circ-PTPN22在系統性紅斑狼瘡患者外周血單個核細胞中的翻譯及意義
2020-08-18 10:00:34仲志婷苗青青蔣柱燕張夢潔唐雋
中華皮膚科雜志 2020年7期
關鍵詞:檢測
仲志婷 苗青青 蔣柱燕 張夢潔 唐雋
1安徽醫科大學附屬解放軍聯勤保障部隊第901 醫院皮膚科,合肥230031;2陸軍軍醫大學病理生理與高原病理學教研室,重慶400038
環狀RNA(circRNA)是一種非編碼RNA分子,沒有5′端帽和3′端poly(A)尾部,通過共價鍵形成環狀結構[1]。circRNA 可作為miRNA 的“海綿”來調節下游靶基因的表達[2],調節親本基因轉錄[3],與RNA 結合蛋白相互作用[4],以及翻譯蛋白質[5]。我們前期研究[6]采用高通量RNA測序,分析了3例系統性紅斑狼瘡(SLE)患者與3例健康對照外周血單個核細胞(PBMC)中circRNA 的差異表達,通過生物信息學分析篩選出4 個候選circRNA,并經實時熒光定量PCR(qRT-PCR)證實circ蛋白酪氨酸磷酸酶非受體22型(PTPN22)在SLE患者PBMC中表達降低,且與SLE 疾病活動指數(SLEDAI)評分呈負相關。本研究探討circ-PTPN22的翻譯能力及其翻譯蛋白對Jurkat細胞活化和凋亡的作用。
對象與方法
一、研究對象
2019 年5 月10 日至9 月30 日分別于西南醫院中西醫結合風濕免疫科和體檢中心收集6 例SLE女性患者(SLEDAI評分10 ~14分)和9例健康女性對照全血樣本,年齡25 ~50 歲。本研究經解放軍第105 醫院(現更名為解放軍聯勤保障部隊第901醫院)倫理委員會批準( 倫理編號105LL20150308),所有受試者均簽署知情同意書。
二、細胞來源及主要試劑
Jurkat 細胞系來源于美國ATCC 細胞庫。RiboTM熒光原位雜交試劑盒(廣州銳博生物技術有限公司),circ-PTPN22-cy3探針和18S-cy3對照探針均由廣州銳博生物技術有限公司設計合成。內參甘油醛-3-磷酸脫氫酶(GAPDH)單抗(美國Bioss公司),山羊抗兔抗體、山羊抗小鼠抗體(北京中杉金橋生物技術有限公司),circ-PTPN22-FLAG 和circ-PTPN22-NC-FLAG 重組慢病毒由上海生博生物醫藥科技有限公司構建,circ-PTPN22-shRNAFLAG和circ-PTPN22-shRNA-NC-FLAG慢病毒由上海泰兒圖生物科技有限公司合成。Dynabeads?Human T-Activator CD3/CD28 用于活化和擴增人T細胞(美國Thermo Fisher Scientific 公司)。藻紅蛋白標記的抗人白細胞介素(IL)-2 抗體、7-AAD Viability Staining Solution(美國Biolegend 公司),小鼠抗FLAG?M2 單抗(美國Sigma-Aldrich 公司),ELISA 試劑盒(英國abcam 公司),RIPA 裂解液、一抗稀釋液、二抗稀釋液、Quick-Blocker 封閉液、2×十二烷基硫酸鈉聚丙烯酰胺凝膠(SDS-PAGE)蛋白上樣緩沖液(上海碧云天生物技術有限公司),ExpressPlus?PAGE Gels 4%~20%(南京金斯瑞生物科技有限公司),EasySep?人T細胞分離試劑盒、EasySep?人B 細胞分離試劑盒、EasySep?人NK 細胞分離試劑盒(加拿大STEMCELL Technologies 公司),eBioscience?Foxp3 轉錄因子染色液(美國Thermo Fisher Scientific公司)。
三、方法
1.PBMC分離:全血與磷酸鹽緩沖液(PBS)及樣本密度分離液體積比為1∶1∶1,805×g離心20 min,取白膜層,加PBS 10 ml,201×g 離心5 min 后棄上清液,得到PBMC。加入二甲基亞砜和胎牛血清(體積比1∶9),-80 ℃過夜,第2天轉入液氮中保存。
2.熒光原位雜交法觀察circ-PTPN22在健康人PBMC中的表達定位:針對circ-PTPN22剪接位點設計一段帶有cy3 標記的探針circ-PTPN22-cy3,對照組探針為18S-cy3。從3 例健康人5 ml 全血樣本中分離PBMC。50 μl PBS 重懸PBMC,取2 張多聚賴氨酸處理過的載玻片,分別標記為circ-PTPN22-cy3組和18S-cy3組。各取10 μl細胞懸液到載玻片,靜置待干。然后依次用4%多聚甲醛室溫固定10 min,預冷通透液4 ℃5 min,200 μl預雜交液37 ℃30 min,避光下將circ-PTPN22-cy3 和18S-cy3 兩種探針各1.5 μl 分別加入100 μl 預熱的雜交液混勻,棄去載玻片上的預雜交液,將探針混合物滴到含有細胞的載玻片上,放入濕盒,37 ℃避光過夜。第2天,42 ℃避光條件下,分別用雜交洗液Ⅰ、雜交洗液Ⅱ、雜交洗液Ⅲ洗滌,然后用4′,6-二脒基-2-苯基吲哚(DAPI)染色10 min,最后熒光顯微鏡下拍照。
3. Jurkat 細胞培養與慢病毒轉染:用RPMI 1640培養基(含10%胎牛血清、1%青霉素鏈霉素)、在37 ℃5% CO2恒溫孵箱中培養Jurkat 細胞,第2 天換液。為驗證預測的circ-PTPN22 蛋白編碼功能,分別將circ-PTPN22-FLAG、circ-PTPN22-NCFLAG、circ-PTPN22-FLAG + circ-PTPN22-shRNAFLAG、circ-PTPN22-shRNA-NC-FLAG 重組慢病毒[病毒載體含綠色熒光蛋白(GFP)基因]轉入Jurkat細胞(circ-PTPN22-FLAG 組、circ-PTPN22-NCFLAG 組、circ-PTPN22-shRNA-FLAG 組、circ-PTPN22-shRNA-NC-FLAG 組),同時設置Jurkat 細胞對照組(僅正常培養)。采用12 孔板,每孔5 ×105個細胞,每組各3 個復孔,重組慢病毒感染復數(MOI)均為50。培養48 h后收集細胞,期間可根據細胞生長狀態添加完全培養基和傳代。
4.circ-PTPN22pro 抗體制備與鑒定:委托上海烈冰生物醫藥科技有限公司測定circ-PTPN22核酸序列。根據測序結果,委托杭州華安生物技術有限公司[動物試驗許可證號:SYXK(浙)2017-0009]針對circ-PTPN22 剪接位點處的特異性氨基酸序列GNSNYLPVSLQELARP 合成肽段并制備多克隆抗體,然后使用親和層析柱純化抗體。采用常規多克隆抗體制備方法,選用2 只體重2.5 kg 左右的健康新西蘭雄性白兔(編號為4137、4138),采用多點皮下注射各免疫3次,首次免疫后第14天進行2次免疫,第2 與第3 次免疫間隔7 d,第3 次免疫后第7 天,于兔耳中動脈采少量全血提取血清進行間接ELISA檢測。檢測合格后,進行加強免疫,7 d后可采集全血。采用轉染了circ-PTPN22 的Jurkat 細胞鑒定所制備的多克隆抗體,將轉染的Jurkat細胞裂解上清液經SDS-PAGE電泳后,用所制備兔抗血清作為一抗進行Western印跡法分析。
5. 間接ELISA 法檢測制備的兔抗circ-PTPN22pro 抗體效價:按ELISA 試劑盒說明書依次進行包被抗原(合成的目的肽段)、1×TBST緩沖液洗滌、TBST配制的1%牛血清白蛋白(BSA)封閉,加入稀釋的待檢樣品(即制備的抗體)37 ℃反應1 h、洗滌2 次。設置陽性對照孔(陽性血清)與陰性對照孔(BSA)。然后加辣根過氧化物酶標記二抗羊抗兔抗體孵育,1×TBST 緩沖液洗3次。加底物顯色,2 mol/L硫酸終止反應后,于酶標儀450 nm波長處讀取吸光度。各實驗孔與BSA陰性對照孔A450比值>2.1時,為抗體滴度判定終點。
6.磁珠分選人PBMC:根據EasySep?Human T Cell Isolation Kit,分別用250 μl EasySep?緩沖液重懸3 例健康對照和3 例SLE 患者各15 ml 全血提取的PBMC,并用細胞計數板計數,每管加入50 μl/ml Isolation Cocktail,混勻后室溫等待5 min,振蕩30 s后每管加入40 μl/ml RapidSpheres?,用EasySep?緩沖液補齊至2.5 ml,混勻后放入配套的磁鐵中,室溫靜置3 min,垂直將細胞懸液倒入另一新流式管中,將新管放入磁鐵中,靜置3 min,同樣的方法倒出,得到分選后的T 細胞。參照上述方法分選B、NK細胞。用相應的抗體檢測細胞純度。
7. Western 印跡法檢測細胞中circ-PTPN22pro蛋白表達:按照“3.Jurkat 細胞培養與慢病毒轉染”中方法轉染48 h 后,收集各組Jurkat 細胞,RIPA 裂解液提取總蛋白。加入2 × SDS 蛋白上樣緩沖液(1∶1),煮沸變性。每孔上樣量30 μg,160 V恒壓電泳35 min,25 V 25 mA 6 min 半干轉到聚偏二氟乙烯膜上,室溫封閉30 min,分別加一抗小鼠抗FLAG標簽抗體(1∶1 000)、兔抗人GAPDH抗體(1∶4 000)4 ℃孵育過夜。TBST 洗膜5 次,每次6 min;加二抗山羊抗兔、山羊抗鼠抗體(均為1∶4 000)室溫搖晃孵育1 h,TBST 洗膜3 次,每次10 min,然后用Bio-Rad 成像儀曝光拍照。使用Image Lab 6.0 軟件計算每組circ-PTPN22pro 條帶與GAPDH 條帶的調整灰度值之比。此外,采用上述方法檢測3例健康對照和3 例SLE 患者PBMC 及另3 例健康對照和3 例SLE 患者T、B、NK 細胞亞群中circ-PTPN22pro 表達,一抗為制備的兔抗人circ-PTPN22pro 抗體(1∶500),二抗為山羊抗兔抗體。
8. 流式細胞儀檢測circ-PTPN22 對細胞活化和凋亡的影響:按照“3.Jurkat 細胞培養與慢病毒轉染”中方法將circ -PTPN22、circ-PTPN22-NC、circ-PTPN22 - shRNA、circ - PTPN22 -shRNA-NC 慢病毒分別轉染Jurkat 細胞(circ-PTPN22 組、circ-PTPN22-NC 組、circ-PTPN22-shRNA 組、circ-PTPN22-shRNANC 組)24、48 和72 h 后,每孔加Dynabeads ?Human T-Activator CD3/CD28 4 μl,恒溫孵育6 h 后收集轉染的Jurkat 細胞。分別用200 μl PBS重懸,各組取1×106細胞,加入200 μl穿孔液室溫20 min,500 μl 通透緩沖液453 × g 離心8 min,洗滌2次。重懸后每管加入1 μl藻紅蛋白抗人IL-2抗體,室溫避光20 min后洗滌2次,用含2%胎牛血清的PBS重懸細胞,流式細胞儀檢測IL-2陽性細胞比例,以檢測細胞活化狀態。此外,收集各組轉染48、72、96 h后的Jurkat 細胞,檢測前10 min每管加7-AAD Viability Staining Solution 2 μl,流式細胞儀檢測細胞凋亡率,即7-AAD陽性細胞比例。
9.統計學方法:應用SPSS25.0 軟件,采用兩因素重復測量方差分析分別分析過表達和干擾circ-PTPN22 不同時間對Jurkat 細胞活化及凋亡的影響。采用GraphPad Prism8 軟件,兩組計量資料的比較采用兩獨立樣本t檢驗,P<0.05認為差異有統計學意義。
結 果
一、circ-PTPN22在健康人PBMC細胞中的定位
熒光原位雜交法顯示,circ-PTPN22 和陽性對照18S 均主要表達在健康人PBMC 細胞質內,見圖1。
二、circ-PTPN22的翻譯能力預測
測序結果顯示,circ-PTPN22基因全長1 429 bp,含9 個外顯子。為預測該基因是否具有翻譯蛋白的能力,通過circRNADb 網站(http://reprod.njmu.edu.cn/circrnadb)進行比對,預測出circ-PTPN22 第718 ~838位堿基共121 bp的內部核糖體進入位點(IRES)序列(圖2A 綠色序列)。因為測得的circ-PTPN22全長為1 429 bp,所以排除1 399 ~1 507 bp的潛在IRES 序列。翻譯能力預測見圖2B,ORF 框從910 bp 跨剪接位點至50 bp 處,潛在翻譯的蛋白質序列共189 個氨基酸,將其命名為circ-PTPN22pro;GNSNYLPVSLQELARP 序列為該編碼蛋白不同于全長PTPN22 蛋白的特異氨基酸序列(圖2B)。

圖1 熒光原位雜交法檢測circ-PTPN22在健康人外周血單個核細胞中的定位 圖像融合后可見circ-PTPN22與18S均主要表達在細胞質中

圖2 circ-PTPN22蛋白編碼預測 2A:circ-PTPN22基因的預測編碼序列;綠色部分代表IRES序列,灰色部分代表ORF框,跨剪接位點翻譯,紅色標記ATG、TGA分別為翻譯的起始密碼子和終止密碼子;2B:circ-PTPN22pro的預測氨基酸序列,序列的紅色部分是不同于全長PTPN-22蛋白的獨特氨基酸序列
三、circ-PTPN22pro多克隆抗體制備及鑒定
ELISA 結果顯示,3 次免疫后,2 只兔血清效價均達到1∶256 000。Western 印跡法鑒定circ-PTPN22pro 多克隆抗體顯示,4 137 號兔血清的特異性較好,相對分子質量為20 000左右的目的條帶顯示清晰,但未檢測到全長型PTPN22pro(相對分子質量98 000)(圖3),提示circ-PTPN22pro 多克隆抗體制備成功,特異性針對circ-PTPN22pro。因此,在后續臨床相關實驗中使用4 137 號兔血清進行實驗。
四、circ-PTPN22編碼功能驗證
轉染重組慢病毒后48 h,circ-PTPN22-FLAG組、circ-PTPN22-NC-FLAG組、circ-PTPN22-shRNAFLAG 組、circ-PTPN22-shRNA-NC-FLAG 組細胞均有綠色熒光表達(圖4A),說明重組慢病毒轉染成功。Western 印跡結果顯示,circ-PTPN22-FLAG 組circ-PTPN22pro 表達明顯高于circ-PTPN22-NCFLAG 組和細胞對照組,且相對分子質量約為20 000;circ-PTPN22-NC-FLAG 組及細胞對照組有少量表達,提示Jurkat 細胞內源性表達circ-PTPN22pro(圖4B)。與circ-PTPN22-FLAG組相比,circ-PTPN22-shRNA-FLAG 組circ-PTPN22pro 表 達明顯降低,見圖4C。
五、circ-PTPN22對細胞活化和凋亡的影響
轉染24、48、72 h后,流式細胞儀檢測顯示,circ-PTPN22 組IL-2 表達(22.20% ± 8.92%,31.10% ±5.88%,53.20% ± 10.25%)均低于circ-PTPN22-NC組(30.90% ± 11.00%,51.23% ± 10.70%,69.67% ±9.00%);circ-PTPN22-shRNA組IL-2表達(35.57%±8.79%,78.10%± 10.08%,88.63%± 3.89%)均高于circ-PTPN22-shRNA-NC 組(26.73% ± 4.92%,41.03% ± 10.45%,41.33% ± 4.96%),見圖5A、5B。兩因素重復測量方差分析顯示,circ-PTPN22 組與circ-PTPN22-NC 組間差異有統計學意義(F =284.881,P=0.003);circ-PTPN22-shRNA 組與circ-PTPN22-shRNA-NC 組間差異亦有統計學意義(F=293.818,P=0.003)。

圖5 流式細胞儀檢測circ-PTPN22 對Jurkat 細胞活化和凋亡的影響 轉染24、48 和72 h 后,circ-PTPN22 組IL-2 表達均明顯低于circ-PTPN22-NC組(5A),circ-PTPN22-shRNA組IL-2表達均明顯高于circ-PTPN22-shRNA-NC組(5B);轉染48、72、96 h后,circ-PTPN22組細胞凋亡率均高于circ-PTPN22-NC組(5C),circ-PTPN22-shRNA組細胞凋亡率亦明顯高于circ-PTPN22-shRNA-NC組(5D)
轉染48、72、96 h后,流式細胞儀檢測顯示,circ-PTPN22 組細胞凋亡率(7.08% ± 1.33%,19.17% ±3.29%,30.03% ± 9.24%)高于circ-PTPN22-NC 組(5.01% ± 1.52% ,5.63% ± 0.81% ,14.22% ±3.96%),且circ-PTPN22-shRNA 組細胞凋亡率(24.63% ± 10.58%,59.17% ± 15.64%,84.87% ±2.91%)亦明顯高于circ-PTPN22-shRNA-NC 組(6.59% ± 3.37% ,5.33% ± 1.65% ,15.42% ±7.31%),見圖5C、5D。兩因素重復測量方差分析顯示,circ-PTPN22 組與circ-PTPN22-NC 組間差異有統計學意義(F = 81.287,P = 0.012);circ-PTPN22-shRNA 組與circ-PTPN22-shRNA-NC 組間差異亦有統計學意義(F=111.813,P=0.009)。
六、circ-PTPN22pro 在 健 康 對 照 和SLE 患 者PBMC中的差異表達
Western 印跡結果顯示,SLE 組PBMC 中circ-PTPN22pro 表達低于健康對照組,幾乎不表達(圖6A)。進一步用磁珠分選后結果顯示,SLE 組T、B細胞中circ-PTPN22pro 表達(0.398±0.147、0.072±0.027)均顯著低于健康對照組(1.090 ± 0.365、0.512 ± 0.157;t 值分別為3.047、4.806;P 值分別為0.038、0.009),SLE組與健康對照組NK細胞中circ-PTPN22pro 表達差異無統計學意義(0.139 ± 0.147比0.088±0.032;t=0.582,P=0.592),見圖6B。
討 論
一些有IRES 序列及ORF 框的circRNA 具有編碼蛋白能力[7]。我們在circRNADb數據庫比對circ-PTPN22序列,發現circ-PTPN22有跨剪接位點翻譯的ORF 框,并在Jurkat 細胞中檢測出circ -PTPN22pro表達。通過熒光原位雜交法,我們檢測出circ-PTPN22 主要在細胞質中表達,提示circ-PTPN22 可能主要在細胞質中發揮作用。與circ-PTPN22-NC 組相比,過表達circ-PTPN22 可抑制Jurkat 細胞活化,促進Jurkat 細胞凋亡;而干擾了circ-PTPN22 的表達可促進細胞活化,同時促進細胞凋亡。推測細胞凋亡的原因可能是干擾circ-PTPN22 表達后,circ-PTPN22pro 蛋白表達降低,導致細胞過度活化誘導了細胞凋亡[8]。
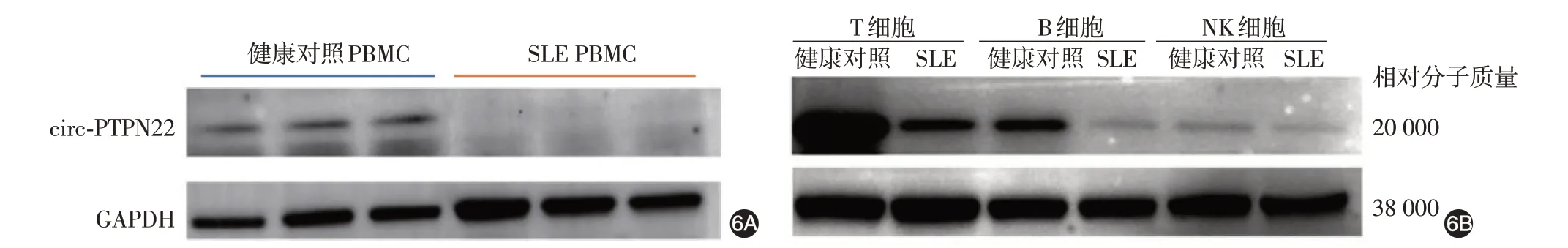
圖6 系統性紅斑狼瘡(SLE)患者外周血單個核細胞(PBMC)及其T、B、NK 細胞亞群中circ-PTPN22 蛋白表達 6A:PBMC 中circ-PTPN22pro表達 3例SLE患者均明顯低于3例健康對照;6B:T、B細胞中circ-PTPN22pro表達 SLE患者均明顯低于健康對照,但兩組NK細胞中circ-PTPN22pro表達無明顯差異
通過制備兔抗circ-PTPN22pro 特異性多克隆抗體,我們檢測了SLE患者和健康對照PBMC及其T、B、NK細胞亞群中circ-PTPN22pro的表達。結果顯示,SLE 患者組PBMC 中circ-PTPN22pro 表達明顯低于健康對照組,甚至不表達;SLE 患者組T、B細胞亞群中circ-PTPN22pro 表達亦顯著低于健康對照組,而兩組NK 細胞中circ-PTPN22pro 表達無明顯差異。提示circ-PTPN22 和circ-PTPN22pro 蛋白可能對SLE的發生發展有一定作用,是影響SLE疾病的重要靶標分子之一。
PTPN22是一種淋巴特異性磷酸酶(Lyp),可抑制T細胞活化;當PTPN22第620位精氨酸(R)突變成色氨酸(W)時,PTPN22活性增強,可抑制T 細胞受體(TCR)的信號傳導,從而促進自身免疫的發展[9]。PTPN22突變體PTPN22-R620W(LypW)在多種自身免疫疾病中起重要作用,比如SLE[10]、類風濕關節炎[11]等。SLE患者血清干擾素(IFN)-α水平升高,且IFN 驅動的下游基因表達增高[12]。然而,攜帶PTPN22 突變體LypW 的SLE 患者與非攜帶SLE患者血漿IFN-α水平無明顯差異,但攜帶LypW的患者產生Toll 樣受體(TLR)7 誘導的Ⅰ型IFN 能力降低,提示攜帶LypW 的SLE 患者可能存在Toll樣受體或漿細胞樣樹突細胞依賴的宿主防御或抗炎功能缺陷[13]。PTPN22 是circ-PTPN22 的親本基因,推測circ-PTPN22 可能與其親本基因之間有相互作用或者具有類似PTPN22的作用。在后續研究中,我們將對circ-PTPN22與PTPN22相互作用進行深入研究,探索circ-PTPN22的作用機制,研究circ-PTPN22 或circ-PTPN22pro 蛋白在SLE 中的作用及機制。
志謝陸軍軍醫大學免疫學教研室楊玓老師和田志強老師;西南醫院中西醫結合風濕免疫科和體檢中心
利益沖突所有作者均聲明不存在利益沖突
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
