中亞高海拔梯度山盆系統草地凈初級生產力時空演變格局及其驅動機制分析
2020-08-20 09:06:58畢旭徐向超張立新
生態環境學報 2020年5期
畢旭 ,徐向超,張立新
1. 山西財經大學資源環境學院,山西 太原 030006;2. 北京師范大學地理科學學部,北京 100875;3. 山西省科學技術情報研究所,山西 太原 030024;4. 北京大學城市與環境學院,北京 100871
草地生態系統是陸地生態系統的重要組成部分,幾乎占據了陸地面積的30%(Houghton,1994),在全球碳循環中發揮著重要作用(Ahlstr?m et al.,2015)。除此之外,草地生態系統在涵養水源及維持生物多樣性穩定等生態服務功能和初級生產及保障畜牧業可持續發展等社會生產功能方面起著重要的作用(鄭曉翾等,2007;賈文雄等,2015)。陸地生態系統在過去幾十年經歷了嚴峻的變化,氣候變化和人為活動共同導致了陸地生態系統的動態變化(Haberl,1997)。目前,人類可在一定程度上準確的利用遙感對陸地生態系統動態進行監測,但目前對氣候變化和人類活動對草地生態系統變化的影響還沒有統一的結論(Fensholt et al.,2012)。特別是在半干旱和干旱地區,氣候變化和人類活動的加劇可能導致生態系統退化和嚴重的生態和經濟損失(Wessels et al.,2008)。中國草地面積為393×104km2,退化草地面積約 86.67×104km2。近年來的研究表明,中國北方地區約90%的草地已發生了一定程度的退化(Nan,2005)。因此,準確評價和區分人為活動和氣候變化對生態系統變化的相對影響,了解草地退化和恢復的主要原因,對于制定有效的生態管理和調控政策具有重要意義。
山盆系統(MBS)是中亞干旱區典型的地貌結構(Zhang,2001),是由山地和盆地組合而成的一種地貌組合(圖1、2)。草地是該區域的主要土地利用覆被類型;因此,草原生態系統的可持續性利用對社會穩定和區域生態安全起著至關重要的作用。干旱地區草地生態系統由于土壤貧瘠、植被稀少等原因,對人類活動和氣候變化更為敏感(Seddon et al.,2016)。一些研究表明中亞地區的草地在氣候變化和頻繁的人類活動的影響下經歷了退化(Zhang et al.,2017)。以往的研究表明在降水量增加,全球氣溫變暖及生態保護工程實施的影響下,草地 NPP 的變化呈增加趨勢(Zhou et al.,2015)。除此之外,一些研究表明草地NPP的下降是過度放牧和干旱氣候影響的結果(Wang et al.,2016)。由于北疆地區山盆系統地形較為復雜,且跨越較大的海拔梯度,使得氣候變化和人類活動對不同草地類型的影響顯示出了很強的空間異質性(Cudahy et al.,2016)。近些年,關于氣候變化和人類活動對植被多樣性的影響日漸增多,但是主要集中在流域(Wang et al.,2015;Xu et al.,2017),區域(Xu et al.,2016;Liu et al.,2018)和全球尺度(Thomas et al.,2014;Liu et al.,2015)。然而,對于高海拔梯度差的干旱區山盆系統的研究相對較少。
植被退化的一個重要表現是植被生產力的下降,而植被凈初級生產力(NPP)(Matsushita et al.,2002)是表征生產力狀況的一項重要指標,是評價陸地生態系統質量狀況的重要因子,且是地表碳循環的重要組成部分(Liang et al.,2015)。氣候變化和人類活動所引起的草地恢復和退化會直觀表現為草地NPP的變化,因此,NPP已被廣泛用于監測氣候變化與生態系統的人為改變的生態效益的評價指標(Ma et al.,2012)。為了更好地理解人類活動對生態系統的影響,Haberl提出了凈初級生產(NPPH)的人為占用(Haberl,1997)這一概念。一些學者已經應用NPPH來估計人類活動對生態系統的作用(Guo et al.,2017;Zhou et al.,2017)。該方法為定量分析氣候變化和人類活動對草地 NPP變化的相對影響提供了有效途徑。
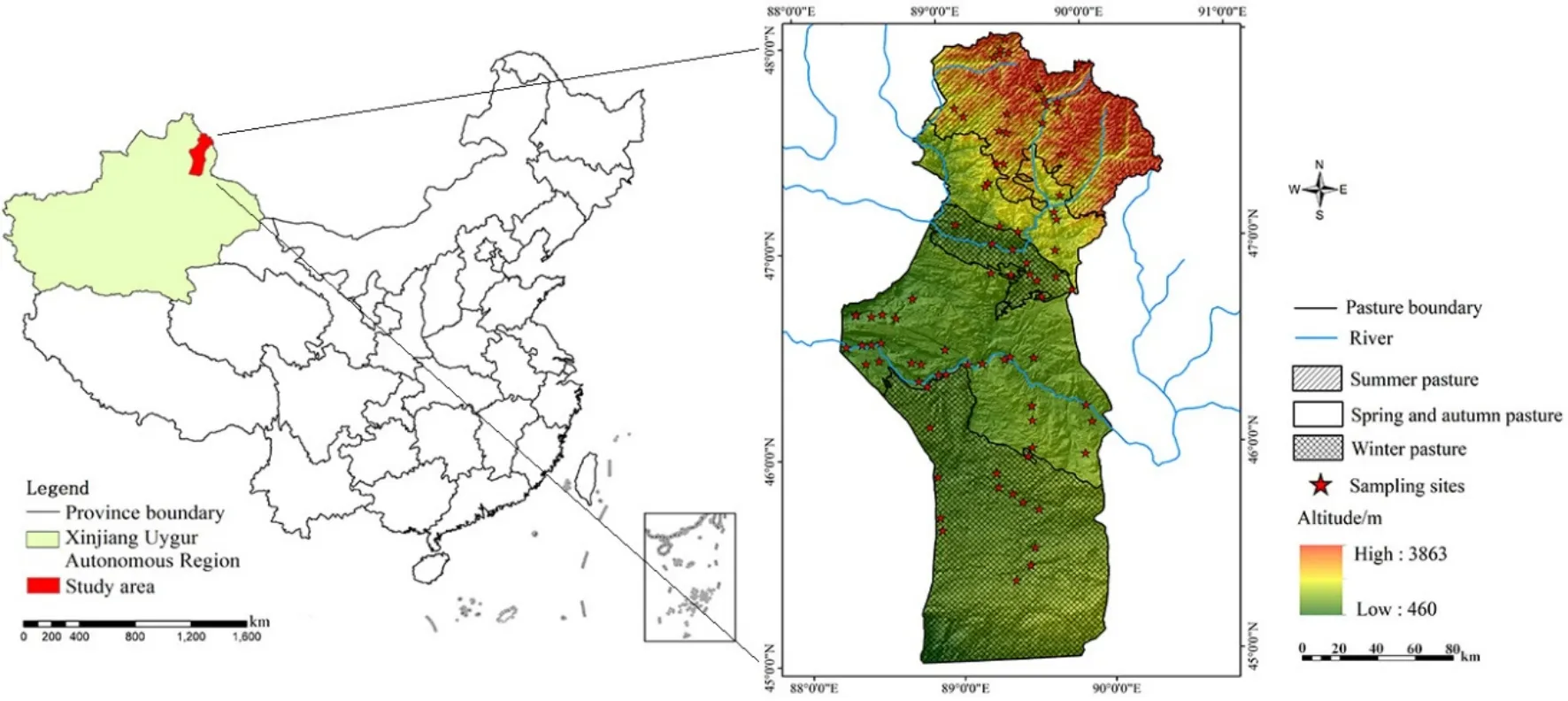
圖1 研究區地理位置圖Fig. 1 Map of geographical location of the study area
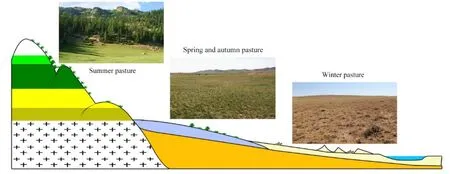
圖2 山盆系統景觀剖面和不同季節牧場景觀照片Fig. 2 Landscape profiles of mountain basin systems and photographs of pasture landscapes in different seasons
本文利用多源遙感數據,以北疆阿勒泰地區富蘊縣為例,在模擬草地實際NPP和潛在NPP的基礎上,進行了以下研究:(1)探索富蘊縣2001—2015年草地NPP的空間分布格局和動態;(2)分析草地NPP變化中氣候變化與人為活動的相對貢獻;(3)研究不同海拔梯度和不同季節牧場草地 NPP與降水和氣溫的相關性及影響其變化的主要因素。該研究有助于中亞干旱地區山盆系統的草地資源管理、草地可持續發展和草地退化的防治。
1 材料與方法
1.1 研究區概況
富蘊縣地處新疆維吾爾自治區最北部的阿勒泰地區的東面,位于 45°00′—48°03′N,88°10′—90°31′E之間,北邊與蒙古國接壤。東臨清河縣,西接福海縣,南延準噶爾盆地。行政區域面積3.37×104km2,海拔范圍為317—3863 m,是中亞地區具有代表性的山盆系統,富蘊縣的海拔梯度使得氣候和植被的垂直分布特征明顯。研究區位于溫帶干旱半干旱氣候區,年平均氣溫為4.60 ℃(2007—2016),年平均降水量為208.41 mm(2007—2016)。境內河流主要有兩條,即額爾齊斯河和烏倫古河。草地是該區域的主要生態系統類型,總面積26656.33 km2,占區域總面積的82.82%。研究區草地資源采用中國草地資源分類系統進行分類(Ren et al.,2008),劃分為 10個草地類。牧民和牲畜根據季節在不同海拔的草原上遷徙,根據牧民對草原的利用時間,將草原分為不同季節的草場(夏牧場、春秋牧場、冬牧場)(圖1)。夏牧場分布在海拔1200 m以上阿爾泰山區,內含高寒草甸、山地草甸、高寒草原以及少部分的溫性草甸草原。牧草群落多為豆科和草本科牧草及部分雜草類,草質優良。放牧始于6月上旬末終于9月上旬末,約90 d。冬牧場絕大部分是低劣草場,主要由溫性荒漠構成。分布于海拔 500 m以下,多系荒漠草原,牧草群落多為旱生小灌木為主,草質較差。放牧期始于12月初,終于次年3月下旬,約 120 d。春秋牧場分布于冬、夏牧場之間,主要在海拔500—1200 m的丘陵和平原荒漠地區,牧草群落以旱生禾草為主,還有一些小灌木和雜草。放牧時間分兩季:春季始于3月下旬,終于6月上旬末,利用時間約70 d;秋季放牧始于9月上旬末,終于12月初,利用時間約85 d。春秋牧場主要由部分溫性草甸草原、溫性典型草原、溫性草原化荒漠、溫性荒漠草原以及溫性荒漠構成。
1.2 數據來源
本研究所用氣象數據來源于中國氣象科學數據共享服務網(http://data.cma.cn/),共收集到新疆維吾爾族自治區54個國家級氣象站點2001—2015年的氣溫、降水數據,以及11個站點2001—2015年的太陽輻射數據。本研究所用MODIS NDVI數據源于美國國家航空航天局(ftp://ladsweb.nascom.nasa.gov),2001—2015年MOD13Q1數據集,空間分辨率為250 m,時間分辨率為16 d。利用MRT軟件進行數據拼接及投影轉換,同時采用最大化合成法(MVC)得到每月NDVI數據。利用ANUSPLIN軟件,對新疆區域的氣溫、降水和太陽輻射進行插值,之后提取出富蘊縣區域。
由于實測NPP的獲取難度較大,本研究采用生物量換算的NPP數據代替實測NPP數據進行模型驗證。生物量數據采用2015年8月野外樣方采集數據,在研究區進行草地生物量樣方調查,共設置樣地83個(圖1),每個樣地面積10 m×10 m,每個樣地內選擇草地植物群落分布較均勻的地區設置 3個樣方。在每個樣方中,收集每個草地類型的代表性物種的地上生物量部分和地下生物量部分。不同草地類型的地下與地上部分生物量比例系數與樸世龍等(2004)關于不同草地類型的地下與地上部分生物量比例系數的研究基本吻合。地上生物量的測定使用烘干法,在65 ℃的溫度下烘干至恒重后稱重。為了計算實際的 NPP,依據每種草地類型地上生物量和地下生物量之比,用地上生物量除以地上生物量和轉換為總生物量(公式1、2),比例系數參見樸世龍等(2004)。然后將總生物量乘以0.475,轉換為NPP(by C)/(g·m?2·a?1)(Scurlock et al.,1999)。
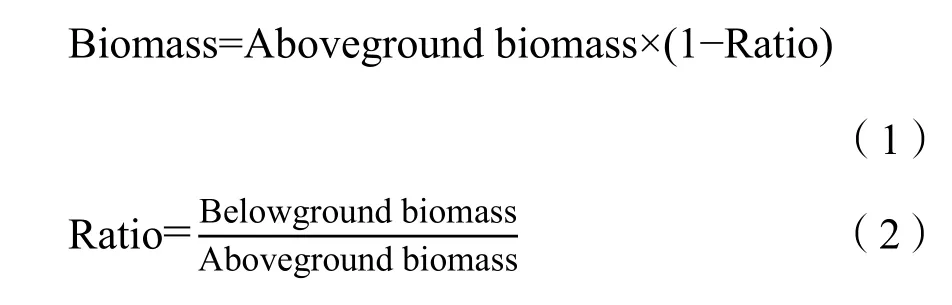
其中,Biomass指總生物量;Aboveground biomass和Belowground biomass分別代表地上生物量和地下生物量;Ratio表示地下生物量與地上生物量的比例系數。
1.3 研究方法
1.3.1 NPP的定量計算
本研究定義了3種類型的NPP,以區分受人類活動和氣候變化分別影響的草地變化。NPPA(actual NPP)是指氣候變化和人類活動共同影響下的植被實際NPP,使用CASA模型進行計算。CASA模型利用植被遙感原理,通過歸一化植被指數NDVI獲取植被對光合有效輻射的吸收系數(FPAR),再利用太陽總輻射(PAR)和FPAR計算植被吸收的光合有效輻射(APAR),進行NPP估算。基于CASA模型對植被凈初級生產力進行估算需要由APAR和光能利用率(ε)兩個變量來確定,其計算公式如下(楊紅飛等,2014):
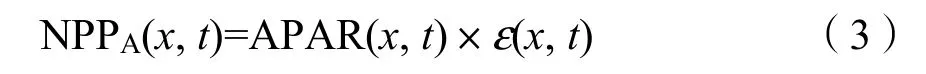
式中,APAR(x,t)和ε(x,t)分別表示像元在t月吸收的光合有效輻射和實際光能利用率。植被吸收的光合有效輻射(APAR)由太陽總輻射和植被對光合有效輻射的吸收比例計算而來,用公式(4)計算:
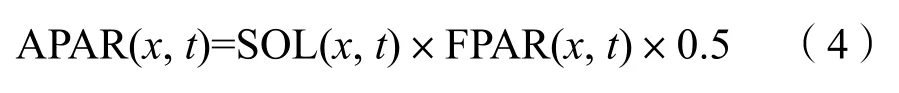
式中,SOL(x,t)表示像元在t月的太陽總輻射量,單位是MJ·m?2;常數0.5表示植被所能利用的太陽有效輻射占太陽總輻射的比例;FPAR表示植被層對入射的光合有效輻射的吸收比例。在一定范圍內FPAR與NDVI存在較好的線性關系,因而可以通過遙感影像提取NDVI對FPAR進行估算。ε是指植被把所吸收的光合有效輻射(PAR)轉化為有機碳的效率,主要受氣溫和水分兩個因子的制約,用公式(5)計算:
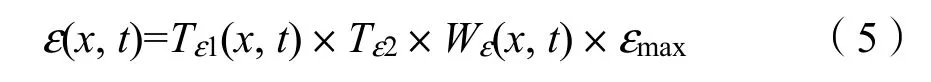
式中,Tε1(x,t)和Tε2(x,t)表示溫度對光能轉化率的影響;Wε(x,t)表示水分條件對光能轉化率的影響;εmax表示在理想狀態下植被的最大光能利用率。在本研究中參考朱文泉等(2007)利用中國植被實測數據模擬得到的最大光能利用率進行植被 NPP的估算,PFAR,Tε1(x,t),Tε2(x,t),和Wε(x,t)的計算方法及改進見文獻。
植被氣候生產力是指在無人類活動干擾且其他條件均適宜的情況下,只受氣候影響下可達到的一種理想狀態的植被凈初級生產力(周廣勝等,1996),其實質是一種潛在生產力。在本研究中用NPPP(potential NPP)來表示草地的潛在生產力,選用Thornthwaite Memorial模型來計算氣候潛在生產力,通過年均氣溫及年降水量數據,估算草地的氣候生產力NPPP。
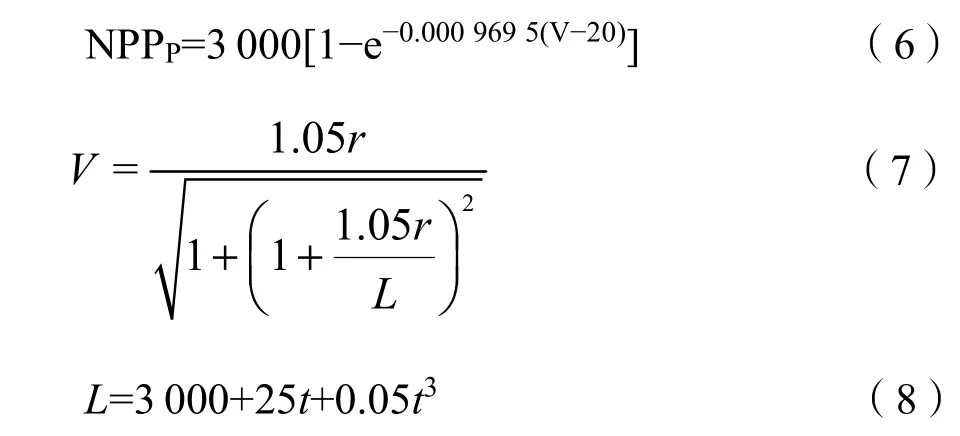
式中,NPPP(by C)/(g·m?2·a?1)為氣候潛在凈初級生產力,r為年降水量(mm);L為年最大蒸散量(mm),是年均氣溫t(℃)的函數。
第3種NPP是NPPH。采用凈初級生產力的人類占有(Human Appropriation of Net Primary Productivity,HANPP)模型法來進行氣候變化和人類活動在草地NPP變化中相對作用的定量分析(許端陽等,2011)。HANPP是指人類活動引起的草地凈初級生產力的變化,本研究中將其表示為 NPPH(Human-induced NPP),代表人類活動對草地凈初級生產力的影響,假設草地凈初級生產力的損失只是由于人類活動所引起的,計算潛在 NPPP和實際NPPA的差值得到凈初級生產力的人類占有(NPPH=NPPP?NPPA)(Haberl et al.,2002)。

通過2015年8月的草地實地觀測數據(83個數據點)驗證了CASA模型計算所得的結果的準確性。圖3為草地NPP實測值與CASA模型模擬值的一致性檢驗。統計結果(R2=0.790,P<0.001)表明研究區CASA模型是可靠的。
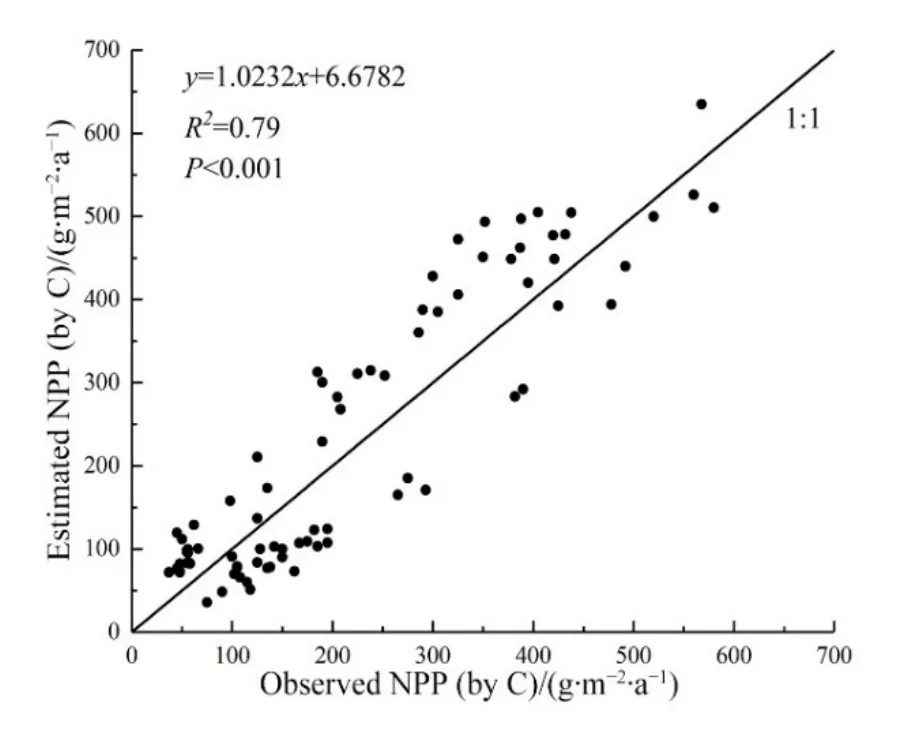
圖3 草地NPPA模擬值與實測值的一致性檢驗Fig. 3 Consistency test between the measured value of grassland NPP and the simulated value of NPPA
1.3.2 Theil-Sen median趨勢分析與Mann-Kendall檢驗
Theil-Sen median趨勢分析與Mann-Kendall檢驗相結合可用來判斷長時間序列數據的變化趨勢(Jiang et al.,2015)。其中,Theil-Sen median 趨勢分析的計算公式為:
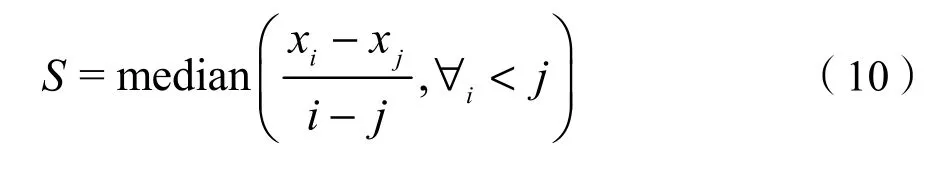
其中,1
Mann-Kendall檢驗用于判斷時間序列中是否具有上升或下降的趨勢,Mann-Kendall檢驗方法以其不受少數異常值干擾及不需要樣本遵從一定的分布的顯著優勢在水文、氣象研究中得到了廣泛的應用(Fensholt et al.,2012)。具體計算過程如下,將某時序的x值看作一組獨立分布的樣本數據,以參數Zc作為像元x衰減指標,計算公式如下:

其中,xk和xi為樣本時序數據集合;n為數據集合長度;sign為符號函數。在給定顯著性水平α下,當|Zc|>U1?α/2時,表示研究的時間序列數據在α水平上顯著的變化,其中,±Z1?α/2是標準正態偏差。在顯著性水平α=0.01、0.05和0.1時,對應的Z值分別為±2.576、±1.96及±1.645。在實際研究中一般取α=0.05的顯著性水平,當|Zc|>1.96時表示時間序列置信水平α<0.05,|Zc|<1.96表示置信水平α>0.05(劉洋等,2016)。
1.3.3 草地NPP變化的情景分析和定量評價
定量分析在草地恢復和草地退化過程中氣候變化和人類活動的相對貢獻,可通過計算草地實際NPPA、潛在NPPP和人類活動引起NPPH的變化趨勢S(NPPA)、S(NPPP)和S(NPPH)實現。S(NPPA)大于 0,代表的草地NPPA呈增加趨勢,即處于草地恢復狀態;S(NPPA)小于0,代表草地NPPA呈減少趨勢,即處于草地退化狀態。如果S(NPPP)為正,說明氣候變化促進了草地恢復;S(NPPP)為負,說明氣候變化促進了草地退化。如果S(NPPH)為正,說明人類活動促進了草地退化;S(NPPH)為負,說明人類活動促進了草地恢復。表1為本研究定義NPP變化的6種情景。
1.3.4 載畜壓力指數的計算
為了分析和評價放牧對草地NPPA的影響及草畜矛盾,本研究采用草地載畜壓力指數來對放牧壓力進行定量化(樊江文等,2011),公式如下:

式中,GPI為草地載畜壓力指數,CS是草地現實載畜量,CP為草地理論載畜量。如果GPI=1,則表明草地處于草畜平衡狀態;若GPI>1,說明草地已過度放牧;若GPI<1,說明草地尚有載畜能力。CS的計算方法如下:

CS為各個季節牧場的現實載畜量,即單位面積草地實際承載的羊單位數量(標準單位sheep·hm?2);Amount是每個季節牧場的 6 月末家畜存欄數(羊單位)(富蘊十二五統計年鑒,2016);Gt為不同季節草場利用時間(春秋牧場 155 d,夏牧場90 d,冬牧場120 d),Ar為每個鄉鎮不同季節牧場的草地面積。理論載畜量的計算公式為:
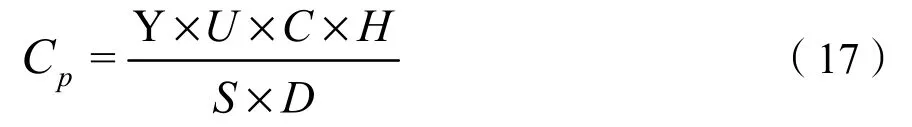
式中,Cp為理論載畜量(標準單位 sheep·hm?2);Y為單位面積草地的產草量(kg·hm?2),產草量的計算通過 CASA 模型計算出的 NPP (by C)/(g·m?2·a?1),除以0.475換算成干物質量后,再根據不同草地類型地下與地上生物量比例系數,計算地上產草量;U為牧草利用率;C為草地可利用率,H為草地可食牧草比率。S為一個羊單位家畜的日食干草量(羊單位日食量按每個羊單位每天采食4 kg鮮草計算,根據相關資料,牧草的干鮮比按1:3計算)。D為草地放牧利用時間。牧草利用率、草地可利用率和草地可食牧草比率的取值通過參考《天然草地合理載畜量的計算》(NY/T635—2002)標準(Su et al.,2003),并咨詢當地專家后確定。
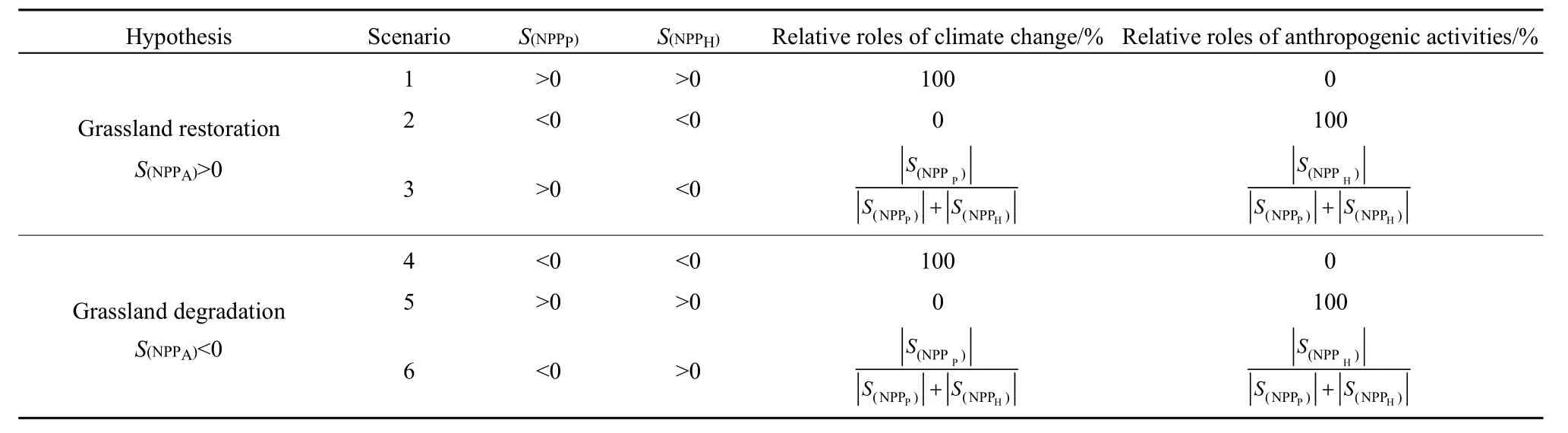
表1 不同情景下氣候變化和人類活動在草地NPPA變化中的相對作用Table 1 Scenarios of the respective influences of climate and anthropogenic activities on NPPA change in grasslands
2 結果
2.1 草地NPP的空間分布
2001—2015年平均NPPA和NPPP的空間分布特征均呈現出沿著海拔高度自北向南下降的趨勢(圖 4a、b)。研究區草地年平均 NPPA為 C 121.37 g·m?2·a?1,最大值為 C 652.26 g·m?2·a?1(圖 4a);草地年平均 NPPP為 C 363.57 g·m?2·a?1,最大值為 C 860.49 g·m?2·a?1(圖 4b);草地年平均 NPPH為 C 242.51 g·m?2·a?1,大部分地區 NPPH在 C 200—300 g·m?2·a?1之間(圖 4c)。
2.2 NPP變化趨勢的空間分異特征
采用 Theil-Sen median趨勢分析法,得到2001—2015年草地NPPA、NPPP和NPPH的變化趨勢S(NPP)。本研究根據S(NPP)的實際情況將S(NPP)分為5個等級,運用Mann-Kendall法進行顯著性檢驗。選取置信水平為0.05,根據顯著變化(|Z|>=1.96)和不顯著變化(|Z|<1.96)對結果進行分類(表2)。本研究以研究區最低海拔460 m為起點,將500 m作為一個海拔梯度,從而探討不同海拔梯度內的草地NPP的變化趨勢。
研究區 2001—2015年 NPPA變化趨勢如圖 5a所示。草地NPPA增加區域的面積占總草地面積的35.81%,且主要集中在研究區的北部山區(圖5a),輕微改善區域分別占1460—1960、1960—2460、2460—2960、2960—3460 m 海拔地區面積的68.84%,87.35%,75.30%和74.07%(圖6a)。無顯著變化區域的面積占草地總面積的 0.14%。草地NPPA退化區域的面積占64.05%,主要分布在調查區域的中南部(圖5a),輕微退化區域占據460—960 m及 960—1460 m 中低海拔地區面積的 72.44%和67.34%(圖6a)。如圖5b和圖6b所示,2001—2015年研究區氣候生產力模型估算的草地潛在 NPPP大體全部呈現出增加的趨勢。研究區近96.64%的區域草地潛在 NPPP呈增加趨勢,各個海拔梯度均以輕微改善趨勢為主。人類活動引起的NPPH明顯改善的區域的面積僅占0.02%;輕微改善的區域面積占7.35%,主要分布于研究區最北部的高海拔山地(1960—2460 m和2460—2960 m);輕微退化的區域的面積占85.81%,主要集中于1960 m以下地區;顯著退化的區域占6.81%,分布于研究區夏牧場的南部以及春秋牧場和冬牧場(1460 m以下)(圖5c、圖 6c)。
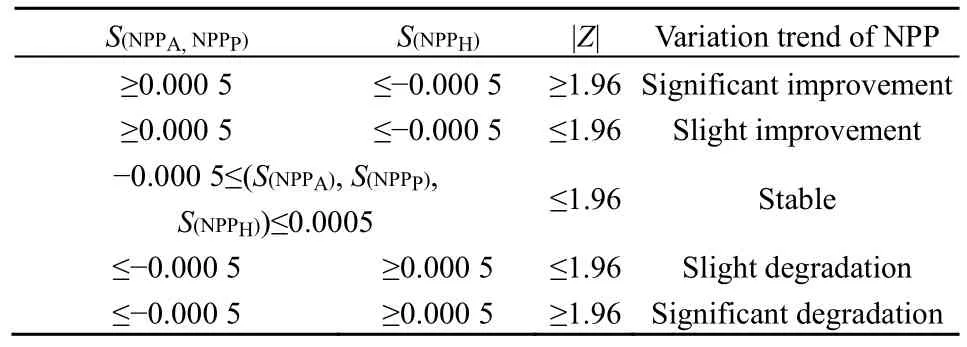
表2 研究區2001—2015年NPPA,NPPP and NPPH的變化趨勢Table 2 Variation trend and Significance levels of NPPA, NPPP and NPPH from 2001 to 2015
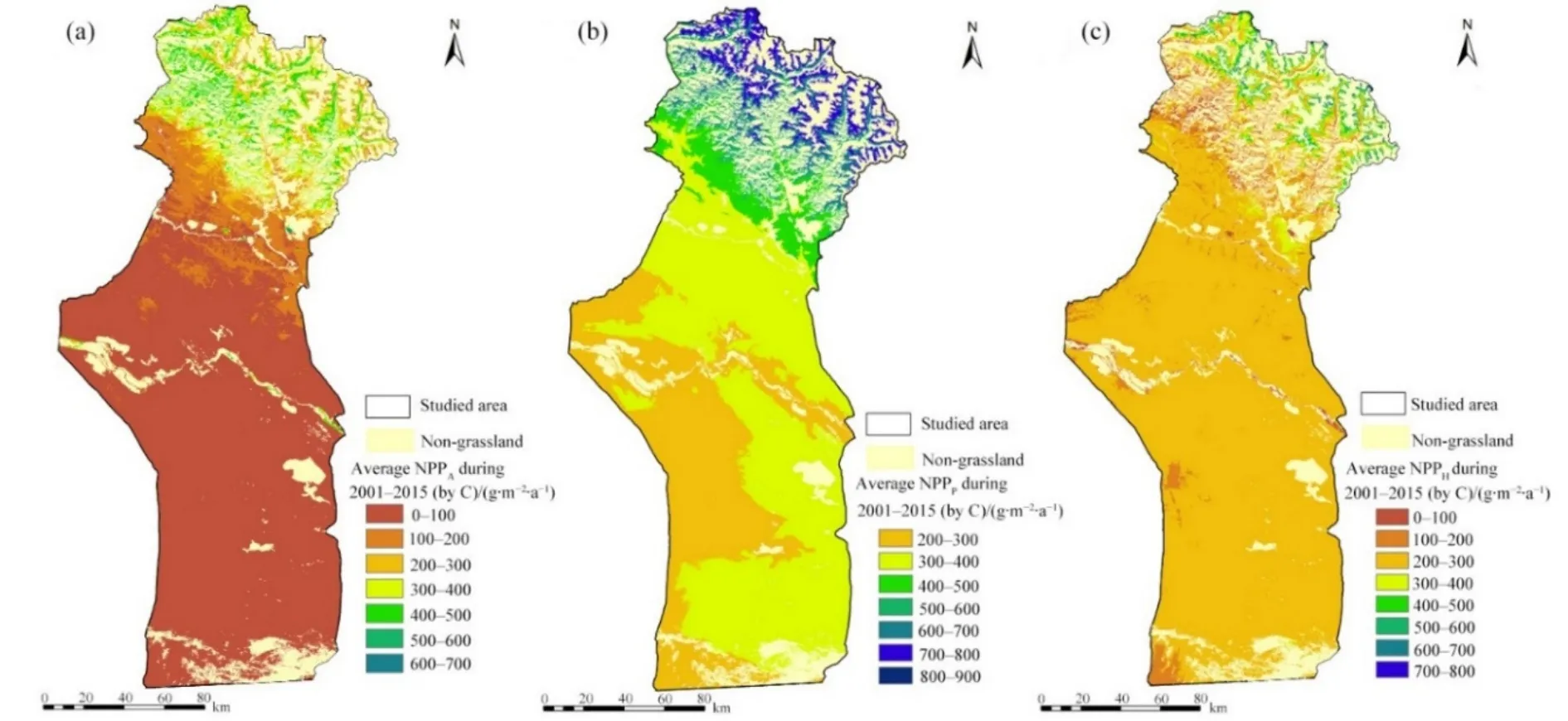
圖4 2001—2015年平均草地實際NPP的空間分布Fig. 4 Spatial distribution of grassland NPP from 2001 to 2015
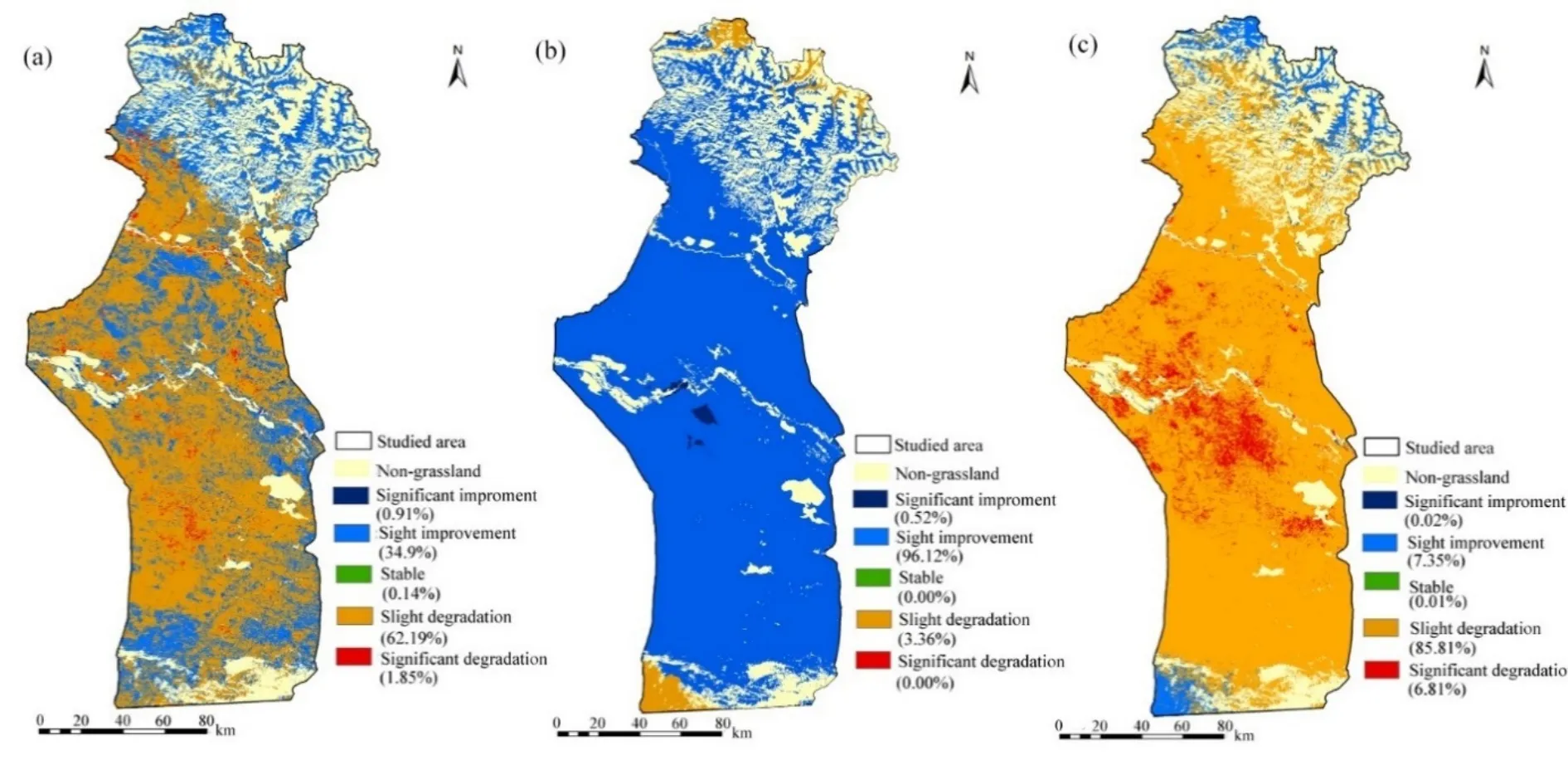
圖5 2001—2015草地NPPA,NPPP和NPPH的變化趨勢Fig. 5 Variation trend of grassland NPPA (a), NPPP (b) and NPPH (c) from 2001 to 2015
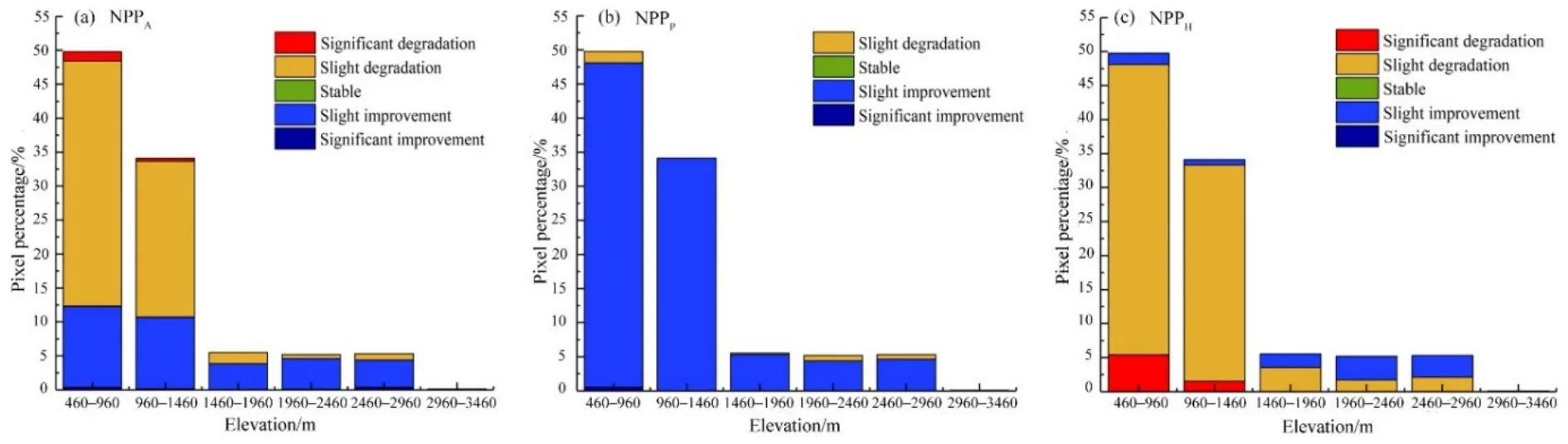
圖6 2001—2015不同海拔梯度內草地NPPA,NPPP和NPPH的變化趨勢Fig. 6 Variation trend of grassland NPPA (a), NPPP (b) and NPPH (c) in different elevation gradient from 2001 to 2015
2.3 氣候變化和人類活動的影響
圖7a顯示了2001—2015年氣候變化和人類活動對草地NPPA變化的相對作用的空間分布。總體來看,草地NPPA變化呈上升趨勢的區域所占面積(35.88%)小于呈下降趨勢的區域(64.12%)。2001—2015年,草地NPPA變化呈增加趨勢的面積占研究區草地面積的35.88%,主要分布于海拔1460 m以上的中高海拔山地。其中完全由氣候變化(CI)促進草原 NPPA增加的區域面積占 NPPA增加的區域總面積的81.30%,集中分布在夏牧場的南部;完全由人為活動(HI)引起的NPPA增加的區域面積僅占NPPA增加區域總面積的 6.19%,主要分布在夏牧場最北端;人類活動和氣候變化(BCI+BHI)共同促進草地 NPPA增加的區域占研究區 NPPA增加的區域總面積的12.51%,且主要集中分布在夏牧場的中部(圖7a)。草地NPPA變化呈減少區域的區域面積占研究區草地面積的64.12%,主要位于460—960 m及960—1460 m中低海拔地區。其中完全由氣候變化(CD)導致NPPA減少的區域面積占NPPA減少區域面積的1.04%;完全由人類活動(HD)導致的 NPPA減少的區域面積占 NPPA減少區域的總面積的98.21%,主要分布在研究區中南部的春秋牧場和夏牧場;氣候變化和人類活動(BCD+BHD)共同導致草地 NPPA減少的區域面積占 NPPA減少區域的總面積0.75%(圖7a)。
夏牧場草地 NPPA呈增加趨勢的區域面積(84.33%)遠遠大于草地NPPA呈降低趨勢的區域面積(15.67%)。人類活動和氣候變化及二者共同作用下促進草地 NPPA增加的面積分別占總 NPPA增加區域總面積的 13.87%,51.63%和 34.50%。對于NPPA減少區域,人類活動引起的草地NPPA減少的區域面積占總NPPA減少區域總面積的90.75%。人類活動和氣候變化共同影響下引起草地NPPA增加的區域集中分布在夏牧場的中部。完全由氣候變化引起夏牧場草地NPPA增加的區域集中分布在夏牧場的南部。草地NPPA呈增加趨勢的面積分別占春秋牧場和冬牧場面積的32.02%和25.60%,呈減少趨勢的面積分別占春秋牧場和冬牧場面積的67.98%和 74.40%。在春秋牧場和冬牧場中,NPPA增加區域主要由氣候變化所引起,NPPA減少區域主要由于人類活動所導致(圖7b)。
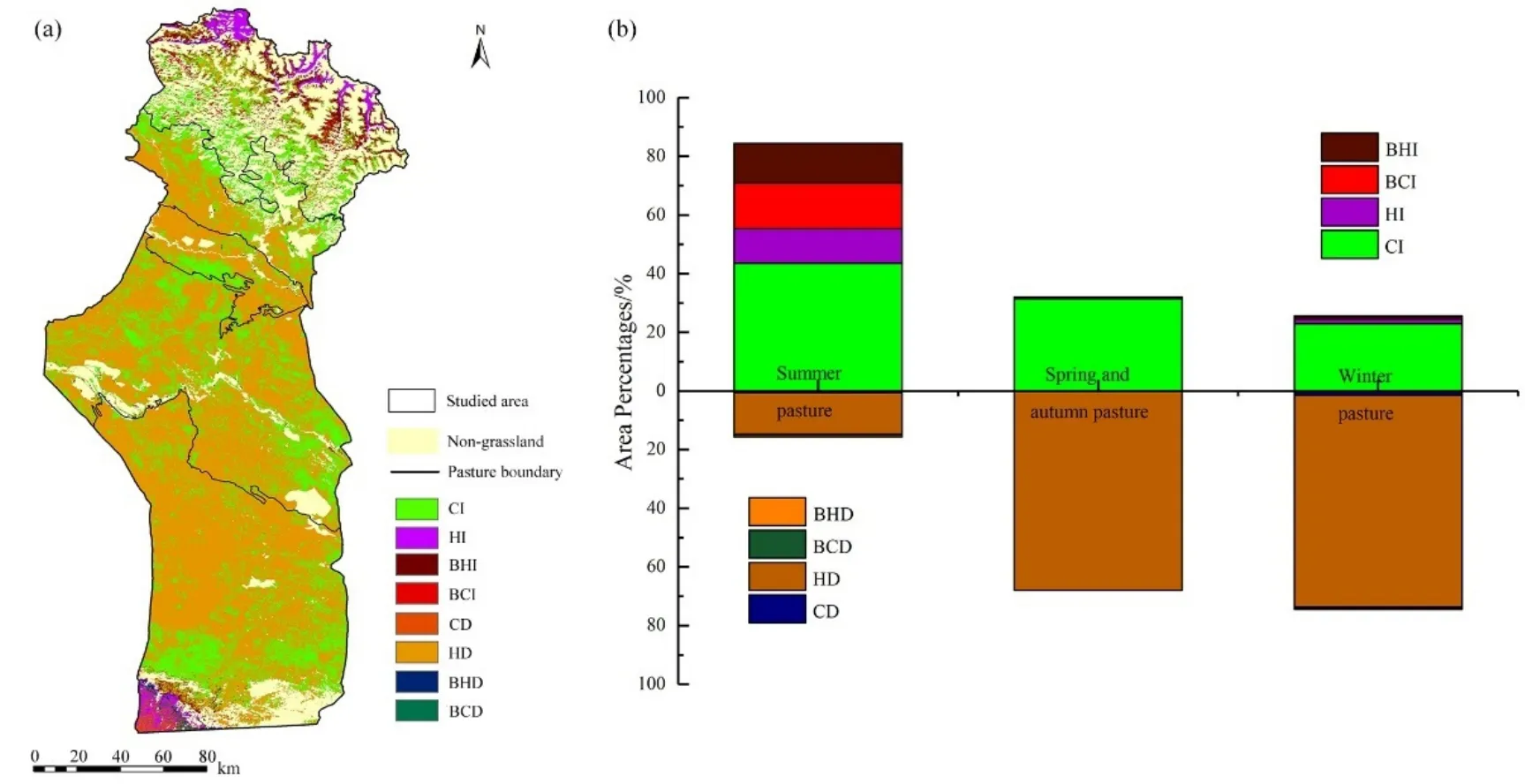
圖7 氣候變化和人類活動對草地NPP的影響Fig. 7 Effects of climate change and anthropogenic activities on grassland NPP
2.4 草地載畜壓力的時空變化特征
2001—2015年間,夏牧場、春秋牧場和冬牧場的平均理論載畜量分別為 1.32、0.25、0.30(單位sheep·hm?2);在年際變化上,夏牧場的理論載畜量呈上升趨勢,春秋牧場和冬牧場的理論載畜量略有下降。15年間,夏牧場、春秋牧場和冬牧場的平均現實載畜量分別為 1.36,0.72和 0.53(單位sheep·hm?2);在年際變化上,各季節牧場的現實載畜量均呈增加趨勢。夏牧場、春秋牧場和冬牧場的平均載畜壓力指數分別為1.04、3.03、1.83;在年際變化上,夏牧場的載畜壓力指數呈現出逐年下降的趨勢,而春秋牧場和冬牧場的載畜壓力指數呈現出逐年上升的趨勢(圖8)。
3 討論
3.1 氣候變化對草地NPP的影響
草地生態系統易受氣候變化的影響,在干旱地區尤其如此(Lioubimtseva,2004)。氣候變化可通過降水和溫度影響土壤水分和微生物,決定不同水熱條件下草地的生長狀況和生態系統生產力(He et al.,2015)。研究結果表明,氣候變化對草地NPP增加的貢獻率為81.30%,說明氣候因子在草地恢復的過程中起了顯著性的作用。為了進一步探討氣候因子對草地NPP變化的相對影響,采用了偏相關分析方法來探討2001—2015年間草地NPP對降水和氣溫變化的響應(圖9)。
年均NPPA與年降水量呈正相關的面積占研究區草地面積的85.16%(圖9b),研究結果表明,無論是山地、綠洲或荒漠地區,降水量的增加均是促進研究區草地NPPA增加的主導因素。以上研究結果與張山清等(2010)和張仁平(2017)研究結論一致,干旱和半干旱地區的植被NPP主要受降水控制。降水量是影響新疆NPPA變化的主要氣候因子,氣溫的影響相對較弱,且新疆地區草地NPPA與夏秋兩季降水具有明顯的正相關關系。降水是干旱區植被生長主要驅動因素,大氣降水是中亞干旱生態系統植被水分的主要來源(Zhang et al.,2017),直接決定了與植被生長息息相關的水資源可利用量。降水一般會提高土壤水分含量,使得土壤可為植被提供更多的水分供給,植被的光合作用增強,從而植被NPPA得到提高(Zheng et al.,2006)。研究區草地NPPA與降水負相關的比例占草地面積的14.84%,其中負相關的區域主要位于高海拔山地(圖9a、b)。當降水量的增加超過了植被生長的需要時,會引起太陽輻射的減少和相對濕度的增加,從而抑制植被的光合作用(Ukkola et al.,2016)。此外,過度降水會降低土壤有機質,加劇水土流失和洪澇災害,破壞植物棲息(Qu et al.,2018)。
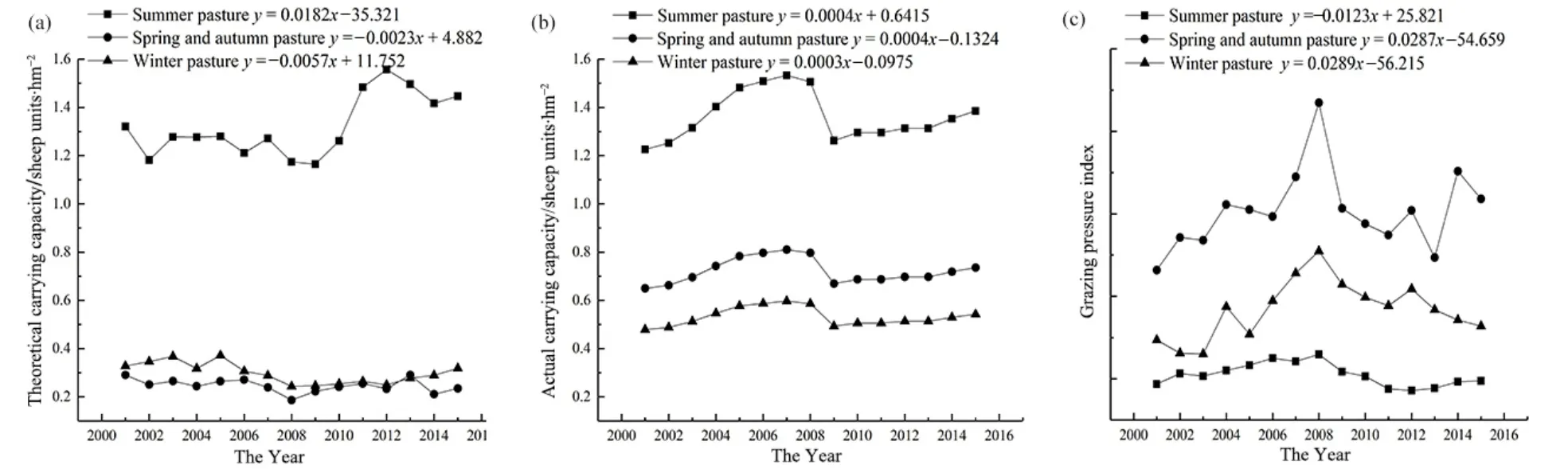
圖8 2001—2015年草地理論載畜量(a)、現實載畜量(b)和載畜壓力指數(c)變化動態Fig. 8 Changes of theoretical carrying capacity (a), actual carrying capacity (b) and grazing pressure index (c) in grassland from 2001 to 2015
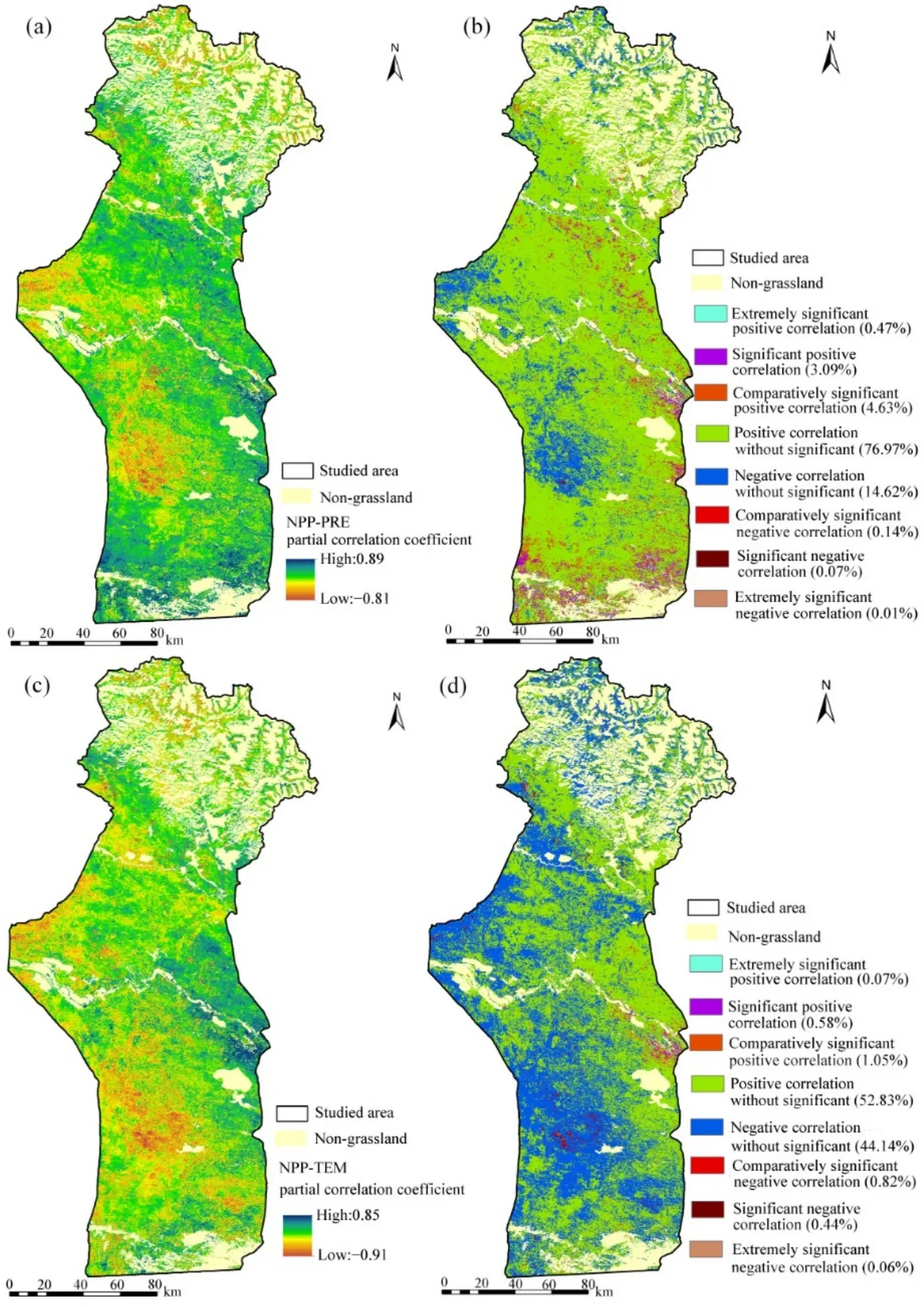
圖9 草地NPPA與氣象因子偏相關系數和顯著性水平時空分布圖Fig. 9 Spatial distributions of partial correlation coefficients and significant level between annual NPPA and climatic factors
年均NPPA與年平均氣溫呈正相關區域占研究區面積的 54.53%,呈負相關的區域占總面積的45.46%(圖9c、d)。草地NPPA對溫度變化的響應沒有明顯的區域分布規律。除此之外,草地NPPA與溫度之間幾乎沒有顯著的相關關系。Chen et al.(2019)的研究也指出,草地NPP對溫度變化不敏感,在中亞大部分地區溫度對草地的生長起著微不足道的作用。圖 10a顯示 2001—2015年 NPPA和NPPP的年際變化與降水變化趨勢大體一致,而與氣溫變化趨勢并不一致。研究結果表明區域平均NPPA和NPPP的最低值均出現在2008年(分別為C 94.05、279.24 g·m?2·a?1),可能由于 2007、2008 年的降水量最少所導致(圖10a)。這個現象表明,相比于氣溫,草地生長對降水更加敏感(Yang et al.,2016)。對于同一地點,降水的年際變化較大,而溫度年際變化較小,因此對于草地NPP的年際變化而言,降水是影響草地生長的決定性因素,特別是在干旱地區(Herrmann et al.,2005)。
此外,為了探討氣候變化對草地NPP的空間分布特征的影響,我們將研究區分為不同的海拔梯度,對草地NPP與降水和氣溫的相關性進行了研究(圖10b)。研究結果表明在460—960、960—1460、1460—1960、1960—2460 m 海拔梯度內,草地NPPA與年均降水量呈顯著正相關,與年均氣溫呈顯著負相關,且與年均降水量的相關性要高于年均氣溫。在2460—2960 m海拔梯度內,草地NPPA與年均降水量呈顯著負相關,而與年均氣溫呈顯著正相關,且與年均氣溫的相關系數要高于與年均降水量的相關系數。在2960—3460 m海拔梯度內,草地NPPA與年均降水量和年均氣溫均呈正相關,且與年均氣溫的相關性較大。研究表明,草地NPP與降水在整個研究區域的不同海拔上基本均成正相關(圖10b),而草地NPP與氣溫在中低海拔區域大部分地區呈負相關,在高海拔區域大部分地區呈正相關(圖 10b)。在溫度較高的中低海拔地區(460—2460 m),植被NPP對降水更為敏感,降水是草地NPP生長的限制性因素,降水的升高會促進草地 NPP的增加;而在降水較充沛的高海拔地區(2460 m以上),氣溫成為NPP生長的限制因素,植被NPP對氣溫更為敏感,氣溫的升高會促進草地NPP的增加(圖10b)。在新疆北部的低海拔地區,溫度較高,降水稀少,干旱限制了植被的生長(Guo et al.,2018)。降水可直接影響水資源可利用量,氣溫通過蒸發間接影響了水資源可利用量。降水量的變化比溫度的變化帶來更多的水資源可利用量的變化,因此,在中低海拔地區降水的影響更大。隨著海拔的升高,降水量增加,氣溫降低,低溫成為制約植被生長的更重要因素(Ma et al.,2019)。在溫度較低的區域,溫度的小幅升高可能會帶來較大的熱平衡增加,這可以解釋高海拔地區草地NPP對溫度的敏感性。
3.2 人類活動對草地NPP的影響
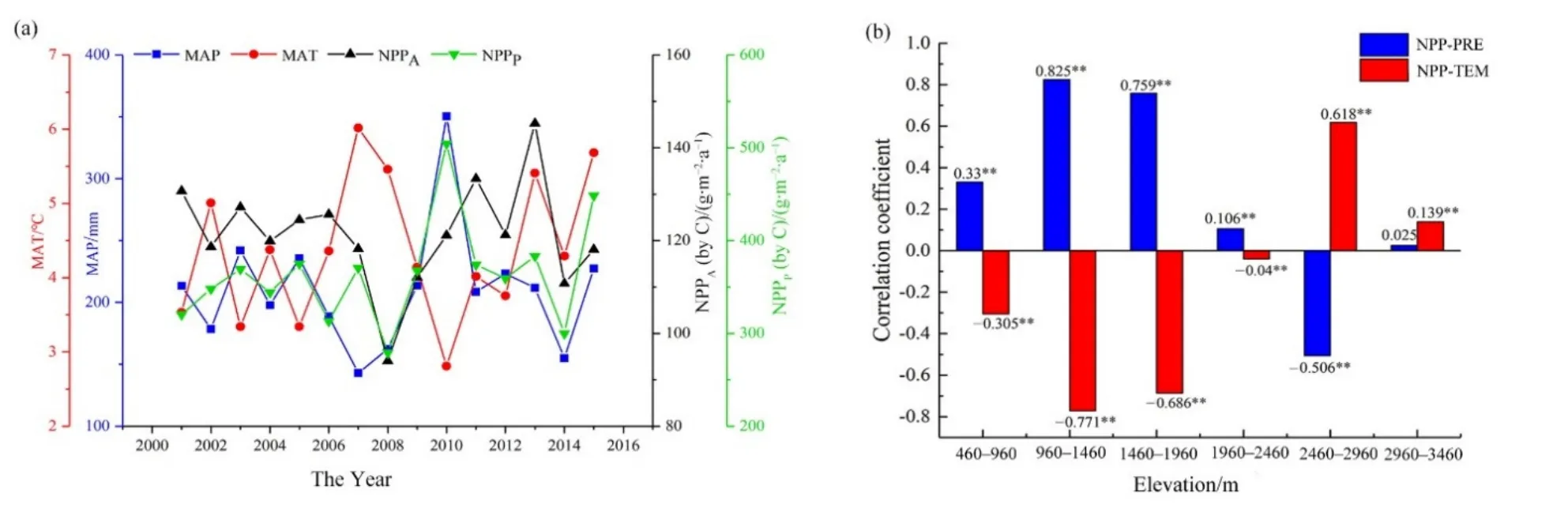
圖10 草地NPP與降水和氣溫的年際變化趨勢及NPPA與降水和氣溫的相關性Fig. 10 The annual change tendency of grassland NPP, precipitation and temperature and the correlation of NPPA with precipitation and temperature
已有研究表明,人類活動是草地動態變化的主要驅動力(Yang et al.,2016;Chen et al.,2017)。研究結果顯示,人類活動對草地NPP變化的相對影響為65.19%,而人類活動對草地NPP增加和減少的貢獻率分別為2.22%和62.97%。此外,輕微退化和顯著退化區域均位于海拔1960 m以下的中低海拔地區,春秋牧場和冬牧場的退化區域中有 100%和97%的區域都是由于人類活動所致。這一結果與Chen et al.(2019)的研究一致,在中亞地區荒漠草原比山地草原更易受到人類活動的影響。夏牧場十五年間平均草地壓力指數為1.04,基本處于草畜平衡狀態。十五年間夏牧場草地載畜壓力指數變化的斜率為正(圖8c),15年間載畜壓力指數的減少是夏牧場由人類活動為主導促進草地NPPA增加的直接原因。完全由人類活動引起的草地NPPA增加的區域(HI)分布在夏牧場的北部,即海拔較高的高寒草甸,該地區屬于邊遠牧場,由于路途遙遠、道路難行等原因人類和牲畜難以到達,放牧活動較少,草地受人類活動的干擾很少,因此放牧壓力的減小可能是草地NPPA增加的原因之一,這或許與近年來實施的草地生態保護工程措施有關(Wang et al.,2014)。完全由氣候變化引起夏牧場草地NPPA增加的區域集中分布在夏牧場的南部(圖7a),因為該區域海拔相對較低,放牧活動頻繁,人類活動對該區域的影響是使 NPPA減少,故該區域草地NPPA增加的完全是由于氣候變化所引起的。2001—2015年間春秋牧場和冬牧場的平均草地載畜壓力指數分別為3.03和1.83,超載率達203%和83%。春秋牧場的平均 NPPA較低,平均值為 C 103.72 g·m?2·a?1,本身載畜能力小,加之利用時間長達 155 d,利用時段又正是春季牧草萌發和秋季牧草結籽的關鍵時期,利用強度過重影響了牧草的生長繁殖。冬牧場草場面積與載畜量能力有很大差異,且NPPA平均值最低,僅為 C 70.79 g·m?2·a?1,處于草畜不平衡狀態。春秋牧場和冬牧場的載畜壓力指數變化的斜率均為正,呈現出增大的趨勢,因此放牧壓力增大是引起春秋牧場和冬牧場草地退化的主要原因,Chen et al.(2014)研究也指出放牧是人類活動引起草地變化的主要驅動力。
因此,需采取措施降低放牧強度,當地政府可實施牧民定居工程并提高牧民生計的多樣性,通過圈養并提供相應的基礎設施以降低草地放牧強度;此外,天然草地受氣候變化的影響較大,造成春秋牧場與冬牧場NPP的變異程度較大。需要開展穩定高產優質牧草的人工草地的建植,結合研究區的自然條件,建議在春秋牧場附近額爾齊斯河沿岸以及冬牧場附近烏倫古河沿岸開展優質高產的人工牧草的建植,為牲畜提供穩定充足的飼草料,以達到用小面積優質高產的人工草地建設來換取大面積天然草地的保護與修復的目的。可通過人工草地的建植,解決春秋牧場利用時間過長以及冬牧場草地面積與載畜量能力相差懸殊的難題。減輕春秋牧場和冬牧場的放牧壓力,緩解春秋牧場及冬牧場的草畜不平衡程度,使草地資源的利用向趨于合理的方向發展。
4 結論
本研究在模擬富蘊縣草地實際 NPP和潛在NPP的基礎上,定量分析了草地 NPP的時空變化特征,并對氣候變化和人類活動對草地NPP變化造成的影響進行了定量的分析,主要結論如下:
(1)實驗結果表明,2001—2015年間,草地NPPA的變化呈輕微下降趨勢。研究區35.88%區域的草地NPPA呈增加趨勢,64.12%區域的草地NPPA呈降低趨勢。氣候變化、人類活動及兩者共同促進草地NPPA增加的貢獻率分別為81.30%、6.19%和12.51%。2001—2015年期間,對于草地NPP的年際變化而言,降水是控制該地區草地NPP變化的主導氣候因子。對于草地NPP的空間分布而言,在溫度較高的中低海拔地區(460—2460 m),草地NPP與降水呈正相關,植被NPP對降水更為敏感;而在降水較充沛的高海拔地區(2460 m以上)草地NPP與氣溫呈正相關,且植被NPP對氣溫更為敏感。
(2)人類活動是草地NPPA減少的主要原因,人類活動引起草地 NPPA減少的區域占草地 NPPA減少總面積的98.21%,且主要發生在中低海拔的春秋牧場和冬牧場。
(3)夏牧場基本處于草畜平衡狀態,而春秋牧場和冬牧場處于草畜不平衡狀態,且春秋牧場的不平衡狀態表現尤為突出。草地載畜壓力指數的增加加劇了春秋牧場和冬牧場草地的退化。
猜你喜歡
少先隊活動(2022年5期)2022-06-06 03:45:04
家庭科學·新健康(2022年3期)2022-05-10 00:32:13
大科技·百科新說(2021年6期)2021-09-12 02:37:27
中老年保健(2021年2期)2021-08-22 07:31:10
好孩子畫報(2020年5期)2020-06-27 14:08:05
意林·全彩Color(2019年6期)2019-07-24 08:13:50
海峽姐妹(2018年3期)2018-05-09 08:20:40
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
奧秘(2015年2期)2015-09-10 07:22:44
