論植物雜種優勢的研究途徑
2020-07-27 16:30:13潘春桂
天津農業科學 2020年5期

摘? ? 要:植物群體葉面積指數與平均單位葉面積截獲光能的反相關導致群體光能截獲雜種優勢形成的必然性,各種養分環境消耗進入臨界點同步性改善導致養分獲取雜種優勢形成的必然性。植株質量或作物產量的雜種優勢主要由雙親群體光能截獲和各種養分獲取的特征及差異所決定。遺傳與環境的相互作用決定了雜種優勢的形成過程。動態跟蹤植株群體光照結構、植株質量或產量形成過程、根際環境中各種養分的濃度等因子有助于理解雜種優勢形成過程,單獨研究雙親遺傳差異不能獲得雜種優勢形成的一般機制。
關鍵詞:光能;養分;反相關;同步性;必然性;雜種優勢
Abstract:The inverse correlation between the leaf area index of plant population and the light energy intercepted per unit leaf area leads to the inevitability of the formation of heterosis of population light energy intercepted, and the? improvement of synchronicity of various nutrient environmental consumption into the critical point leads to the inevitability of the formation of heterosis of nutrient obtained. The heterosis of plant weight or crop yield is mainly determined by the characteristics and differences of light interception and various nutrients acquisition of parents. The interaction between heredity and environment determines the formation of heterosis. Dynamic tracking of light structure, plant weight or yield formation process, concentration of various nutrients in rhizosphere environment and other factors is helpful to understand the formation process of heterosis. The general mechanism of heterosis formation can not be obtained by studying the genetic difference between parents in isolation.
Key words: light energy; nutrient; inverse correlation; syncronicity heterosis; inevitability; heterosis
從達爾文時代就已經認識到雜種優勢現象,人類利用雜種優勢增加經濟效益以及研究雜種優勢的形成機理也有近一個世紀。對其機理的解釋,經典遺傳學提出了顯性、超顯性、上位性效應假說,分子遺傳學認為基因差異表達、表觀遺傳修飾包括DNA甲基化和組蛋白修飾參與雜種優勢的形成[1],以上假說也獲得了相關試驗的支持。另外,一些學者也提出了遺傳平衡、活性基因遺傳振動活成、基因網絡系統及自組織理論等假說[2],不難看出,以上假說共同性在于將雜種一代內在相關過程的特異性等價為雜種優勢的形成機理或產生原因,實際上它們仍然為雜種優勢的表現或結果,就像雜種一代營養體較大,生物或經濟產量較高一樣,不能作為雜種優勢的形成原因。可能這些研究潛在地認為親本兩套遺傳物質相互作用就產生了雜種優勢,排斥或弱化了環境因素在雜種優勢形成中的地位。相同遺傳類型在不同環境中具有生理、生化、形態過程的差異性,表明環境效應從遺傳表達到形態建立各個環節中均產生重要影響。假設各基因位點加性表達,在外部環境作用之下,這種遺傳過程是不是能夠轉變呢?從能量平衡而言,雜種一代的優勢應該與獲得更多的光能和養分有關。本研究從上述基本假設出發,論述雜種優勢形成機理可能的研究途徑。
1 光能截獲的雜種優勢
如果一個乘積性狀等于兩個單因子性狀的乘積,兩個單因子大小在雙親間反向變化,并假設它們均為加性遺傳,則乘積性狀具有正向中親優勢或雜種優勢;單因子差異越大,雜種優勢越大。類似地,如果兩個單因子在雙親間同向變化,則存在負向雜種優勢,兩個單因子差異越大,雜種優勢越小[3]。當至少有一個單因子雙親相等時,雜種優勢為零;任一單因子雙親均乘以相同系數時,不改變雜種優勢大小。由于加性遺傳僅是理論上成立的,以上均為理論雜種優勢。在實際情形中,如果以上條件近似滿足時,結論也是適用的。
植物群體光能利用率隨群體密度而變化,當群體密度較小或葉面積指數較小時,群體冠層光能反射率較低,而透光率較高,各層葉片均可獲得較好的光照。當群體葉面積指數過大時,群體冠層光能反射率較高,而透光率下降,上部葉片經常處在飽和點之上,下部葉片可處在光補償點之下,平均單位葉面積截獲光能下降。這是一個必然的過程,群體葉面積指數越大,葉片或葉層互相遮掩越嚴重。在密度相同的情況下,以單株葉面積代替群體葉面積,單株截獲光能等于單株葉面積與單位葉面積截獲光能的乘積,可見這個乘積性狀兩個構成單因子具有反相關傾向。由于單位葉面積截獲光能和單株葉面積、葉片大小、形狀等有關,如果認為單株葉面積和單位葉面積截獲光能均近似為加性遺傳,則從上文分析可知,單株截獲光能的雜種優勢是一個本能過程[3]。下文從一些引文數據的計算結果來分析光能截獲和產量形成雜種優勢之間關系。司洪華[4]的15個玉米雜交種試驗表明,當以單株葉面積與單位葉面積籽粒產量的乘積來模擬上述單株光能截取過程時,光能截獲理論雜種優勢與實際籽粒產量雜種優勢相關系數為0.523 3(P<0.05)。吳才君等[5]的6個蕓薹種蔬菜交種試驗表明,當以單株葉面積與單位葉面積株重的乘積來模擬光能截獲時,光能截獲理論雜種優勢與單株重實際雜種優勢相關系數為0.843 4(P<0.05),原文結果亦表明,利用CDNA-AFLP技術研究雙親與雜交種的基因差異表達,未發現任何一種表達類型比例與株重雜種優勢有顯著相關性。由于植株體單株分枝數與單株葉面積存在線性關系,當以單株分枝數與單個分枝的產量的乘積來模擬單株光合產物時,趙禹凱等[6]的18個桔梗雜交種試驗表時,光合產物理論雜種優勢與桔梗果實數實際雜種優勢相關系數為0.471 3(P<0.05)。余政軍等[7]棉花5×6完全雙列雜交試驗表明,這里以正反交平均產量表示組合產量,當以單株果枝數與單個果枝皮棉產量的乘積來模擬光合產物時,光合產物理論雜種優勢與實際皮棉產量雜種優勢相關系數為0.526 4(P<0.05);楊祥波[8]30個大豆雜交種試驗表明,當以單株分枝數與單個分枝的籽粒產量乘積來模擬光合作物時,光合產物理論雜種優勢與產量實際雜種優勢相關系數為0.860 2(P<0.001),其研究結果顯示,SSP標記的分子距離與單株粒質量雜種優勢的相關系數為-0.42(P<0.05)。李素萍[9]的試驗數據顯示,如果構建一個乘積性狀,向日葵單株粒質量=(單株葉片數×生育期)×[單株粒質量/(單株葉片數×生育期)],親本間以上兩個乘積因子相關系數為-0.672(P<0.05),單株粒質量理論雜種優勢與實際雜種優勢相關系數為0.443 6(P<0.05),以上結果并不總是出現的,因為沒有考慮到產量形成過程或葉片功能期長短等的遺傳差異,雖然如此,以上結果也足夠提示了光能截獲這個乘積性狀的深入研究對于理解產量雜種優勢機制的重要性。以上光合產物或產量這個乘積性狀2個構成單因子均直接與光能截獲有關,由于可有多種方法將光合產物或產量分解為2個單因子乘積,例如棉花皮棉產量可以為籽棉產量與衣分的乘積,也可以分解總成鈴數與單鈴質量的乘積;水稻產量可以為總穗數與單穗質量的乘積,也可以為總粒數與粒質量的乘積。然后按照單因子加性遺傳來計算乘積性狀理論雜種優勢,這與上文從光能截獲角度來研究雜種優勢是否有同等效果呢?這值得進一步研究。
2 養分獲取的雜種優勢
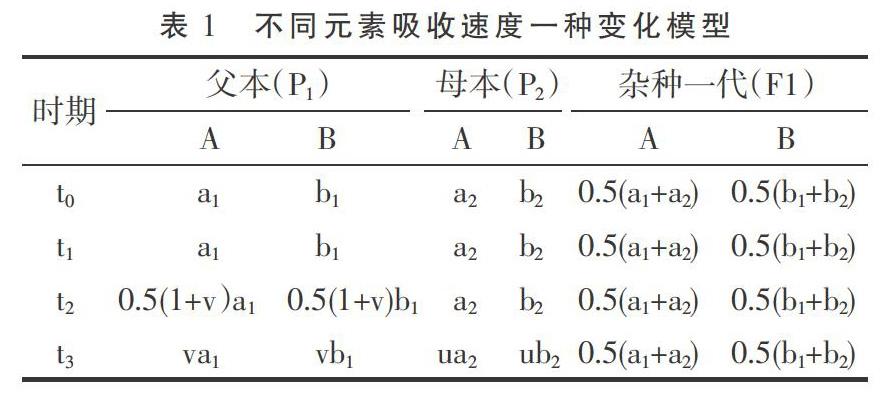
雜種優勢的形成也與吸收更多的營養元素有關。植株體對外部環境中養分吸收速度與遺傳、環境養分濃度及植株體大小等有關,可用Logistic曲線來模擬植株生長過程[10-11]。可近似認為,在一定條件之下,(1)植株體對各種營養元素的吸收存在飽和濃度或飽和期,此時增加任何一種養分的濃度或環境供給,植株體對各種元素的吸收速度不再增加[12],這里假定不考慮鹽害濃度;(2)在飽和期雜種一代對各種元素的吸收速度為加性遺傳;(3)植株體對各種元素吸收速度為固定比例,只與遺傳類型有關;(4)各種營養元素均存在臨界環境濃度或臨界點,對某種元素而言,此時減少其環境濃度,植株體對這種元素的吸收速度會下降,其他元素也協同下降,但吸收速度仍保持固定比例。基于以上假設,如果一些元素飽和期吸收速度在雙親間存在差異,則最大元素吸收速度被雙親所具有,因此,經過一段生長時間后,雙親均因為各自具有最大元素吸收速度可能比雜種一代更早地通過臨界點,其后雜種一代對各種元素吸收速度便超過中親值,因此,各器官或單株干質量的雜種優勢便開始形成。
根據上文假設條件,易得2F1/(p1+p2)>1,即此時段已形成正向中親優勢或雜種優勢。當然,對于某一種元素而言,不同生長時期飽和吸收速度存在差異,臨界環境濃度也不同,以上分析假設t0-t3時間較短或各種養分飽和吸收速度不變。
以上述養分吸收模型可產生如下推論。(1)在飽和期,理論上是不存在雜種優勢的;或者說,在植株生長很早時期和環境養分供給極端充分時,雜種優勢平均值較小或趨于零,變化也較小。可能這個時期很短,以致不易被分辨出來。(2)如果生長時間足夠長或各種養分已進入相對脅迫期,植株體養分吸收與環境供給相容性好易獲得較大的質量(鮮質量或干質量,下文同),相容性與質量呈正向關系。相容性好指各種養分有較大的飽和吸收速度、和環境供給有較好的協調性即各種養分進入臨界點時間相近以及各種養分有較大的(臨界)環境消耗率等。由于以相同的外部環境作共同參照,雙親質量越大,則遺傳相似性越好;雙親質量越小,由于存在各類不相容性,雙親總體上有較大的遺傳差異,但不同組合間可有一定變化。環境相容性好的雙親從通過臨界點到其后延續時間里,各種營養元素吸收速度的衰減(相對于飽和吸收速度衰減,下文同)和雜種一代相近,衰減期也和雜種一代較同步,因此不易獲得較大的雜種優勢;環境相容性差且遺傳差異較大時,可導致雙親間各養分平均衰減幅度更大,平均衰減時間更早,易形成較大的雜種優勢。因此在大多數情況下,雙親質量之和與雜種優勢易呈顯著的負相關。(3)植株體質量的變化與飽和養分吸收速度及其衰減均有關。如果前期一直處于飽和期,在臨界期附近;或植株生長很早時期和養分供給相對充足條件下,植株質量主要與各種養分尤其是大量元素飽和吸收速度有關,雙親質量之差可一定程度上反映遺傳差異;其較大時,雙親之一或雙親養分吸收速度在后續時間內有較大的衰減,衰減時間也較早;因此,此時刻雙親質量之差與鄰近較短時段內植株體質量增加量雜種優勢可呈正相關。(4)由于植株體質量取樣時間、試驗環境、遺傳背景的巧合,質量較小的親本或因飽和期養分吸收速度慢未通過或較遲通過臨界點,或因飽和期養分吸收速度慢通過臨界點后衰減也慢,也可能它們遺傳相似性好;質量較大的親本則相反。但通過臨界點之后延續時間均不長,飽和期養分吸收速度主要決定了植株體質量。這時雙親質量之和與雜種優勢可呈弱正相關或正相關。但不排除一些不同試驗條件下可產生相似的遺傳過程。下文通過文獻數據的計算結果來證明以上推論。馬謙[13]的試驗數據表明,大田條件下水稻播種后第20天的幼苗干質量雜種優勢為-5.8±19.4(%),而第30天為23.41±19.1(%),第20天到第30天幼苗干質量增加量雜種優勢與第20天雙親干質量之差絕對值呈正相關,相關系數為0.379 9(P<0.05),與第30天干質量雙親之和呈負相關,相關系數為-0.527 2(P<0.005)。另外,第20天幼苗干質量雜種優勢與同時刻雙親干質量之和相關系數為-0.044 5,第30天幼苗干質量的雜種優勢與此時雙親之和呈弱負相關,相關系數為-0.323 7(P<0.1)。以上結果表明,第20天時幾乎所有親本仍處于飽和期或在臨界點附近。第30天時,所有親本均已通過臨界期,但其后延續時間均較短或各種養分環境脅迫仍不嚴重。馬謙[13]還有一個黑暗條件之下水稻種子發芽4天后幼芽干質量數據,幼芽干質量的雜種優勢與雙親之和呈負相關,相關系數為-0.677 8(P<0.05),雜種一代幼芽干質量和中親值之差(離親值,下文同)與中親值相關系數為-0.603 4(P<0.1),由于發芽時種子貯藏的干物質是唯一能量來源,說明第4天時,各種營養元素已存在明顯的脅迫。余麗霞[14]棉花雜交種愈傷組織培養試驗數據計算表明,愈傷組織質量雜種優勢與同時刻雙親之和的相關系數在第35天、第65天、第80天分別為-0.597 4(P<0.002)、-0.675 0(P<0.001)、及-0.696 5(P<0.001);雜種一代愈傷組織重量離親值與同時刻中親值相關系數在第35天、第65天及第80天分別-0.601 0(P<0.002)、-0.670 0(P<0.001)、-0.700 6(P<0.001);第35天到第65天愈傷組織質量增加量雜種優勢與第35天、第65天、第80天雙親質量之和相關系數分別為-0.508 2(P<0.01)、-0.714 1(P<0.001)、-0.708 9(P<0.001),說明在第35天時,各種養分早已進入協迫階段。第65天到第80天愈傷組織質量增加量雜種優勢與第35天、第65天、第80天雙親質量之和相關系數分別為-0.544 2(P<0.005)、-0.533 1(P<0.01)、-0.590 3(P<0.002),可能生長后期外界養分已較嚴重虧缺,雜種一代對各種養分吸收速度也衰減嚴重,雜種優勢變得較不活躍,雙親遺傳差異大小與愈傷組織增加量雜種優勢相關性減弱。王章奎[15]營養液栽培小麥試驗表明,拔節期單株根系干質量雜種優勢與雙親之和相關系數為-0.750 5(P<0.001),雜種一代單株根系干重離親值與中親值相關系數為-0.704 9(P<0.001),說明這時營養液各種養分的供給已進入較重的脅迫階段。溫玲[16]甜瓜雜交種試驗表明,4葉一心期單株根系干質量雜種優勢與雙親之和相關系數為0.743 5(P<0.05),這與上文推論第(4)點有關。同時也表明取樣時各種養分的環境供給可能仍比較充裕。蔡麗艷等[17]苜蓿雜交種數據也可能屬于同樣性質,苜蓿干草單株質量的雜種優勢與雙親之和相關系數為0.388 3(P<0.05)。大田作物經過一個生長季節,土壤養分已處于相對虧缺狀態,這時作物產量雜種優勢與雙親之和可為顯著或極顯著負相關。林欣立等[18]蘿卜雜交試驗表明,單根質量雜種優勢與雙親之和相關系數為-0.583 8(P<0.05)。潘春桂[19]計算結果表明,皮棉產量雜種優勢與雙親之和相關系數為-0.696 6(P<0.001)。徐靜斐和汪路應[20]的水稻試驗表明,水稻產量的雜種優勢與雙親之和相關系數為-0.523 4(P<0.002)。司洪華[4]玉米雜交試驗表明,玉米產量雜種優勢與雙親之和相關系數為-0.840 3(P<0.001)。吳利民等[21]小麥雜種試驗,小麥產量雜種優勢與雙親之和相關系數為-0.483 3(P<0.05)。梁耀平等[22]西瓜雜交試驗表明,西瓜產量雜種優勢與雙親之和相關系數為-0.715 7(P<0.001)。閻冬生[23]大豆雜交試驗表明,大豆產量雜種優勢與雙親之和相關系數為-0.848 5(P<0.001)。桑世飛等[24]的油菜雜交試驗表明,油菜產量雜種優勢與產量雙親之和相關系數為 -0.523 4(P<0.001)。由于雜種優勢與雙親之和相關性受到自相關的影響,進一步計算出雜種一代產量離親值與中親值的相關性,絕大多數仍呈顯著或極顯著負相關,只有林欣立、司洪華的試驗中兩者呈弱負相關,這里未列出相關系數。另外,上文已經提及,雙親之和越小時,雙親遺傳差異在不同組合間可變化較大。因此,從線性相關的計算原理可知,由于親本選配的隨意性,有些情況下去除雙親之和最小的一些組合,可顯著提高雜種一代離親值或雜種優勢與中親值相關性。當然與以下原因有關。如果雜交試驗中存在很多如下類型組合,因各種養分的吸收速度或(臨界)環境消耗率較小導致雙親質量(之和)越小,雜種一代質量由于各種養分進入臨界點同步性改善而獲得的離親值反向同步增大受到限制,因此離親值與中親值或雙親之和的反相關性減弱或不存在。這在公開發表的文獻中十分常見。例如桑世飛等[24]油菜雜交試驗原有組合46個,去除產量雙親之和最小的6個組合,雜種一代產量離親值與中親值相關系數由原-0.454 8(P<0.002)變為-0.564 3(P<0.005)。王建軍等[25]油菜雜交試驗原有雜交組合36個,去除籽粒產量雙親之和最小的2個組合,籽粒產量離親值與雙親之和相關系數由原-0.326 3(P<0.1)增加為-0.474 3(P<0.005)。唐梅[26]水稻雜交試驗重慶試點原有組合35個,現去除單株粒質量雙親之和最小的8個組合,離親值與雙親之和的相關系數由原-0.183 0增加為-0.539 4(P<0.005)。由于植株體主要由大量元素構成,但它對大量元素和少(微)量元素的獲取是相互協同和相互制約的關系,因試驗環境、遺傳背景、目標性狀及取樣時間不同,雜種優勢與親本生物體質量的相關性存在可解釋的變化,但在生長后期或環境養分相對虧缺時,負相關性幾乎是必然的,有興趣的讀者可以去驗證。
3 結論與討論
光能截獲與環境養分獲取兩種模型在解釋雜種優勢形成時可存在兼容性。例于在某種條件下大田作物生長早期,群體葉面積指數很小,單株葉面積與單位葉面積截獲光能在雙親間不易形成反向變化關系,甚至由于葉片形態等的差異,這兩個因子在雙親間呈同向變化。因此,根據前文的結果,這時雜種優勢大多數組合為零,甚至出現負向雜種優勢。從養分獲取模型來說,由于此時大多數親本仍處于養分飽和期或臨界點附近,所以也不易形成正向雜種優勢。大田作物要獲得很高的產量,必須群體葉面積、葉片功能期、光合效率均表現較好;產量很低時,可允許這3個因子優劣的不同組合。從光能截獲模型考慮,雙親產量越高時越不易形成雜種優勢;甚至存在一些組合雙親產量之和較大,表現為一個親本產量很高,另一個產量一般,光能截獲構成因子在雙親間同向變化從而形成負向雜種優勢。雙親產量減小時,易獲得光能截獲的反極因子效應[3],形成雜種優勢;從養分獲得模型考慮,雙親產量越高,養分吸收和環境供給相容性越好,遺傳相似性越好,越不易獲得雜種優勢,正如上文所述。從上文不難看出,光能截獲的反極因子效應和養分獲取的雜種優勢形成機理使植物體整體或部分質量存在正向中親優勢或雜種優勢近似于一種本能或必然的自然現象。這從公開發表的各種作物雜交試驗的相關文獻中能夠發現[3-9,18-26],幾乎所有雜交試驗中,各組合中親優勢優勢平均值大于零或者正向中親優勢的組合比例大于50%,占很大比例的試驗中均接近100%。但對于多年生喬本植物而言,雜種一代早期群體光能截獲的雜種優勢不能適時地轉換為生殖器官相關性積累,絕大部分用于加快群體營養生長或葉面積指數增加,以致群體過于繁茂引起的平均單位葉面積截獲光能下降或低于雙親,經過較長時間或數年后,植株營養生長量例于主莖高度及莖圍、材積等性狀正向中親優勢或雜種優勢獲得的必然性顯著下降,各組合雜種優勢平均值或趨于零[27-28]。目前,對于雜種優勢產生機理的各種解釋,如顯性、超顯性、上位性效應、基因網絡、遺傳振動等假設,都未能觸及雜種優勢的必然性,更由于它們所涉及的遺傳方式均具有隨機性,也不能形成雜種優勢的必然性。與目前所有研究與雜種優勢相關性的研究方法相比[4-5,8,15-16,21-26]本文所揭示的雜種優勢與有關因子的相關性顯著度大幅度提高,而且是恒定的,或是可解釋的變化。這是因為本文所研究的相關性是揭示雜種優勢內在機理的必然相關性,而不是在以往研究中從尋求相關性過程中去試圖了解雜種優勢形成機理,是兩個相反的過程。本文研究不設定遺傳背景和試驗環境,強化環境效應在雜種優勢形成中的地位,避免了以往研究當試驗條件改變時結論不再適用的情況。很顯然,環境相容性包括養分的供給數量、空間分布及供給時期等方面,還包括溫度、光照、病害、土壤濕度等方面。筆者認為,本文的研究方法或是雜種優勢形成機理終極揭秘一個全新的重要視角。
參考文獻:
[1]張義榮,姚穎垠,彭惠茹,等.植物雜種優勢形成的分子遺傳機理研究進展[J].自然科學通訊,2009,19(7):697-703.
[2]何光明,鄧興旺.植物雜種優勢分子機理研究:機遇和挑戰[J].中國基礎科學植物科學專刊,2016(1):28-34.
[3]潘春桂.基于反極因子的雜種優勢形成途徑[J]. 陜西農業科學,2018,64(4): 71-73,76.
[4]司洪華 .利用玉米雜交當代雜種優勢預測成株期雜種優勢的研究[D].沈陽:沈陽農業大學, 2006.
[5]吳才君,曹家樹,董德坤,等. 蕓薹種蔬菜雜交種及其親本蓮座期基因差異表達與雜種優勢的關系[J].中國農業科學,2004,37(11):1654-1659.
[6]趙禹凱,魏建和,楊成民,等.桔梗主要農藝性狀的雜種優勢分析[J]. 中藥材,2010, 33(10):1523-1527.
[7]余政軍,邵小強,肖新宇,等.棉花主要性狀的遺傳相關性分析[J].湖南農業科學,2014(1):17-20.
[8]楊祥波. SSR分子標記遺傳距離預測大豆親本雜種優勢的初步研究[D].長春:吉林農業大學,2008.
[9]李素萍.食用向日葵雜種優勢及配合力研究[D].呼和浩特:內蒙古農業大學,2006.
[10]李秋元,孟德順. Logistic曲線的性質及其在植物生長分析中的應用[J].西北林學院學報,1998,8(3): 81-86.
[11]莫紅,陳瑛.基于Logistic方程的植物生長過程模型與最優分析[J].焦作大學學報, 2006(4):70-71.
[12]羅林會.不同配方水培營養液對皺葉留蘭香生長發育的影響[J].江蘇農業科學, 2009(3):227-228.
[13]馬謙.水稻苗期雜種優勢分析及赤霉素與苗期雜種優勢生物學基礎的關系研究[D].武漢:華中農業大學,2011.
[14]余麗霞.陸地棉主要經濟性狀的遺傳及雜種優勢研究[D].武漢:華中農業大學,2005.
[15]王章奎 .普通小麥雜種與親本間根系基因表達差異與雜種優勢分子機理[D].北京:中國農業大學, 2004.
[16]溫玲. 甜瓜雜種優勢早期預測指標的篩選[D]. 哈爾濱:東北農業大學,2003.
[17]蔡麗艷,石鳳翎,陳海玲.不同苜蓿雄性不育雜交組合牧草產量雜種優勢分析[J].中國草地學報,2013,35(2):24-30.
[18]林欣立,李寅安,王美菊.蘿卜雜種優勢與配合力的研究[J].中國農業科學,1986,19(20):54-60.
[19]潘春桂. 一種提高作物產量雜種優勢的親本選配方法—以棉花為例[J].安徽農業科學,2017,45(7):28-29,99.
[20]徐靜斐,汪路應.水稻雜種優勢與配合力的初步研究[EB/0L].安徽農學院學報, http://www.cnki.com.cn/Article/CJFD
Total-ANHU197900002.htm.
[21]吳利民,倪中福,王章奎,等. 小麥雜種及其親本苗期葉片家族基因差異表達及其與雜種優勢關系的初步研究[J]. 遺傳學報,2001,28(3):256-266.
[22]梁耀平,王世杰,陳豫梅,等.西瓜親本遺傳差異與雜種優勢的相關性[J].江蘇農業學報,2012, 28(3):622-626.
[23]閻冬生. 大豆籽粒基因差異表達模式與雜種優勢關系的研究[D].長春:吉林大學,2008.
[24]桑世飛,王會,梅德圣,等.利用全基因組SNP芯片分析油菜遺傳距離與雜種優勢的關系[J].中國農業科學, 2015,48(12):2469-2478.
[25]王建軍,雍洪軍,張曉聰,等.12幾個外來玉米群體與我國主要種質配合力和雜種優勢分析[J].作物學報,2012,38(2):2170-2177.
[26]唐梅.中秈雜交水稻雜種優勢的分子際記研究[D].重慶:西南大學,2000.
[27]張一,周志春,金國慶,等.馬尾松雙列雜交親本遺傳距離與雜種優勢相關性分析[J]南京林業大學學報, 2010,34(1):9-14.
[28]王趙民,陳益泰.杉木主要性狀配合力分析及雜種優勢利用[J].林業科學研究, 1998,11(6):614-624.
