心臟高表達基因ZNF425在肺癌中功能的探究
2020-08-14 07:52:50揭雨婷劉鳳萍王志宏吳星瑤張雨璐鄧璐瑤肖梓淇丁浩東范雄偉袁婺洲
湖南師范大學自然科學學報 2020年4期
林 禮,揭雨婷,劉鳳萍,王志宏,吳星瑤,張雨璐,鄧璐瑤,劉 帆,肖梓淇,丁浩東,劉 超,范雄偉,袁婺洲
(湖南師范大學心臟發育研究中心,省部共建淡水魚類發育生物學國家重點實驗室,動物多肽藥物創制國家地方聯合工程實驗室,中國 長沙 410081)
就發病率(2018年全球有約210萬新增肺癌患者,占癌癥病例的11.6%,位居第一)和死亡率(每年約175.8萬例死亡)而言,肺癌是全球最常見的癌癥。事實上,肺癌也是癌癥死亡的首要原因[1]。非小細胞肺癌(NSCLC)約占所有肺癌病例的80%[2]。被診斷為非小細胞肺癌早期的患者,在經手術切除后的生存率相對較高[3]。但是,大多數患者在診斷時已經發展為晚期疾病,并且中位生存期從診斷之日起僅有18個月[4]。未經治療的轉移性非小細胞肺癌患者一年總生存率僅為10%,中位生存期為4~5個月。化學療法顯示了晚期NSCLC患者的生存率略有改善,可減輕癥狀并改善生活質量。實際上,從晚期疾病患者治療過程中不同化學治療劑與鉑化合物聯合治療產生的治療效果來看,不同的雙重測試之間沒有顯著差異[5]。迄今為止,已有許多研究證明MAPK信號通路的過度激活會增加癌細胞的增殖和侵襲能力。同時,也有研究表明KRAB鋅指蛋白與肺癌的發生息息相關。如:ZNF185的表達增加已參與腫瘤生長和轉移的調節[6]及ZNF322A積極調節α-adducin(ADD1)和cyclin D1(CCND1)的轉錄,從而促進肺癌的致癌性[7]。
鋅指(ZNF)基因家族是已知最大的基因家族之一,人類基因組中包含800多個編碼功能蛋白、剪接變異體和偽基因的ZNF轉錄本[8,9],它們可能參與發育、代謝、增殖和致癌等過程[10]。ZNF蛋白質包含多個ZNF域,通常組成集群的串聯重復序列。盡管有20種不同類型的ZNF領域,最常見的是Cys2-His2(C2H2)類[11],人類基因組中大約有700個基因含有C2H2結構[12]。當與DNA結合時, ZNF蛋白通過與KAP1 (KRAB相關蛋白1)的相互作用觸發轉錄抑制,也稱為TRIM28。TRIM28作為一種多分子實體的支架,該多分子實體包括異染色質蛋白1 (HP1),H3K9me3特異性組蛋白甲基轉移酶SETDB1和組蛋白去乙酰化復合物NuRD。這些蛋白通過觸發異染色質的形成來共同抑制轉錄[10,13,14]。
筆者從人類心臟cDNA文庫中克隆了一種新型的KRAB/C2H2型鋅指基因ZNF425。它編碼了一個包含752個氨基酸的鋅指蛋白,其中包含一個保守的KRAB結構域,以及19個C2H2型鋅指結構,且在物種間進化上高度保守。在前期研究中,已經發現ZNF425的mRNA主要存在于3個月大的人類胚胎中,而在其他階段都無法檢測到它的表達。通過構建EGFP-ZNF425融合蛋白,發現其主要定位于細胞核中,支持其轉錄調節蛋白的功能。同時熒光報告系統檢測顯示,ZNF425可以抑制SRE,AP-1和SRF的轉錄活性[15]。這些結果表明,ZNF425蛋白可能是MAPK信號通路的負調節因子,有介導細胞基因表達的功能。也有研究表明在急性T淋巴細胞白血病的Jurkat細胞系中瞬時過表達ZNF425時,能在不影響細胞周期的情況下抑制細胞的凋亡,并且出現ERK活性下降的現象[16]。這些研究都表明ZNF425與MAPK信號通路有著緊密的聯系。
ZNF425也有可能參與了肺癌的調控,但是對其研究甚少。本文先利用生物信息學的方法研究ZNF425與肺癌的相關性,基于癌癥和腫瘤基因圖譜(TCGA)數據庫、癌癥和正常基因表達譜公共數據庫(GEPIA),將ZNF425與肺癌兩種亞型LUAD(肺腺癌)和LUSC(肺鱗狀細胞癌)的數據進行分析,然后利用人類肺癌樣本IHC驗證生物信息學的分析結果,最后構建ZNF425過表達質粒,在人類肺腺癌肺泡基底上皮細胞A549細胞系過表達ZNF425,考察A549細胞中JNK和P38的活性。
1 材料與方法
1.1 材料
1.1.1 A549細胞 A549細胞系從湖南師范大學生命科學學院心臟發育研究中心獲得。
1.1.2 試劑 PRMI1640培養基、PBS(1×)、全式金生物有限公司的轉染試劑TransIntroTMTransfectionELReagent;唯世爾生物科技有限公司的轉染稀釋液、回收膠純化試劑盒;唯贊生物有限公司的T4-DNA連接酶;上海優寧維生物科技股份有限公司的β-actin抗體,JNK,p-JNK,P38及p-P38抗體試劑; Abcam生物有限公司的兔二抗。
1.2 方法
1.2.1 生物信息學分析 利用生物信息學相關網站GEPIA(http://gepia.cancer-pku.cn/)分析癌癥樣本中基因表達情況;利用TCGA(https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga)查詢NSCLC的相關資料;再利用Primerpremier5進行酶切位點分析及引物設計。
1.2.2 免疫組化 將前期使用支氣管內超聲引導針吸活檢術(EBUS-TBNA)取得的不同時期的組織放入3%多聚甲醛中固定,置于高濃度蔗糖溶液中脫水后進行石蠟包埋,然后切片,利用ZNF425特異性抗體進行免疫組化染色。以癌旁組織作為對照,使用Image J軟件進行定量分析,比較ZNF425蛋白水平變化情況。
1.2.3 pCMV-ZNF425-Tag2B過表達載體的構建 以ZNF425的ORF設計引物,經過PCR擴增得到目的基因片段,再進行純化回收。回收產物與pCMV-Tag2B載體通過T4-DNA連接酶連接,連接產物在16 ℃下放置過夜,利用一般轉化法篩選陽性單菌落,提取質粒利用雙酶切驗證,最后通過測序證實pCMV-ZNF425-Tag2B過表達載體構建成功。
1.2.4 Western Blot檢測 按表1比例配置與目的蛋白合適濃度的蛋白膠。首先在蛋白電泳槽中加入適量電泳緩沖液,再依次加入蛋白樣品及蛋白Marker。蛋白80 V恒壓下離開濃縮膠后將電壓增加至110 V繼續跑樣。跑膠結束后對比蛋白Marker切取所需蛋白轉移到NC膜上,在電轉膜儀中轉膜20 min,讓目的蛋白完全轉移至NC膜上。

表1 SDS-PAGE 蛋白膠配方Tab. 1 SDS-PAGE albumin glue
將NC膜在室溫下浸入30 mL 5%的脫脂牛奶中封閉2 h,封閉結束后用TBST液清洗。轉移NC膜在4 ℃環境下過夜或室溫下孵育一抗2 h(體積1∶1 000稀釋),完成后用TBST液清洗5~10 min,重復3次;轉移NC膜在室溫中孵育二抗2 h(體積1∶(3 000~5 000)稀釋),完成后用TBST清洗10 min,重復3次。利用顯影儀顯影, NC膜復蘇二次顯影,再利用蛋白印跡膜再生液室溫下震蕩孵育10 min,用TBST清洗10 min,重復3次,洗滌一抗-二抗復合物,孵育內參抗體。重復上述步驟,顯影。
2 結果
2.1 ZNF425與肺癌相關性的生物信息學分析
基于癌癥和腫瘤基因圖譜(TCGA)數據庫、癌癥和正常基因表達譜分析的公共數據庫(GEPIA),將ZNF425在肺癌兩種亞型LUAD(肺腺癌)和LUSC(肺鱗狀細胞癌)中的數據,按照其生物信息分析模塊構建分析路線圖進行自動運算分析,據此得到多個樣本中ZNF425在同一病患的癌旁組織和患病癌組織的差異表達,生成箱盒圖、生存圖等。首先分析肺癌中ZNF425基因表達差異性,并根據GEPIA生成的帶有散點的盒形圖來比較ZNF425在兩種肺癌亞型的表達,選擇TCGA正態進行差異分析和繪圖。圖1a是基于腫瘤基因組圖譜(TCGA)數據庫的ZNF425mRNA在NSCLC中的表達。將59例LUAD患者組織(n=483)和50例LUSC患者組織(n=486)中ZNF425mRNA的表達與正常肺組織中ZNF425的表達進行比較,結果筆者發現與正常肺組織相比,癌組織中ZNF425的表達有所降低。
然后基于GEPIA數據庫的病理分期數據具體分析ZNF425在肺癌不同時期的表達情況。GEPIA按照LUAD和LUSC癌癥類型和病理分期繪制ZNF425基因的表達譜(如圖1b)。可以發現ZNF425在肺癌Ⅲ期時表達略微下調。
最后基于GEPIA數據庫生存數據,分析ZNF425在肺腺癌和肺鱗狀細胞癌組織中的表達水平對患者生存周期的影響,繪制生存曲線發現,癌癥確診后的100個月內,高表達ZNF425的患者比低表達患者生存率高,這提示ZNF425的高表達可能對癌癥有抑制作用(如圖1c)。

(A)ZNF425在NSCLC中表達下調;(B)利用GEPIA工具繪出的肺癌病理分期不同階段ZNF425的表達圖譜;(C)采用GEPIA工具分析TCGA數據庫中ZNF425表達水平的Kaplan-Meier總體生存(OS)曲線(n(low)=479, n(high)=480)圖1 生物信息學分析(來源于TCGA和GEPIA數據庫)Fig. 1 Bioinformatics analysis from TCGA and GEPIA database
2.2 ZNF425在人肺癌組織中的表達分析
首先對不同分型的病理標本進行石蠟包埋,切片。然后利用ZNF425特異性抗體進行免疫組化,將癌旁組織作為對照,分析癌癥組織較癌旁組織中ZNF425的蛋白水平變化情況。發現癌癥組織相比癌旁組織染色區域面積減小,染色程度也相對較淡(圖2a)。最后使用Image J軟件進行定量分析,發現ZNF425表達水平在3個時期都有所下調,且III期尤為顯著(圖2b)。這提示早中期肺癌的發生抑制了ZNF425的表達。

(a)不同病理分型的癌癥組織及癌旁組織的ZNF425免疫組化(IHC)檢測圖(比例尺:200 μm);(b)定量分析,**P<0.01,ns無顯著性。圖2 ZNF425免疫組化(IHC)及量化圖Fig. 2 ZNF425 immunohistochemical and quantitative images
2.3 ZNF425在肺癌中的功能與MAPK信號通路有關
上述結果說明ZNF425與肺癌早中期的發生具有相關性。本文利用廣泛用于非小細胞肺癌的細胞水平研究的A549腺癌人類肺泡基底上皮細胞系,在其轉入pCMV-ZNF425-Tag2B質粒和pCMV-Tag2B空載,48 h后收集細胞進行WB實驗檢測MAPK信號通路中關鍵調節因子JNK和P38的蛋白水平和磷酸化水平變化。結果顯示在A549細胞中過表達ZNF425后,與空載相比,JNK和P38的蛋白水平變化不明顯,而磷酸化水平都出現了下調的情況(圖3)。
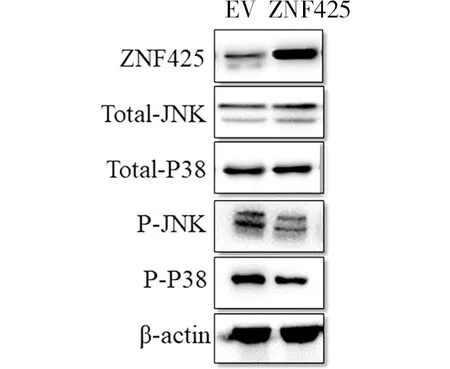
圖3 過表達ZNF425影響MAPK信號通路Fig. 3 Overexpression of ZNF425 affected MAPK
3 討論
KRAB-ZNF家族是高度復雜的,因為它包含了大量的同源基因、基因亞型和假基因,如此巨大的多樣性阻礙了對這些蛋白質的詳細分子特征分析。現在越來越多的研究報道表明KRAB-ZNF涉及了腫瘤生物學的多個方面。有學者利用TCGA項目中LUAD和LUSC數據集分析KRAB-ZNF基因表達模式與臨床參數之間的關系,發現KRAB-ZNF基因的表達大多數具有顯著差異性。且這些差異與肺癌的組織學分型、分子亞型和患者生存率有關[17]。這與本文的初步結果相符合:TCGA數據顯示,ZNF425的表達在兩種肺癌早中期都有所下調。同時數據顯示,在癌癥確診后的100個月內,高表達ZNF425的患者比低表達患者生存率更高。這意味著ZNF425在肺癌的發展中具有一定功能,而且可能對肺癌早中期的發展起到抑制作用。對人類肺癌病理樣本和癌旁組織樣本經過ZNF425的免疫組化和定量分析后發現:與癌旁組織相比,Ⅰ、Ⅱ、Ⅲ期的肺癌組織中ZNF425蛋白水平都有下調的趨勢,其中第Ⅲ期下調最為明顯。這印證了生物信息學的結論。
在前期研究中,已經證明ZNF425可以抑制SRE,AP-1和SRF的轉錄活性,同時在白血病Jurkat細胞系中瞬時過表達ZNF425,使ERK活性下降并對Jurkat細胞產生影響。這些結論都表明ZNF425與MAPK信號通路緊密相關。同時MAPK信號通路在癌癥的發生和治療上有著舉足輕重的地位。其中最著名的是RAS-MAPK信號軸(也稱為RAS-RAF-MEK-ERK途徑),它在約40%的人類癌癥中發生了變化,主要是由BRAF(約10%)及其上游激活因子RAS(約30%)的突變引起的,且上游調節子和下游效應子的突變也很普遍[18]。其次是JNK途徑,有研究表明JNK途徑常常在各種白血病和實體腫瘤包括皮膚、肺、結腸、腦及肝癌中過度活化[19,20],這提示JNK是腫瘤啟動子,也有研究證明JNK能增強癌癥對ERK途徑抑制劑和化學治療藥物的抵抗力[21]。最后是P38途徑,活性氧(ROS)的產生能夠激活P38,誘導癌細胞的死亡,但這僅限于癌癥的起始階段,一旦腫瘤形成,p38活性將支持其生長、轉移和抵抗藥物[22,23]。因此,本文在A549細胞系中瞬時過表達ZNF425后,通過WB檢測MAPK信號通路中的JNK和P38這兩個關鍵調節因子的磷酸化水平,發現這兩個因子的活性都出現了不同程度的下調。由此可以確認,ZNF425蛋白在早中期肺癌的發展中具有一定功能,且很可能是通過抑制JNK和P38的過度活化從而抑制肺癌的生長、轉移和對藥物的抵抗。
隨著對KRAB-ZNF蛋白研究的深入,發現其在生物體內具有重要作用。本文首次探究了ZNF425在肺癌中的功能,粗略地推測ZNF425蛋白可能具有通過抑制JNK和P38的過度活化從而抑制肺癌在早中期的生長、轉移和對藥物的抵抗的功能。其具體的作用機制還需進一步從臨床收集更多的病理樣本并進行藥物耐受實驗。相信對ZNF425的更深入了解能夠轉化為更好的肺癌治療方案。
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
財經(2017年2期)2017-03-10 14:35:35
山東工業技術(2016年15期)2016-12-01 05:31:22
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
財經(2016年6期)2016-02-24 07:41:51
醫學研究雜志(2015年12期)2015-06-10 06:57:46
