脊髓CSF-1誘發小膠質細胞活化在長春新堿誘導神經病理性疼痛大鼠中的作用
2020-08-02 10:49:33付寶軍姜靜靜黃玉瓊林宗航李恒
新醫學 2020年7期
付寶軍 姜靜靜 黃玉瓊 林宗航 李恒
【摘要】目的 探討脊髓集落刺激因子1(CSF-1)誘發小膠質細胞活化在長春新堿誘導神經病理性疼痛大鼠中的作用。方法 采用隨機數表法將30只健康雄性SD大鼠分為3組(每組10只):對照組(Control組)、化學治療痛組(CINP組)、化學治療痛+ CSF-1中和抗體組(CINP+anti組)。化學治療藥物誘導的神經病理性疼痛動物模型采用隔日腹腔注射長春新堿125 μg/kg(共計4次)的方法建立,CINP+anti組在CINP組的給藥基礎上加予CSF-1中和抗體。于首次注射長春新堿前及注射后1、3、5、7 d采用機械縮足反射閾值(MWT)和熱縮足反射潛伏期(TWL)評價大鼠機械痛敏和熱痛敏;采用蛋白免疫印跡法檢測脊髓CSF-1以及脊髓小膠質細胞標志物離子化鈣結合適配分子1(Iba1)的表達情況;采用逆轉錄PCR法檢測脊髓CSF-1 mRNA和Iba1 mRNA的表達水平。結果 與Control組比較,CINP組大鼠在首次注射長春新堿后3、5、7 d的MWT和TWL更低(P均< 0.01);與CINP組比較,CINP+anti組首次注射長春新堿后5、7 d的MWT和TWL更高(P均< 0.01)。與Control組比較,CINP組脊髓CSF-1、Iba1表達上調(P均< 0.01);與CINP組比較,CINP+anti組CSF-1、Iba1表達下調(P均< 0.05)。與Control組比較,CINP組脊髓Iba1 mRNA表達上調(P < 0.01);與CINP組比較,CINP+anti組脊髓Iba1 mRNA表達下調(P < 0.01)。結論 長春新堿誘導神經病理性疼痛,其機制可能與大鼠脊髓CSF-1活化脊髓小膠質細胞有關。
【關鍵詞】長春新堿;神經病理性疼痛;小膠質細胞; 集落刺激因子1
【Abstract】Objective To investigate the role of spinal colony-stimulating factor 1 (CSF-1)-induced microglia activation in rat models with neuropathic pain induced by vincristine. Methods Thirty healthy male SD rats were divided into 3 groups by random number table method (10 rats in each group): control group (control group), chemotherapy-induced neuropathic pain group (CINP group) and chemotherapy-induced neuropathic pain+CSF-1 neutralizing antibody group (CINP+anti group). The CINP rat models were established by intraperitoneal administration of vincristine 125μg/kg on alternate days for 4 times. In the CINP+anti group, CSF-1 neutralizing antibody was supplemented. Mechanical allodynia and heat hyperalgesia were evaluated by mechanical withdrawal threshold (MWT) and thermal withdrawal latency (TWL) before and 1-, 3-, 5- and 7-d after initial administration of vincristine, respectively. The expression levels of CSF-1 and ionized calcium-binding adapter molecule 1 (Iba1) proteins were detected by Western blot. The expression levels of CSF-1 and Iba1 mRNA were measured by RT-PCR. Results Compared with the control group, the MWT and TWL at 3-, 5- and 7-d after vincristine administration were significantly decreased in the CINP group (both P < 0.01). The MWT and TWL at 5- and 7-d after vincristine administration in the CINP+anti group were significantly higher than those in the CINP group (both P < 0.01). Compared with the control group, the expression levels of CSF-1 and Iba1 in the spinal cord were significantly up-regulated in the CINP group (both P < 0.01). Compared with the CINP group, the expression levels of CSF-1 and Iba1 in the spinal cord were significantly down-regulated in the CINP+anti group (both P < 0.05). Compared with the control group, the expression of Iba1 mRNA was significantly up-regulated in the CINP group (P < 0.01). Compared with the CINP group, the expression of Iba1 mRNA was remarkably down-regulated in the CINP+anti group (P < 0.01). Conclusion The mechanism of neuropathic pain induced by vincristine may be related to the activation of spinal microglia induced by CSF-1 in rats.
【Key words】Vincristine;Neuropathic pain;Microglia;Colony-stimulating factor 1
長春新堿是一種常用化學治療藥物,被用于治療各種癌癥,特別是急性淋巴細胞白血病、霍奇金淋巴瘤和非霍奇金淋巴瘤。長春新堿與其他化學治療藥物一樣會伴不良反應,其中化學治療誘導的神經病理性疼痛(CINP)最為常見,這是一種劑量限制性毒性[1-2]。目前用于化學治療引起CINP的治療藥物僅限于抗驚厥藥、阿片類藥物和三環抗抑郁藥,但這些藥物的應用往往因其不可被接受的不良反應而受限[3-4]。因此闡明CINP發生機制,對于探索藥物作用靶點、開發有效的治療藥物具有重大現實意義。隨著對疼痛機制研究的不斷深入,細胞因子尤其集落刺激因子1(CSF-1)在CINP中的作用得到越來越多研究者關注[5-7]。近期的研究證實,在脊神經結扎、背部痛、關節炎疼痛模型中,CSF-1在慢性疼痛信息調制中扮演重要角色,但是其在化學治療藥物誘導CINP中的作用未見報道[8-10]。本研究采用長春新堿誘導CINP大鼠模型,探討長春新堿誘導機械痛敏和熱痛敏的可能機制。
材料與方法
一、材? 料
實驗動物為清潔級健康雄性Sprague Dawley(SD)大鼠30只,體質量200~230 g,由清遠市人民醫院實驗動物中心提供[生產許可證號:SYXK(粵)2019-0206]。注射用硫酸長春新堿(浙江海正藥業股份有限公司),兔抗大鼠CSF-1單克隆抗體、兔抗大鼠離子化鈣結合適配分子1(Iba1)單克隆抗體(Abcam公司,美國),PCR試劑盒(武漢博士德生物科技有限公司),von Frey細絲(Stoelting公司),熱痛刺激儀(Stoelting公司),CSF-1中和抗體(Abcam公司),TRIzol? reagen(Invitrogen公司),焦碳酸二乙酯、BeyoR TTM cDNA 逆轉錄試劑盒(上海碧云天生物技術有限公司南通分公司),實驗用PCR引物(天跟生化科技北京有限公司)。本實驗符合動物實驗的倫理要求。
二、方 法
1.動物分組與處理
將30只大鼠適應飼養1周后,將其按隨機數表法分為3組各10只:對照組(Control組)、化學治療痛組(CINP組)、化學治療痛+CSF-1中和抗體組(CINP+anti組)。整個實驗過程中動物自由攝食和飲水,分籠單獨飼養, 室溫(22±1)℃, 光照周期12 h(7:00 ~ 9:00 pm 光照,19:00 ~ 7:00 黑暗)。
2. 鞘內置管
參照Yaksh等[11]報道的方法對大鼠實施鞘內置管, 置管成功后觀察其肢體運動情況, 有運動功能障礙大鼠將被剔除后續實驗,經導管注射利多卡因后30 s內出現雙后肢麻痹現象即確認為導管位置正確,置管5 d后測試大鼠無感覺及運動異常即可用于實驗 。
3.實驗動物模型建立及給藥方法
對大鼠進行隔日腹腔注射長春新堿,注射量為125 μg/kg(共計 4 次),注射當日視為 1 d,通過檢測大鼠痛閾值變化確定化學治療藥物誘導的CINP模型建立成功。CINP+anti組從1 d開始連續7 d于行為學測試前30 min鞘內給予CSF-1中和抗體(10 μg/10 μl),每次給藥后用10 μl生理鹽水沖洗聚乙烯管。
4. 行為學的測定
參照Chaplan 等[12]的方法測定機械縮足反射閾值(MWT),以Up-down法推測閾值,并在閾值上下各刺激5次,用中位數法計算50%的反應閾值。參照Hargreaves等[13]的方法測定熱縮足反射潛伏期(TWL), 為防止組織損傷設定自動切斷時間為20 s。在整個實驗過程中熱刺激強度保持一致,測定5次TWL(間隔3 min)取平均值。在給藥前、給藥后1、3、5、7 d分別采用MWT和TWL評價大鼠機械痛敏和熱痛敏。
5. 蛋白免疫印跡法檢測脊髓CSF-1、Iba1蛋白
采用戊巴比妥鈉50 mg/kg麻醉大鼠后處死,取出其脊髓L4 ~ 5節段進行勻漿(加入裂解液),于4℃下12 000轉/分離心5 min,并進行二辛可寧酸法(BCA)蛋白定量,每份樣品各取20 μg蛋白質。配置分離膠和濃縮膠,當溴酚藍染料前端電泳至分離膠末端處時即停止電泳,轉膜后封閉2 h, 加入β-actin(兔抗大鼠,1∶2000)、CSF-1(兔抗大鼠,1∶1000)、? Iba1(兔抗大鼠,1∶1000)一抗,4℃孵育過夜后洗膜3次(每次10 min)。加入辣根過氧化物酶(HRP)標記的山羊抗兔IgG(1∶3000)室溫孵育后洗膜、顯色、曝光、顯影,采用Image J軟件檢測目的蛋白條帶及β-actin蛋白條帶的光密度值,以目的蛋白條帶灰度值/β-actin蛋白條帶灰度值作為目的蛋白表達量。將3張以上相同趨勢灰度值取平均值記為本次實驗結果。
6. 逆轉錄-PCR測定脊髓CSF-1 mRNA、Iba1 mRNA??注射長春新堿后7 d,每組各取3只大鼠實施安樂死后進行檢查。用TRIzol? reagent提取大鼠L4 ~ 6脊髓總RNA,逆轉錄為互補DNA。用ΔΔCT 法來測定CSF-1 mRNA、CSF-1受體mRNA、Iba1 mRNA含量。引物合成為:CSF-1上游引物:5-TGCTAAGTGCTCTAGCCGAG-3;下游引物5- CCCCCAACAGTCAGCAAGAC-3。Iba1上游引物:5-CCATGACCTTCCAAGAGAATGC-3;下游引物:5-ACCGGCTTGTGCTGTAGTC。β-actin上游引物:5-CGTTGACATCCGTAAAGACCTC-3;下游引物:5-TAGGAGCCAGGGCAGTAATCT-3。擴增條件:94℃預變性5 min、94℃ 30 s、54℃ 30 s、72℃ 20 s,共45個循環,72℃延伸10 min。計算目的基因表達灰度/β-actin灰度值作為目的基因表達量。
三、統計學處理
采用GraphPad Prism 7.0軟件進行統計學處理,實驗數據均以表示,行為學數據結果采用重復測量方差分析,同一時間點多重比較(Control組與CINP組;CINP組與CINP+anti組)采用Sidak法;蛋白免疫印跡、逆轉錄PCR數據結果采用單因素方差分析,多重比較(Control組與CINP組;CINP組與CINP+anti組)采用Tukey法。P < 0.05為差異有統計學意義。
結果
一、制模情況
本實驗中的大鼠首次給藥后3 d 的MWT和TWL開始明顯降低,并出現跛行、抬足、舔舐足底增多以及自發嘶叫等疼痛行為學變化,直到本實驗觀察結束,表明CINP模型制備成功,無被剔除的大鼠。
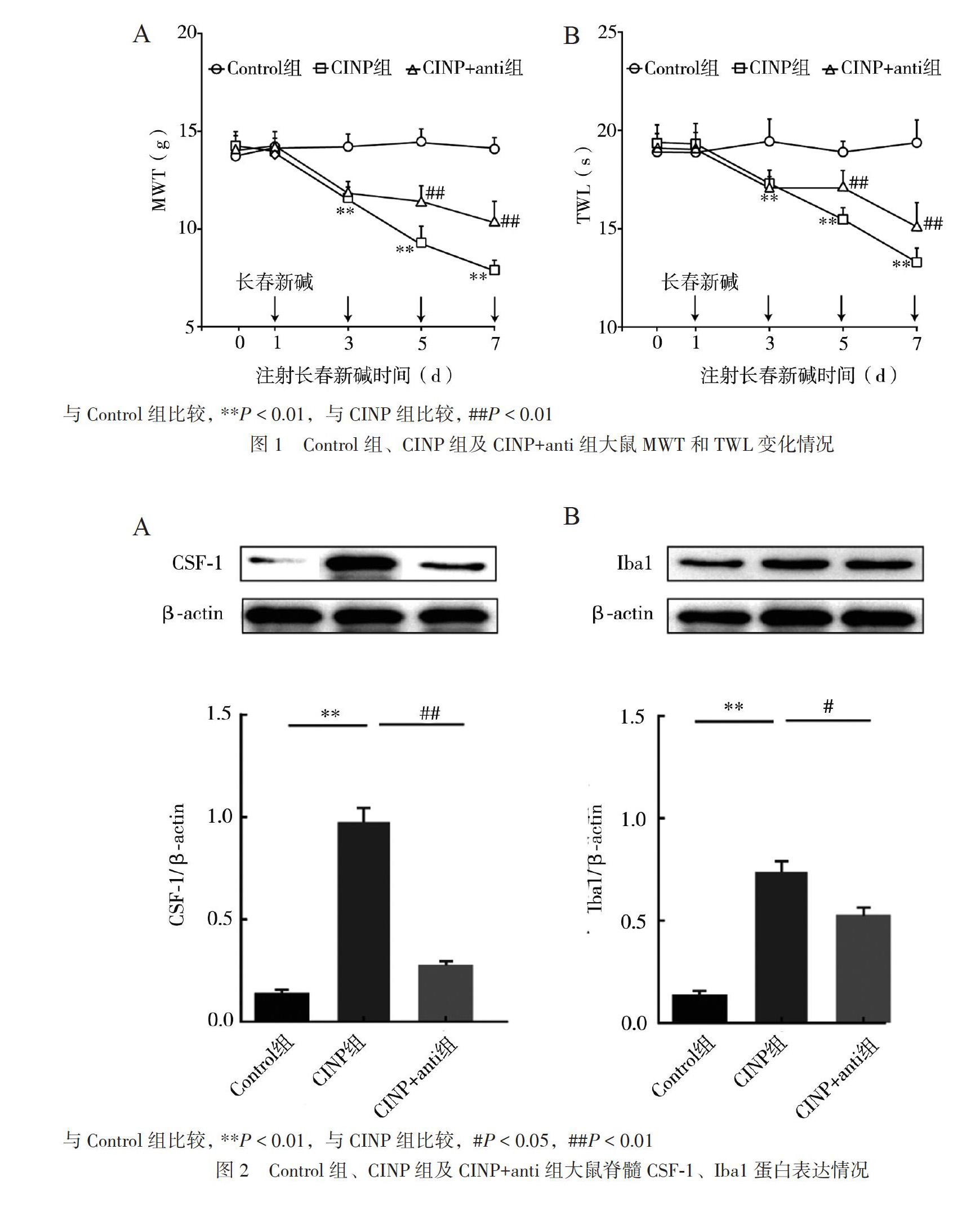
二、Control組、CINP組及CINP+anti組大鼠 MWT和TWL 變化情況
重復測量資料方差分析結果:時間和分組之間存在交互效應[MWT(F = 27.11,P < 0.01)和TWL(F = 28.61,P < 0.01)],故分析簡單效應與Control組比較,CINP組大鼠在3、5、7 d, MWT[3 d:(12.6±0.5)g,P < 0.01;5 d:(9.3±0.8)g,P < 0.01;7 d:(8.1±0.7)g,P < 0.01)和TWL[3 d: (17.1±0.6)s,P < 0.01;5 d:(15.5±0.6)s,P < 0.01;7 d:(12.9±0.7)s,P < 0.01]均降低,同時大鼠跛行、抬足、舔舐足底、自發嘶叫明顯增加。與CINP組比較,CINP+anti組大鼠在5、7 d MWT[5 d:(11.7±1.1)g,P < 0.01;7 d:(11.5±0.5)g,P < 0.01]和TWL[5 d:(17.2±0.6)s,P < 0.01;7 d:(14.8±0.5)s,P < 0.01]均升高;同時大鼠跛行、抬足、舔舐足底、自發嘶叫明顯減少,見圖1。
三、Control組、CINP組及CINP+anti組大鼠大鼠脊髓CSF-1和Iba1蛋白表達情況各組大鼠的CSF-1蛋白(F = 283.20, P < 0.01)和Iba1蛋白(F = 21.25, P < 0.01)表達差異均有統計學意義。與Control組比較,CINP組脊髓CSF-1蛋白(0.85±0.10)表達上調(P < 0.01);與CINP組相比,CINP+anti組脊髓CSF-1蛋白(0.25±0.05)表達下調(P < 0.01),見圖2A。與Control組相比,CINP組脊髓Iba1蛋白(0.75± 0.12)表達上調(P < 0.01);與CINP組相比,CINP+anti組脊髓Iba1蛋白(0.53±0.06)表達下調(P = 0.012),見圖2B。
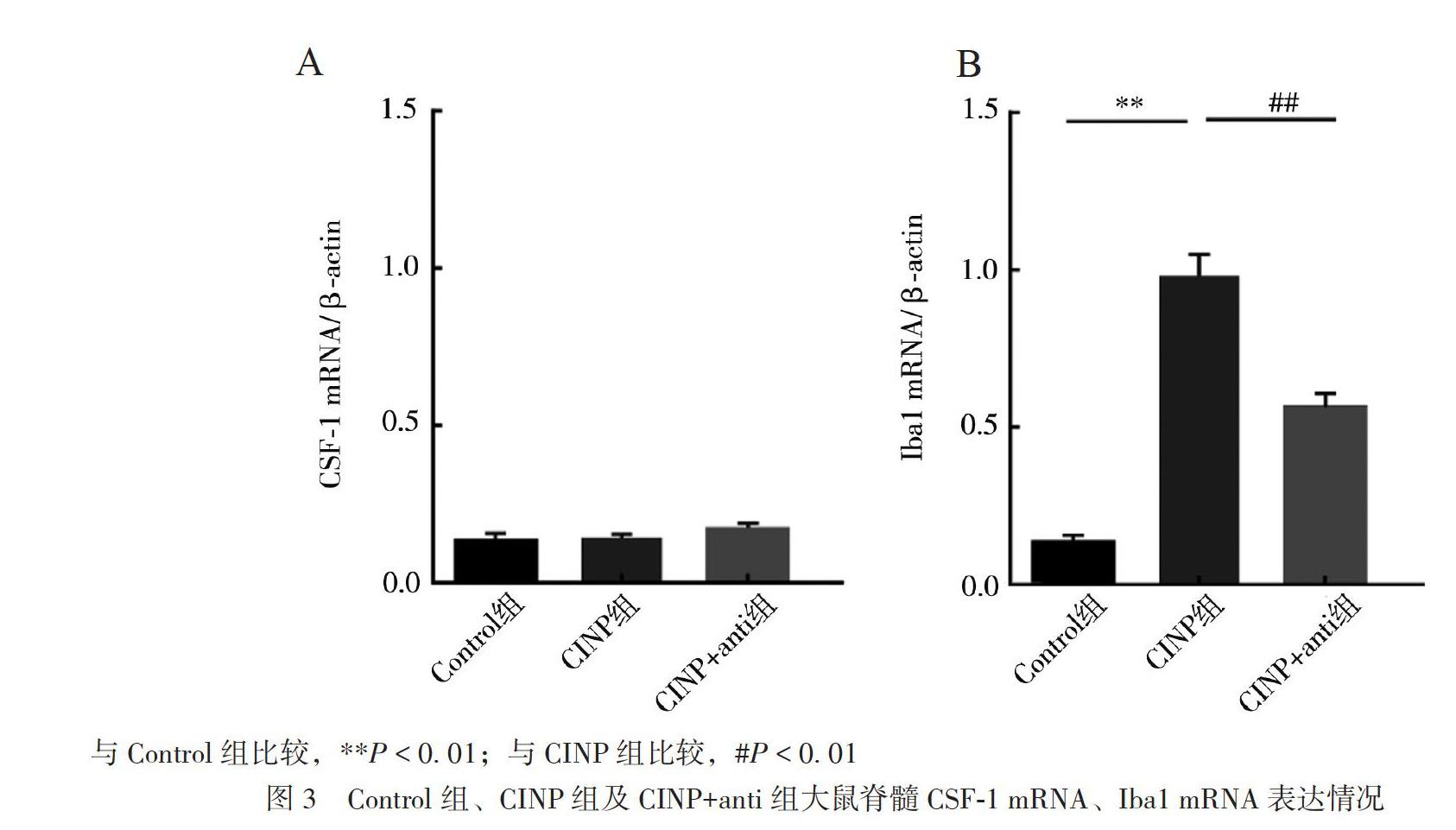
三、各組大鼠脊髓CSF-1、Iba1 mRNA表達情況
各組脊髓CSF-1mRNA表達差異無統計學意義(F = 1.71,P = 0.256),見圖3A。各組脊髓Iba1 mRNA表達差異有統計學意義(F = 108.50,P< 0.01),與Control組比較,CINP組脊髓Iba1 mRNA表達(0.95±0.09)上調(P < 0.01);與CINP組比較,CINP+anti組脊髓Iba1 mRNA表達(0.63± 0.06)下調(P < 0.01),見圖3B。
討論
本研究采用許愛軍等[14]介紹的方法制備大鼠CINP模型,該模型由于可操作性強、重復性好、與臨床CINP特征有相似之處等優點,已被廣泛應用于驗證藥物和探討機制的動物實驗中。本實驗的CINP模型制備成功。
CSF-1是一種細胞因子,它通過與Ⅲ型受體酪氨酸激酶偶聯CSF-1受體結合來發揮作用,在調節單核細胞、巨噬細胞和小膠質細胞存活、增殖和分化中發揮重要作用[15]。越來越多證據顯示CSF-1在CINP中發揮重要作用[5-6]。我們的研究結果表明,與Control組比較,CINP組大鼠脊髓CSF-1蛋白表達上調,但兩者脊髓CSF-1 mRNA 表達差異無統計學意義,提示可能存在其他部位CSF-1蛋白向脊髓轉運的情況,本課題組(2019年)進行的前期實驗顯示,在構建CINP模型前結扎L4 ~ 5背根,脊髓CSF-1降低與正常對照組無差異,也進一步證實背根節CSF-1向脊髓轉運。另有研究顯示缺血痛模型建立后6 h脊髓背角活化的星形膠質細胞產生CSF-1,相反,星形膠質細胞代謝抑制劑氟代檸檬酸顯著抑制缺血6 h后CSF-1上調,其CSF-1產生部位和我們實驗的差異可能與使用模型以及動物種屬不同有關[16]。既然存在CSF-1向脊髓轉運,那么CSF-1如何發揮作用?最近的研究表明,脊神經結扎大鼠背根節,可使初級感覺神經元中CSF-1促進脊髓小膠質細胞的激活,以及促發促傷害性基因[8]。Iba1是小膠質細胞的標志物,是一個17 kDa的EF手性蛋白,在小膠質細胞的活化過程中表達升高。在本研究中,予大鼠腹腔注射長春新堿后,CINP組脊髓Iba1蛋白、Iba1 mRNA均升高,與CINP組相比,CINP+anti組脊髓Iba1蛋白、Iba1 mRNA均降低,這提示脊髓CSF-1蛋白可能活化了脊髓背角小膠質細胞介導CINP發生。
綜上所述,長春新堿誘導的CINP,其機制可能與脊髓CSF-1激活小膠質細胞有關。然而在本實驗中脊髓CSF-1蛋白表達升高是否源于背根節或其他部位的轉運還有待今后的實驗進一步證實。
參 考 文 獻
[1] 梁承華,鄭曉明, 張亞琴, 雷普潤, 蔡梓凱, 魏波. 含奧沙利鉑化學治療方案對Ⅱ期結直腸癌患者肝纖維化的影響. 新醫學, 2019, 50(6):427-431
[2] Brewer JR, Morrison G, Dolan ME, Fleming GF. Chemotherapy-induced peripheral neuropathy: current status and progress. Gynecol Oncol,2016, 140(1):176-183.
[3] Busse JW, Wang L, Kamaleldin M, Craigie S, Riva JJ, Montoya L, Mulla SM, Lopes LC, Vogel N, Chen E, Kirmayr K, Oliveria KD, Olivieri L, Kaushal A, Chaparro LE, Oyberman I, Agarwal A, Couban R, Tsoi L, Lam T, Vandvik PO, Hsu S, Bala MM, Schandelmaier T, Scheidecker A, Ebrahim S, Ashoorion V, Rehman Y, Hong PJ, Ross S, Johnston BC, Kunz R, Sun X, Buckley N, Sessler DI, Guyatt GH. Opioids for chronic noncancer pain: a systematic review and meta-analysis. JAMA, 2018, 320(23):2448-2460.
[4] Fornasari D. Pharmacotherapy for neuropathic pain: a review. Pain Ther, 2017, 6(Suppl 1):25-33.
[5] Zhou LJ, Peng JY, Xu YN, Zeng WJ, Zhang J, Wei X, Mai CL, Lin ZJ, Liu Y, Murugan M, Eyo UB, Umpierre AD, Xin WJ, Chen T, Li MG, Wang H, Richardson JR, Tan Z, Liu XG, Wu LJ. Microglia are indispensable for synaptic plasticity in the spinal dorsal horn and chronic pain. Cell Rep, 2019, 27 (13):3844-3859.e6.
[6] Zhang F, Wang YY, Liu Y, Han H, Zhang DD, Fan XZZ, Du XN, Gamper N, Zhang HL. Transcriptional regulation of voltage-gated sodium channels contributes to GM-CSF-induced pain. J Neurosci, 2019, 39(26):5222-5233.
[7] Boakye PA, Rancic V, Whitlock KH, Simmons D, Longo FM, Ballanyi K, Smith PA. Receptor dependence of BDNF actions in superficial dorsal horn: relation to central sensitization and actions of macrophage colony stimulating factor 1. J Neuro-physiol,2019, 121(6):2308-2322.
[8] Guan ZH, Kuhn JA, Wang XD, Colquitt B, Solorzano C, Vaman S, Guan AK, Reinsch ZE, Braz J, Devor M, Werner SLA, Lanier LL, Lomvardas S, Basbaum A. Injured sensory neuron-derived CSF1 induces microglial proliferation and DAP12-dependent pain. Nat Neurosci, 2016, 19(1):94-101.
[9] Yang G, Chen LH, Gao ZH, Wang Y. Implication of microglia activation and CSF-1/CSF-1Rpathway in lumbar disc degeneration-related back pain. Mol Pain, 2018, 14:1744806918811238.
[10] Saleh R, Lee MC, Khiew SH, Louis C, Fleetwood AJ, Achuthan A, Forster I, Cook AD, Hamilton JA. CSF-1 in inflammatory and arthritic pain development. J Immunol, 2018, 201(7):2042-2053.
[11] Yaksh TL, Rudy TA. Chronic catheterization of the spinal subarachnoid space. Physiol Behav, 1976, 17(6):1031-1036.
[12] Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Method, 1994, 53(1):55-63.
[13] Hargreaves K, Dubner R, Brown F, Flores C, Joris J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain, 1988, 32(1):77-88.
[14] 許愛軍, 曹菲, 田玉科. 長春新堿誘發外周神經病理性疼痛模型的建立. 中國疼痛醫學雜志, 2008, 21(9): 647-651.
[15] Liu J, Ying YY, Wang S, Li JY, Xu JQ, Lv PP, Chen JH, Zhou CY, Liu YF, Wu YQ, Huang Y, Chen Y, Chen LF, Tu SJ, Zhao W, Yang M, Hu YJ, Zhang RJ, Zhang D. The effects and mechanisms of GM-CSF on endometrial regeneration. Cytokine, 2020, 125:154850.
[16] Tang Y, Liu L, Xu D, Zhang WS, Zhang Y, Zhou JS, Huang W. Interaction between astrocytic colony stimulating factor and its receptor on microglia mediates central sensitization and behavioral hypersensitivity in chronic post ischemic pain model. Brain Behav Immun, 2018, 68:248-260.
(收稿日期:2020-01-20)
(本文編輯:洪悅民)
