǎn)物與基因的分子大小相同,并在4 969 bp處發(fā)現(xiàn)與pGEX-4T空載大小一致的條帶(圖2)。對(duì)質(zhì)粒測(cè)序后,結(jié)果顯示成功插入該載體。
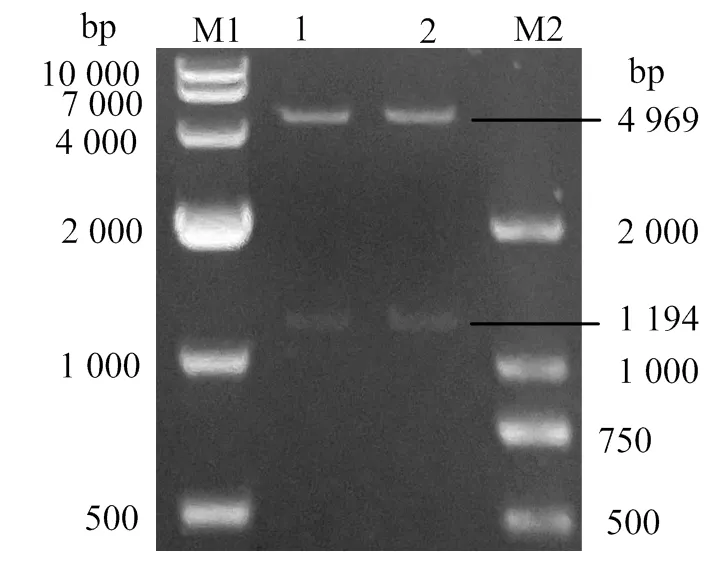
M1、M2,DL10000 DNA、DL2000 DNA 分子標(biāo)準(zhǔn);1、2,重組質(zhì)粒雙酶切的產(chǎn)物
2.3 rOnGal-4原核表達(dá)鑒定
rOnGal-4的原核表達(dá)鑒定結(jié)果(圖3)顯示,rOnGal-4誘導(dǎo)后在68 ku處出現(xiàn)一明顯條帶,而未誘導(dǎo)的rOnGal-4在68 ku處并未出現(xiàn)明顯條帶,表明該蛋白誘導(dǎo)表達(dá)獲得成功。同時(shí)誘導(dǎo)pGEX-4T空載發(fā)現(xiàn)GST標(biāo)簽蛋白大小為26 ku,因此,誘導(dǎo)的rOnGal-4大小為42 ku,與預(yù)測(cè)的目標(biāo)蛋白大小相同。
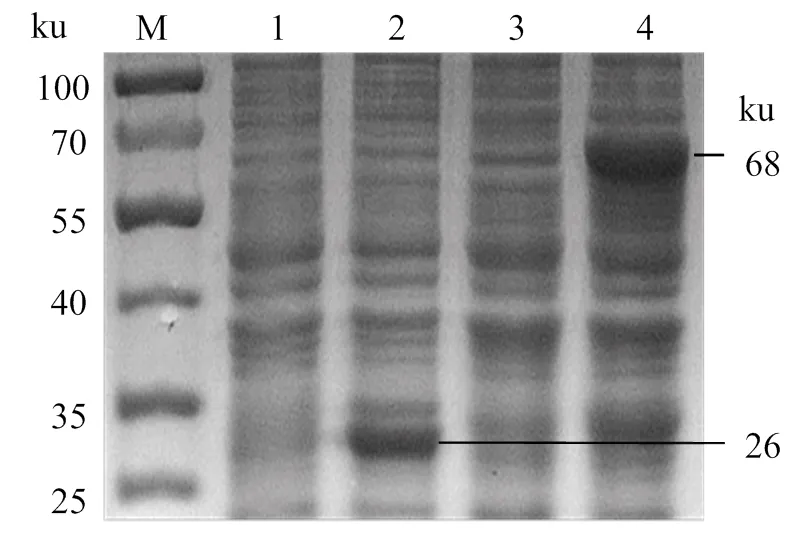
M,蛋白分子標(biāo)準(zhǔn);1、2,未誘導(dǎo)和誘導(dǎo)后的pGEX-4T;3、4,未誘導(dǎo)和誘導(dǎo)后的rOnGal-4
2.4 rOnGal-4表達(dá)條件優(yōu)化
2.4.1 rOnGal-4誘導(dǎo)物濃度優(yōu)化 誘導(dǎo)物IPTG濃度優(yōu)化結(jié)果(圖4)表明,rOnGal-4的表達(dá)量隨IPTG濃度的升高呈現(xiàn)先增加再保持平穩(wěn)最后下降的趨勢(shì),當(dāng)IPTG濃度達(dá)到0.4 mmol/L時(shí),rOnGal-4表達(dá)量達(dá)到最大。再增加IPTG濃度rOnGal-4表達(dá)量不會(huì)明顯增加;但當(dāng)其濃度達(dá)到1.2 mmol/L時(shí),rOnGal-4表達(dá)量反而會(huì)減少。因此,確定其誘導(dǎo)物IPTG的最佳濃度是0.4 mmol/L。
M,蛋白分子標(biāo)準(zhǔn);1,未誘導(dǎo)的rOnGal-4;2-7,IPTG誘導(dǎo)濃度分別為0.2、0.4、0.6、0.8、1.0、1.2 mmol/L
2.4.2 rOnGal-4誘導(dǎo)時(shí)間優(yōu)化 誘導(dǎo)時(shí)間優(yōu)化結(jié)果顯示在IPTG濃度和誘導(dǎo)溫度一定時(shí),rOnGal-4表達(dá)量在一定范圍內(nèi)會(huì)隨著誘導(dǎo)時(shí)間的延長(zhǎng)而逐漸增加,但當(dāng)誘導(dǎo)時(shí)間達(dá)到4 h后再延長(zhǎng)時(shí)間其表達(dá)量無(wú)明顯增加(圖5),由此確定rOnGal-4的最佳誘導(dǎo)時(shí)間是4 h。
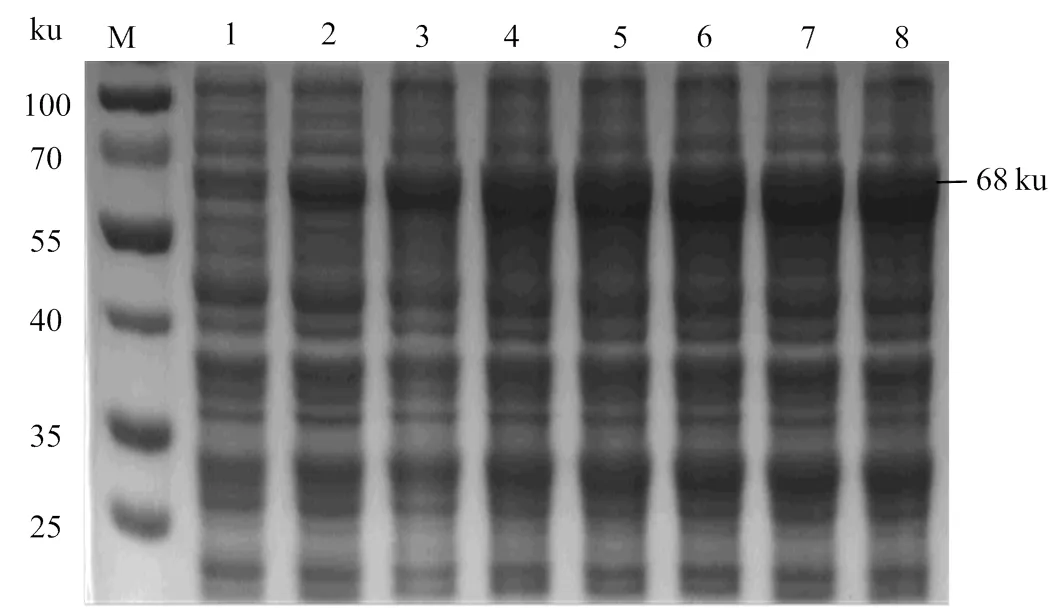
M,蛋白分子標(biāo)準(zhǔn);1-8,誘導(dǎo)時(shí)間濃度分別為0、1、2、3、4、5、6、7 h
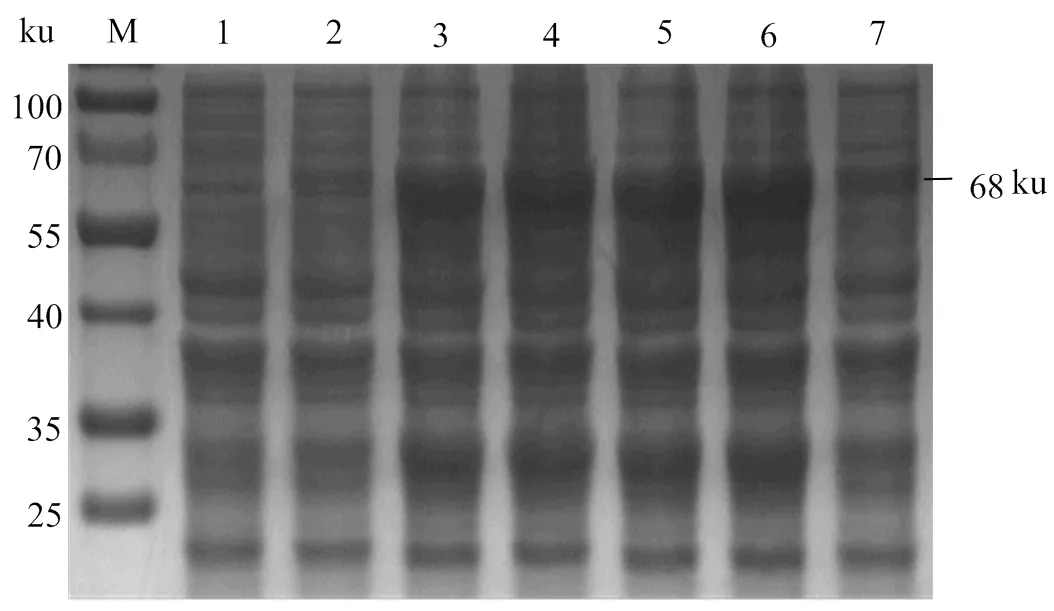
2.4.3 rOnGal-4誘導(dǎo)溫度優(yōu)化 圖6表明,在IPTG濃度和誘導(dǎo)時(shí)間一定時(shí),不同溫度條件對(duì)rOnGal-4在可溶性蛋白及包涵體蛋白中的表達(dá)量均有一定影響。在28℃誘導(dǎo)時(shí)rOnGal-4的可溶性蛋白和包涵體蛋白表達(dá)量最高,在37 ℃誘導(dǎo)時(shí)其可溶性蛋白的表達(dá)量最低。同時(shí)對(duì)rOnGal-4可溶性表達(dá)分析發(fā)現(xiàn),其在包涵體蛋白中的表達(dá)量較高,但是由于可溶性蛋白較易獲得且不會(huì)對(duì)蛋白活性有較多的損傷,更有利于后續(xù)試驗(yàn)的開(kāi)展,所以確定rOnGal-4最佳誘導(dǎo)溫度是28 ℃。
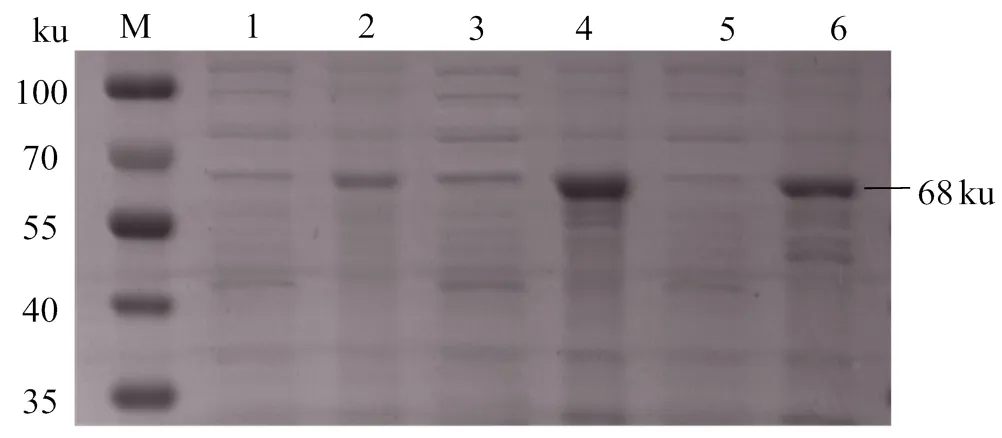
M,蛋白分子標(biāo)準(zhǔn);1、3、5,18、28、37 ℃誘導(dǎo)表達(dá)的可溶性蛋白;2、4、6,18、28、37 ℃誘導(dǎo)表達(dá)的包涵體蛋白
2.5 rOnGal-4純化鑒定和western bolt鑒定
在上述優(yōu)化得到的最佳誘導(dǎo)條件下誘導(dǎo)rOnGal-4。誘導(dǎo)過(guò)的rOnGal-4全菌蛋白、可溶性蛋白和純化后rOnGal-4的SDS-PAGE檢測(cè)結(jié)果(圖7A)顯示,在68 ku處均有條帶且純化后的rOnGal-4呈現(xiàn)單一條帶,表明蛋白純化成功。對(duì)純化后的蛋白進(jìn)行Western bolt驗(yàn)證,結(jié)果顯示為單一條帶(圖7B)。
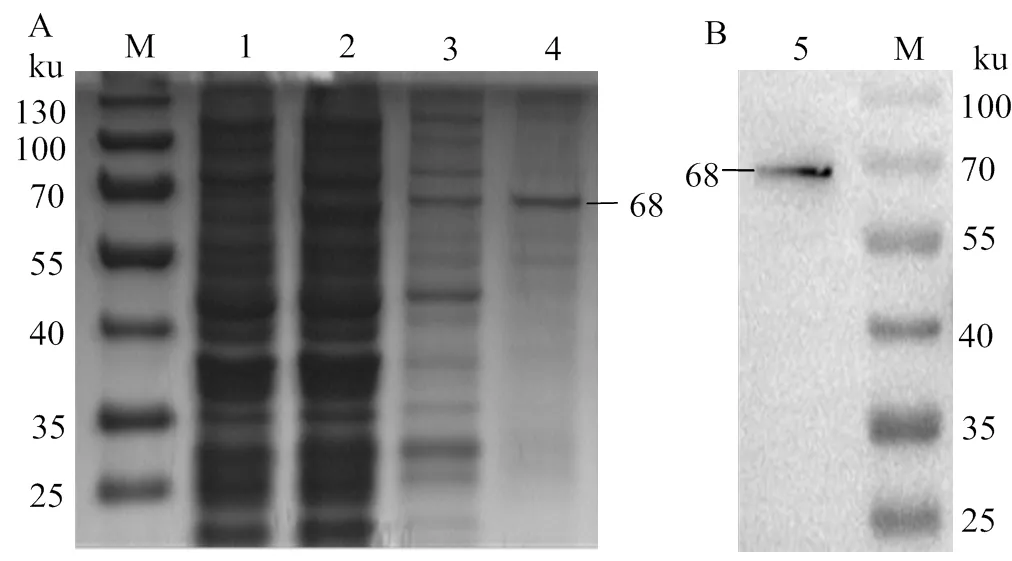
A, SDS-PAGE鑒定; B, Western bolt鑒定; M, 蛋白分子標(biāo)準(zhǔn);1, 未誘導(dǎo)的rOnGal-4;2、3, 誘導(dǎo)后rOnGal-4的全菌蛋白、可溶性蛋白;4、5, 純化后的rOnGal-4
A, identification by SDS-PAGE; B, identification by Western bolt;
M, Protein molecular Marker; 1, rOnGal-4 without inducing; 2 and 3, The whole and Soluble protein of rOnGal-4 after induction; 4 and 5, Purified rOnGal-4 protein
圖7 純化后rOnGal-4的鑒定
Fig. 7 Identification for the purified rOnGal-4
3 討論
原核系統(tǒng)具有在短時(shí)間內(nèi)獲得大量的重組蛋白、簡(jiǎn)單而廉價(jià)的細(xì)菌細(xì)胞培養(yǎng)以及眾所周知的轉(zhuǎn)錄和翻譯機(jī)制,基因修飾的簡(jiǎn)便性和許多細(xì)菌突變體的可用性等優(yōu)點(diǎn),因而在研究中被廣泛應(yīng)用[17]。為獲得大量的重組蛋白,本研究選用大腸桿菌原核表達(dá)系統(tǒng),并選用目前調(diào)節(jié)T7啟動(dòng)子轉(zhuǎn)錄活性最有效的分子誘導(dǎo)劑IPTG作為誘導(dǎo)劑[18]。因原核表達(dá)效果會(huì)受諸多表達(dá)元件和外界條件的影響[19],故對(duì)其誘導(dǎo)劑濃度、誘導(dǎo)時(shí)間和誘導(dǎo)溫度進(jìn)行了優(yōu)化。結(jié)果表明,在IPTG濃度為0.4 mmol/L時(shí)rOnGal-4表達(dá)量最大。羅氏沼蝦雌激素相關(guān)受體() 基因[20]、哈維氏弧菌基因[21]等的原核表達(dá)條件優(yōu)化結(jié)果顯示,IPTG濃度應(yīng)在0.1 ~ 0.8 mmol/L范圍,與本研究結(jié)果一致。IPTG濃度為1.2 mmol/L時(shí)蛋白表達(dá)量會(huì)下降,是IPTG毒性導(dǎo)致高濃度時(shí)抑制蛋白的表達(dá)[22]所致。誘導(dǎo)4 h rOnGal-4表達(dá)量最大,這與尼羅羅非魚(yú)補(bǔ)體3基因[23]和尼羅羅非魚(yú)Galectin-3基因[24]的表達(dá)條件優(yōu)化結(jié)果相似。在28 ℃誘導(dǎo)時(shí),rOnGal-4可溶性蛋白含量最高,與尼羅羅非魚(yú)NCCRP-1基因[25]和人β2微球蛋白[26]的結(jié)果一致。可能原因是在37 ℃時(shí)大腸桿菌處于最佳生長(zhǎng)狀態(tài),合成蛋白的速度快,導(dǎo)致蛋白來(lái)不及折疊形成包涵體。用最佳表達(dá)條件下誘導(dǎo),所得rOnGal-4可溶性蛋白可被成功表達(dá)和純化。本研究可為后續(xù)大量獲取該蛋白制作多克隆抗體和為后續(xù)該蛋白的功能研究奠定基礎(chǔ)。
[1] 甘楨. 尼羅羅非魚(yú)T細(xì)胞激活相關(guān)基因的功能研究[D].湛江:廣東海洋大學(xué), 2015.
[2] EL-SAYED A F M. Total replacement of fish meal with animal protein sources in Nile tilapia,(L.), feeds[J]. Aquaculture Research, 1998, 29(4): 275-280.
[3] LIU L, LU D Q, XU J, et al. Development of attenuated erythromycin-resistantvaccine for tilapia () culture[J]. Journal of Fish Diseases, 2019, 42(5): 693-701.
[4] WANG Y Y, WANG Q, ZENG W W, et al. Establishment and characterization of a cell line from tilapia brain for detection of tilapia lake virus[J]. Journal of Fish Diseases, 2018, 41(12): 1803-1809.
[5] LIU F T, RABINOVICH G A. Galectins: regulators of acute and chronic inflammation[J]. Annals of the New York Academy of Sciences, 2010, 1183(1): 158-182.
[6] MAHLA R S, REDDY M C, PRASAD D V R, et al. Sweeten PAMPs: role of sugar complexed PAMPs in innate immunity and vaccine biology[J]. Frontiers in Immunology, 2013, 4: 248.
[7] HUGHES R C. Secretion of the galectin family of mammalian carbohydrate-binding proteins[J]. Biochimica et Biophysica Acta (BBA) - General Subjects, 1999, 1473(1): 172-185.
[8] CHEN C C, WANG G X, HUANG X H, et al. Overexpression of galectin-4 promotes cell growth of hepatocellular carcinoma cellsand[J]. International Journal of Clinical and Experimental Pathology, 2017, 10(10): 10233-10242.
[9] HONG S H, SHIN J S, CHUNG H, et al. Galectin-4 interaction with CD14 triggers the differentiation of monocytes into macrophage-like cells via the MAPK signaling pathway[J]. Immune Network, 2019, 19(3): e17.
[10] STURM A, LENSCH M, ANDRé S, et al. Human galectin-2: novel inducer of T cell apoptosis with distinct profile of caspase activation[J]. Journal of Immunology, 2004, 173(6): 3825-3837.
[11] BRITTOLI A, FALLARINI S, ZHANG H, et al. “In vitro” studies on galectin-3 in human natural killer cells[J]. Immunology Letters, 2018, 194: 4-12.
[12] MICHALAK M, WARNKEN U, SCHN?LZER M, et al. Detection of malignancy-associated phosphoproteome changes in human colorectal cancer induced by cell surface binding of growth-inhibitory galectin-4[J]. IUBMB Life, 2019, 71(3): 364-375.
[13] NIEPCERON E, SIMIAN F, LOUISOT P, et al. Expression of galectin 4 in the rat small intestine during postnatal development[J]. Biochimie, 2004, 86(2): 115-118.
[14] ARASU A, KUMARESAN V, GANESH M R, et al. Bactericidal activity of fish galectin 4 derived membrane-binding peptide tagged with oligotryptophan[J]. Developmental & Comparative Immunology, 2017, 71: 37-48.
[15] CHEN J H, ZHANG L, YANG N, et al. Expression profiling and microbial ligand binding analysis of galectin-4 in turbot (L.)[J]. Fish & Shellfish Immunology, 2019, 84: 673-679.
[16] WOOTERS M A, HILDRETH M B, NELSON E A, et al. Immunohistochemical characterization of the distribution of galectin-4 in porcine small intestine[J]. Journal of Histochemistry & Cytochemistry, 2005, 53(2): 197-205.
[17] POROWI?SKA D, WUJAK M, ROSZEK K, et al. Prokaryotic expression systems[J]. Post?py Higieny i Medycyny Do?wiadczalnej (Online), 2013, 67: 119-129.
[18] BRIAND L, MARCION G, KRIZNIK A, et al. A self-inducible heterologous protein expression system in[J]. Scientific Reports, 2016, 6: 33037.
[19] GILEADI O. Recombinant protein expression in E. coli: A historical perspective[M]//Methods in Molecular Biology. New York, NY: Springer New York, 2017: 3-10. DOI: 10.1007/978-1-4939-6 887-9_1.
[20] 趙苗鑫, 陳華譜, 劉金磊, 等. 羅氏沼蝦雌激素相關(guān)受體() 基因原核表達(dá)與純化[J]. 廣東海洋大學(xué)學(xué)報(bào), 2017, 37(1): 108-112.
[21] 馬少鴻, 黃郁蔥, 簡(jiǎn)紀(jì)常, 等. 哈維氏弧菌基因的克隆及原核表達(dá)分析[J]. 廣東海洋大學(xué)學(xué)報(bào), 2019, 39(5): 1-7.
[22] DVORAK P, CHRAST L, NIKEL P I, et al. Exacerbation of substrate toxicity by IPTG inBL21(DE3) carrying a synthetic metabolic pathway[J]. Microbial Cell Factories, 2015, 14(1): 1-15.
[23] 牛金中, 黃瑜, 湯菊芬, 等. 尼羅羅非魚(yú)補(bǔ)體3基因片段的原核表達(dá)及條件優(yōu)化[J]. 廣東海洋大學(xué)學(xué)報(bào), 2018, 38(2): 80-84.
[24] 羅國(guó)玲, 牛金中, 黃瑜, 等. 尼羅羅非魚(yú)Galectin-3基因的原核表達(dá)與條件優(yōu)化[J]. 廣東海洋大學(xué)學(xué)報(bào), 2019, 39(4): 35-41.
[25] 黃瑜, 牛金中, 湯菊芬, 等. 尼羅羅非魚(yú)基因的原核表達(dá)及條件優(yōu)化[J]. 基因組學(xué)與應(yīng)用生物學(xué), 2018, 37(9): 3753-3758.
[26] 矯麗媛, 才蕾, 任艷娜, 等. 人β2微球蛋白原核表達(dá)條件優(yōu)化及純化[J]. 生物醫(yī)學(xué)工程學(xué)雜志, 2015, 32(5): 1050-1055.
[27] 馬德昭, 田菲, 吳其中, 等. 青海湖裸鯉IL-8的原核表達(dá)及誘導(dǎo)條件優(yōu)化[J/OL]. 基因組學(xué)與應(yīng)用生物學(xué): 1-7. [2020-04-08]. https://kns.cnki.net/KCMS/detail/45. 1369.Q.20190508.1427.005.html?uid
Prokaryotic Expression and Optimization ofGene from Nile Tilapia ()
ZHANG Zhi-qiang, LIU xin-chao, NIU Jin-zhong, HUANG Yu, WANG Bei, JIAN Ji-chang
(////,524088,)
【】To study the optimization conditions for prokaryotic expression of the Nile tilapia ()gene , and perform functional studiesof Galectin-4.【】A pair of primers were designed based on the tilapiagene sequence (Genbank: XM-019345120), and to expression plasmid (pGEX-4T-OnGal-4) was produced. The Galectin-4 recombinant clone was induced to express rOnCal4 in BL21 cells and the expression conditions were optimized .【】Thegene was successfully amplified and the prokaryotic expression plasmid pGEX-4T-OnGal-4 was constructed. The optimal induction conditions for rOnGal-4 production were 0.4 mmol/L of IPTG at 28 ℃ for 4 h. The rOnGal-4 protein was highly expressed under the optimized expression conditions. Western-blot results showed that the rOnGal-4 could specifically react with GST-tag monoclonal antibody.
;; gene clone; prokaryotic expression; condition optimization
Q78;Q959.223+.63
A
1673-9159(2020)05-0118-06
10.3969/j.issn.1673-9159.2020.05.015
2020-03-05
國(guó)家自然科學(xué)基金青年基金項(xiàng)目(31702386);廣東省科技廳國(guó)際科技合作領(lǐng)域項(xiàng)目(2017A050501037);廣東省普通高校青年創(chuàng)新人才類項(xiàng)目(2018KQNCX105)
張志強(qiáng)(1997-),男,碩士研究生,研究方向?yàn)樗镝t(yī)學(xué)。E-mail:248798423@qq.com
簡(jiǎn)紀(jì)常(1964-),博士,教授,研究方向?yàn)樗a(chǎn)經(jīng)濟(jì)動(dòng)物病害防治。E-mail:jianjc@gdou.edu.cn
黃瑜(1986-),博士,講師,研究方向?yàn)轸~(yú)類免疫學(xué)。E-mail: huangyu@gdou.edu.cn
張志強(qiáng),劉鑫潮,牛金中,等. 尼羅羅非魚(yú)Galectin-4基因的原核表達(dá)及誘導(dǎo)條件優(yōu)化[J]. 廣東海洋大學(xué)學(xué)報(bào),2020,40(5):118-123.
(責(zé)任編輯:劉慶穎)
猜你喜歡
房地產(chǎn)導(dǎo)刊(2022年5期)2022-06-01 06:20:14
能源工程(2022年1期)2022-03-29 01:06:28
建材發(fā)展導(dǎo)向(2021年12期)2021-07-22 08:06:48
建材發(fā)展導(dǎo)向(2021年7期)2021-07-16 07:07:52
中學(xué)生數(shù)理化(高中版.高二數(shù)學(xué))(2021年12期)2021-04-26 07:43:48
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2021年12期)2021-03-08 01:28:50
今日農(nóng)業(yè)(2020年16期)2020-12-14 15:04:59
消費(fèi)導(dǎo)刊(2018年8期)2018-05-25 13:20:08
家庭影院技術(shù)(2018年4期)2018-05-09 07:07:41
電子制作(2017年20期)2017-04-26 06:57:45
