分子標記輔助選擇Pi9基因改良湘晚秈11號的稻瘟病抗性
2020-08-13 09:07:38李枳蒙賴怡帆吳婷婷李博文劉雄倫
作物研究 2020年4期
關鍵詞:水稻
李枳蒙,張 婷,李 典,黃 俊,賴怡帆,吳婷婷,李博文,劉雄倫,2,3*
(1 湖南農業大學農學院,長沙 410128;2 作物基因工程湖南省重點實驗室,長沙 410128;3 水稻油菜抗病育種湖南省重點實驗室,長沙 410128)
水稻(Oryzasativa)是我國乃至世界最主要的糧食作物之一,地球一半以上人口以稻米為食。由子囊真菌Magnaporthegrisea引起的水稻稻瘟病嚴重危害水稻正常的生長發育,影響稻米產量和品質,稻瘟病爆發年份可造成10%~30%的產量損失,嚴重時甚至絕收[1,2]。目前,防治稻瘟病的主要途徑是化學防治和種植抗病水稻品種,但化學防治成本較高且不環保,培育和推廣抗瘟水稻新品種是降低稻瘟病危害的上策[3,4]。稻瘟病抗性屬典型的質量—數量性狀,利用已克隆或精細定位的主效抗性基因,開發高效分子標記進行輔助選擇,能最大限度地提高稻瘟病抗性育種的效率[5,6]。Pi9是一個廣譜持久抗稻瘟病基因,來源于小粒野生稻(Oryzaminuta),經遠緣雜交導入到秈稻品系75-1-127,被定位在水稻第6號染色體的Pi2/9位點且被成功克隆[7,8]。本研究以75-1-127為Pi9基因供體親本,以高感稻瘟病的優質常規晚稻品種湘晚秈11號為受體親本及輪回親本,利用Pi9基因特異高效分子標記,開展標記輔助選擇育種實踐,改良湘晚秈11號的稻瘟病抗性。
1 材料與方法
1.1 試驗材料
水稻材料:Pi9基因供體親本75-1-127;受體及輪回親本湘晚秈11號;感病對照品系及稻瘟病誘發品種CO39。
稻瘟菌菌株:來自國內外不同稻區的24份稻瘟菌菌株,用于室內人工接種。
1.2 稻瘟病抗性表型鑒定及抗菌譜分析
室內接種抗性鑒定和抗菌譜分析:將水稻材料75-1-127、湘晚秈11號、CO39播種于育秧基質,26~28 ℃生長至4葉期,用0.2‰的 Tween 20水溶液將稻瘟菌菌株分生孢子配成濃度約1×105個/mL的懸浮液,活體噴霧接種;接種后用黑布覆蓋,于26 ℃、相對濕度>90%條件下培養24 h后,繼續在同溫濕度、12 h光照下誘導發病5~7 d;當CO39葉片出現明顯病斑時觀察統計各材料抗性表型,參照文獻[9]中苗瘟調查分級標準,將0~2級劃分為抗病類型,3~5級劃分為感病類型,并計算各供試材料對不同菌株的抗性頻率(抗性頻率=接種后不致病菌株數÷接種總菌株數×100%)。
田間病圃抗性鑒定:5月中旬將供試水稻材料和誘發品種CO39浸種催芽后播于湖南瀏陽大圍山自然病圃。6月中旬鑒定苗瘟抗性表型,9月中旬鑒定穗瘟抗性表型。
1.3 高效分子標記鑒定與基因型分析
利用課題組已開發的Pi9基因內的功能標記Ins2-1(引物序列為Forward:5′-AACTGTTTTAAACACTCGGGTGGATA-3′;Reverse:5′-CACGATGATAACTTGGGCTAGGGCG-3′)[10],分析75-1-127與湘晚秈11號之間的多態性,并將其用于標記輔助選擇育種實踐中基因型的鑒定。抽穗前取水稻葉片約0.2 g于2.0 mL 滅菌離心管中,加入液氮研磨成粉末狀,采用CATB法[11]提取總DNA;利用標記引物對各樣品DNA進行PCR擴增;擴增產物經1.0%瓊脂糖凝膠電泳后分析標記基因型。10 μL PCR反應體系組成:DNA模板1.2 μL,2 pmol/μL primer pairs 1.0 μL,10×Buffer 1.0 μL,2.5 mmol/L dNTPs 0.2 μL,5 U/μL Taq酶0.1 μL,ddH2O 6.5 μL。PCR反應程序:94 ℃預變性5 min;94 ℃變性50 s,56 ℃退火50 s,72 ℃延伸40 s,35個循環;72 ℃延伸3 min;16 ℃保存。
1.4 回交自交育種群體構建
用湘晚秈11號作母本,75-1-127作父本,雜交獲得F1代;用湘晚秈11號作輪回親本,在長沙和三亞兩地連續回交直至獲得BC6F1代,自交兩代獲得BC6F3群體。回交群體構建具體方法:自BC1F1代起,每世代田間種植約60株,取單株葉片做標記基因型分析,選農藝性狀與受體親本接近的陽性單株回交,混合收種,獲得下一世代。
2 結果與分析
2.1 供試水稻材料的稻瘟病抗性表現及抗菌譜
根據室內接種結果(表1),75-1-127除了對湘2007A-7、ROR1這2份菌株感病外,其余均為抗病,抗性頻率為91.7%;湘晚秈11號對24份稻瘟菌菌株中的7份表現為抗病,其余17份均為感病,抗性頻率為29.2%;感病對照CO39對所有菌株均表現感病,抗性頻率為0。由田間病圃抗性鑒定結果(圖1)可以看出:受體親本湘晚秈11號高感苗瘟和穗瘟;而75-1-127高抗苗瘟和穗瘟。湘晚秈11號抗菌譜窄、抗性水平低,其稻瘟病抗性急需改良;75-1-127抗菌譜廣、抗性水平高,Pi9基因具有很好的育種價值。
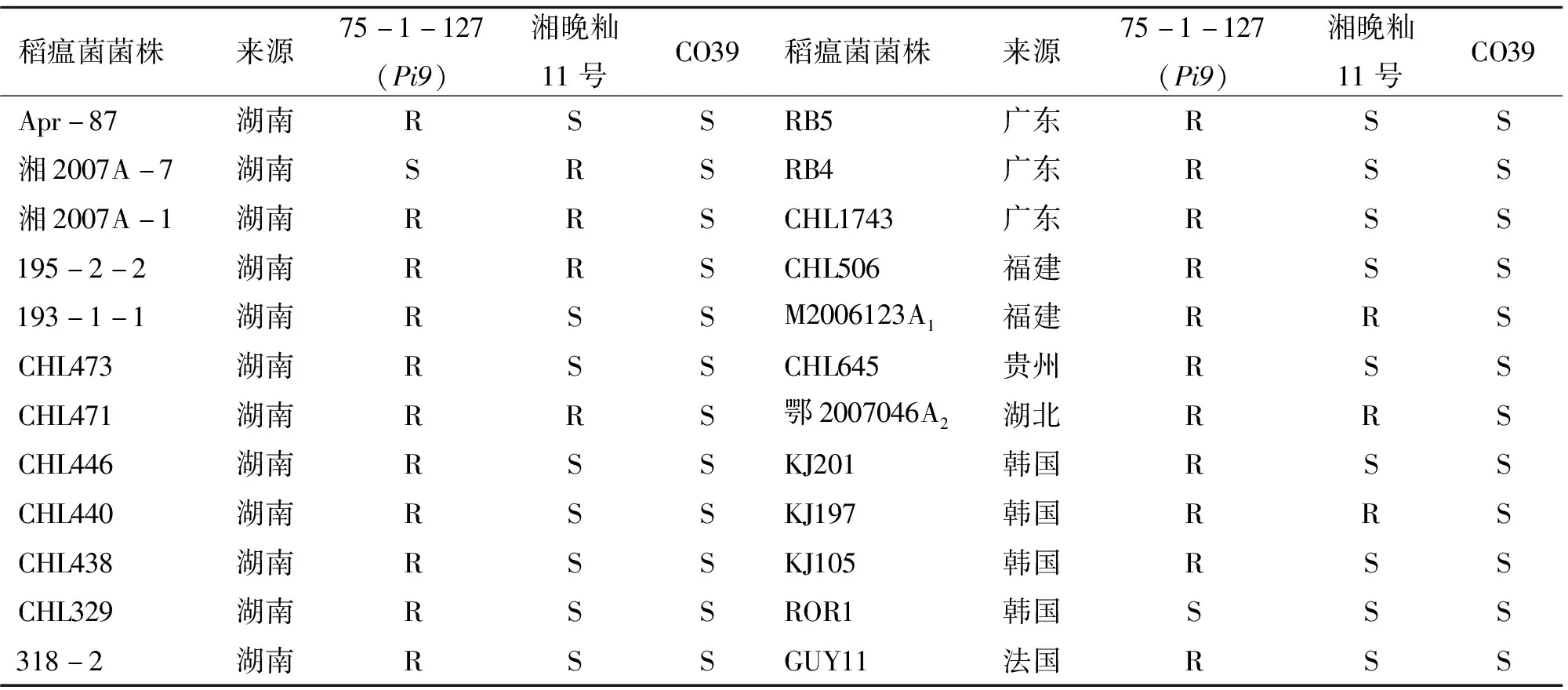
表1 供試材料對24份稻瘟菌菌株的抗譜比較

圖1 供試水稻材料田間病圃稻瘟病抗性表型
2.2 分子標記Ins2-1在雙親間多態性分析
雙親間多態性分析結果表明,Ins2-1是一個顯性DNA標記,利用該標記引物可從75-1-127基因組中擴增出1條532 bp的清晰條帶,而在湘晚秈11號基因組中無擴增產物(圖2A)。并且該標記PCR產物可以用瓊脂糖凝膠電泳檢測分析,操作簡單快速且成本較低。
2.3 湘晚秈11號改良抗病株系的選育
利用Ins2-1標記,從湘晚秈11號×75-1-127的BC1F1代起,開展分子標記輔助選擇育種實踐,于2016年獲得BC6F1群體,2018年篩選出11個BC6F3株系,2019年經標記基因型分析和田間病圃稻瘟病抗性鑒定,遴選出9個高抗稻瘟病的改良株系湘晚秈11號-Pi9(圖2),為進一步培育優異抗病優質水稻新品種奠定了基礎。

圖2 抗病改良株系湘晚秈11 號-Pi9的鑒定
3 討論
水稻稻瘟病抗性屬于典型的質量—數量性狀,抗性表型既受遺傳(基因)控制,又受病原菌、溫度、濕度、光照等環境因素影響,因此傳統育種的表型選擇往往不準確、效率低。分子標記輔助選擇育種是對標記基因型的鑒定和選擇,不受環境因素影響,相對于傳統育種更準確高效,現已被廣泛應用于作物遺傳改良,尤其是稻瘟病抗性育種。本研究鑒定出的顯性標記Ins2-1,可以在供體親本75-1-127與受體親本湘晚秈11號間產生明顯而穩定的多態性,選擇效率高,其擴增產物可用瓊脂糖凝膠電泳檢測,成本低、效率高,具有較好的應用前景。廣譜持久抗稻瘟病基因Pi9被成功克隆后,已被廣泛應用于水稻育種實踐。如廖花等[12]利用Pi9基因序列開發出共顯性標記SPL-1,通過分子標記輔助選擇,將Pi9基因導入秈稻恢復系R747 中,培育出了與受體親本遺傳背景高度相似的高抗稻瘟病新品系R747-Pi9,雜交種的稻瘟病抗性也同時改良;李永聰等[13]通過顯性標記Ins1-1的輔助選擇,成功改良了秈稻恢復系R389及其雜交種的稻瘟病抗性;鄒俊鋒等[14]利用共顯性標記Ins2-3改良了秈稻恢復系E32及其雜交種的稻瘟病抗性;殷得所等[15]通過Pi9基因緊密連鎖的共顯性STS標記的輔助選擇育種,成功改良了揚稻6號、R6547及其雜交種的稻瘟病抗性。
本研究獲得的9個含Pi9基因的湘晚秈11號改良抗病系,將用于進一步的農藝、產量、米質等性狀分析,以及不同生態區稻瘟病抗性鑒定,可望最終培育出高產優質抗瘟水稻新品種。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
