亞洲棉與擬似棉遠緣雜種的合成與鑒定
2020-08-28 16:35:50王一帆鄭赟榮二花吳玉香
生物技術通報 2020年8期
王一帆 鄭赟 榮二花 吳玉香
(山西農(nóng)業(yè)大學農(nóng)學院,太谷 030801)
棉花是世界上最主要的經(jīng)濟作物之一,中國是全球棉花種植面積最多的國家。棉花屬錦葵科(Malvaceae)、棉屬(Gossypium)植物,通過對棉花形態(tài)學、細胞學和地理分布等的研究,曾對棉花提出了多種分類的方法。Fryxell[1]將棉屬分為8個基因組,9個亞組,共包括50個種。之后,Wendel等[2]發(fā)現(xiàn)了2個野生種,并命名為艾克曼棉及斯提芬氏棉。因此,公認棉花共有52個種。中國是亞洲棉次級起源中心之一,亞洲棉早熟、耐旱和耐瘠能力強,對枯萎病也有很好的抗性,但由于栽培棉種質(zhì)資源遺傳多樣性較低,遠緣雜交是快速獲得新基因型并豐富種質(zhì)資源遺傳多樣性的方法,通過遠緣雜交將野生棉的有利性狀導入到栽培棉中,從而改善栽培棉的某些不良性狀[3]。經(jīng)過長期的自然進化與選擇,野生棉通過適應新的環(huán)境變化產(chǎn)生了大量的有利變異,使得其適應環(huán)境的能力增強。因此,野生棉中具有很多潛在的優(yōu)良基因(如D基因組擬似棉有抗寒、抗旱、抗棉鈴蟲、高抗枯萎病、耐銹病、無蜜腺的優(yōu)質(zhì)潛力)。郭旺珍等[4]利用SSR分子標記技術,對棉屬A、D染色體組的二倍體和四倍體代表種進行了遺傳多樣性分析,支持擬似棉是D染色體組最原始的棉種。因此,研究擬似棉的遺傳機理及利用其進行種質(zhì)創(chuàng)新也顯得尤為重要。
遠緣雜交是實現(xiàn)棉花種間雜交、基因重組及遺傳改良的重要途徑。通過遠緣雜交將各棉種的優(yōu)良性狀進行重組,再經(jīng)過篩選培育出高產(chǎn)、優(yōu)質(zhì)、穩(wěn)產(chǎn)的棉花新種質(zhì)[5]。許多棉花育種家通過遠緣雜交進行了大量試驗并取得了一定成果。馮文林等[6]將陸地棉與海島棉進行正反雜交,對后代農(nóng)藝性狀及纖維品質(zhì)的相關性進行測定分析并探究其規(guī)律,為棉花育種提供參考依據(jù)。申狀狀等[7]以陸地棉×斯特提棉組合雜交產(chǎn)生F1,并對雜種進行加倍,成功獲得異源六倍體種質(zhì),斯特提棉中種子腺體延緩發(fā)育性狀在異源六倍體中繼續(xù)表達。Konan等[8]對陸地棉×瑟伯氏棉×長萼棉回交一代的減數(shù)分裂行為與育性進行分析,為提高纖維細度特性提供良好的種質(zhì)資源。
國內(nèi)外關于亞洲棉與擬似棉雜交的相關研究鮮有報道,對擬似棉遺傳機理及致死原因的分子機制也鮮有報道。本研究利用亞洲棉與擬似棉進行遠緣雜交,并對獲得的F1雜種進行形態(tài)學及SSR分子標記鑒定,驗證雜種的真實性,以期對該新種質(zhì)進行加倍,為棉屬物種形成、基因組互作和雜交過程中親本基因組之間發(fā)生的染色體重組提供理論依據(jù),并為研究擬似棉致死機制提供新的種質(zhì),為異源多倍體進化研究提供方向。
1 材料與方法
1.1 材料
所用材料如表1所示,擬似棉分別于2014年、2015年、2016年取材于國家種質(zhì)三亞野生棉圃。

表1 研究材料基本信息
1.2 方法
1.2.1 遠緣雜交 以亞洲棉為母本、野生種擬似棉作父本進行雜交試驗,先將亞洲棉去雄,再將擬似棉的花粉涂抹到亞洲棉的柱頭上,每天上午進行授粉,并重復3次,間隔時間為24 h;并用滴管滴加0.1%的赤霉素在花柄基部用以保鈴。
1.2.2 形態(tài)學鑒定 收獲雜交后的F1種子,并將其與父母本的種子一同播種在營養(yǎng)缽中,待幼苗葉片完全展開時,觀察其形態(tài)特征和完全成熟的組織或器官,將亞擬雜種從植株的形態(tài)、葉片等與親本進行對比,從而初步鑒定雜種的真實性。
1.2.3 SSR分子標記鑒定 選取父母本及雜種植株上新鮮且幼嫩的葉片為材料,通過CTAB法提取葉片中的DNA,再使用銀染法檢測。選取SSR引物共12對(表2),引物來自CMD數(shù)據(jù)庫(http://www.cottonmarker.org,由北京賽百盛基因技術有限公司合成)。PCR反應體系為正、反向引物(2.5 μmol/L)各1 μL、10×Buffer(無Mg2+)2 μL、MgCl21.6 μL、Taq酶(5 U/μL)0.2 μL、dNTPs(10 mmol/L)0.2 μL,去離子水補足。PCR擴增程序為94℃ 5 min;94℃ 50 s,58℃ 50 s,72℃ 2 min,34個循環(huán);72℃10 min,4℃保存。用8%非變性聚丙烯酰胺凝膠電泳檢測。參考申狀狀等[9]方法進行銀染檢測。
1.2.4 數(shù)據(jù)統(tǒng)計與分析 選擇重復性好、條帶清晰的引物,進行重復試驗,選擇重復值相同的條帶用于統(tǒng)計及分析,統(tǒng)計雜種中雙親與新出現(xiàn)的條帶數(shù)。
2 結果
2.1 亞擬遠緣雜交
亞洲棉選擇苞葉和花蕾等長且發(fā)黃時的花蕾合適。但亞洲棉和擬似棉有生殖隔離不易成鈴,需要采用重復授粉和赤霉素保鈴等措施。雜交獲得的鈴大部分沒有種子,采用增加雜交花數(shù)量來保證部分鈴有種子。且雜交獲得的種子不飽滿,不易成苗,需采用精細管理保證成苗。由于亞擬遠緣雜種幼苗期有死亡現(xiàn)象,可能與擬似棉的致死基因有關。觀察發(fā)現(xiàn)強光照射加速雜種的衰老死亡,所以采用盆栽,便于管理,夏天搭建黑色網(wǎng)棚以避免強光照射,冬天轉(zhuǎn)移進溫室,保證雜種苗長成健壯植株。2014年雜交結果為:亞洲棉×擬似棉雜交花數(shù)為100,結鈴數(shù)為4,結鈴率為4.00%,飽滿種子數(shù)4粒,成苗1株。2015年雜交結果為:亞洲棉×擬似棉雜交花數(shù)為105,結鈴數(shù)為3,結鈴率為2.86%,飽滿種子數(shù)3粒,成苗2株。2016年雜交結果為:亞洲棉×擬似棉雜交花數(shù)為120,結鈴數(shù)為5,結鈴率為4.17%,飽滿種子數(shù)5粒,成苗2株(表3)。

表2 研究所使用的部分SSR多態(tài)性引物序列信息
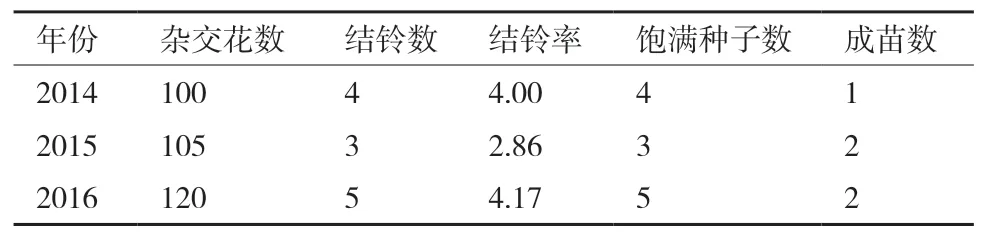
表3 亞擬遠緣雜交結果
2.2 亞擬雜種形態(tài)特征鑒定
亞擬雜種生長受阻且生長緩慢,在雜種植株未開花前就出現(xiàn)死亡現(xiàn)象,無法正常生長。但經(jīng)過避強光處理和溫室培養(yǎng),雜種植株健壯成長,優(yōu)于雙親,但雜種不現(xiàn)蕾不開花(圖1)。
對亞擬雜種的葉片與雙親進行比較(圖2)。亞洲棉呈掌狀分裂,葉裂片數(shù)在5-7片,裂刻及葉脈整體表現(xiàn)較父本顯著,各裂片間差異明顯,中間3個裂片較其他裂片大,且有規(guī)則,葉片表面有細小的茸毛;擬似棉葉裂數(shù)在3-5片,中間裂片較其他裂片大而長,呈柳葉狀,葉片表面無茸毛。亞擬雜種葉片整體形態(tài)偏向于擬似棉,大部分形狀介于兩者之間,葉裂片數(shù)3-5片,中間裂片呈漸尖細長狀,裂刻較雙親小,葉脈深淺基于雙親之間。
高溫對雜種的生長影響很大,會加速雜種的死亡,并且由于基因互作等關系,擬似棉與亞洲棉雜種致死基因呈顯性,雜種出現(xiàn)幼苗致死現(xiàn)象。

圖1 亞擬雜種植株與父母本形態(tài)學比較
2.3 亞擬雜種SSR分子標記鑒定
通過SSR分子標記方法,利用12對引物對父母本及雜種進行DNA水平上的分析,共擴增出12組清晰的多態(tài)性條帶(表4)。對12組條帶進行統(tǒng)計分析(表5),在亞擬雜種中共有61條清晰條帶,其中有28條來自母本,25條來自父本,8條屬于新的重組條帶,遺傳比例分別為45.91%、40.98%和13.11%(圖3)。

圖2 亞擬雜種葉片與父母本的葉片比較
在這些條帶組中,有11組條帶擴增出的分子量范圍為50-500 bp,引物NAU1173擴增出的條帶分子量大于500 bp。在11組引物條帶中,引物BNL2656擴增出的是雙親的互補貢獻帶(圖4-A),在引物BNL4108、BNL4053、NAU2026、NAU1157、NAU1052、NAU1042中除了雙親的互補條帶還出現(xiàn)了新的條帶(圖4),表明雜交過程中基因組之間可能出現(xiàn)了基因互作或染色體重組。進一步從分子水平證明了雜種是組合亞洲棉和擬似棉遺傳成分的真雜種。
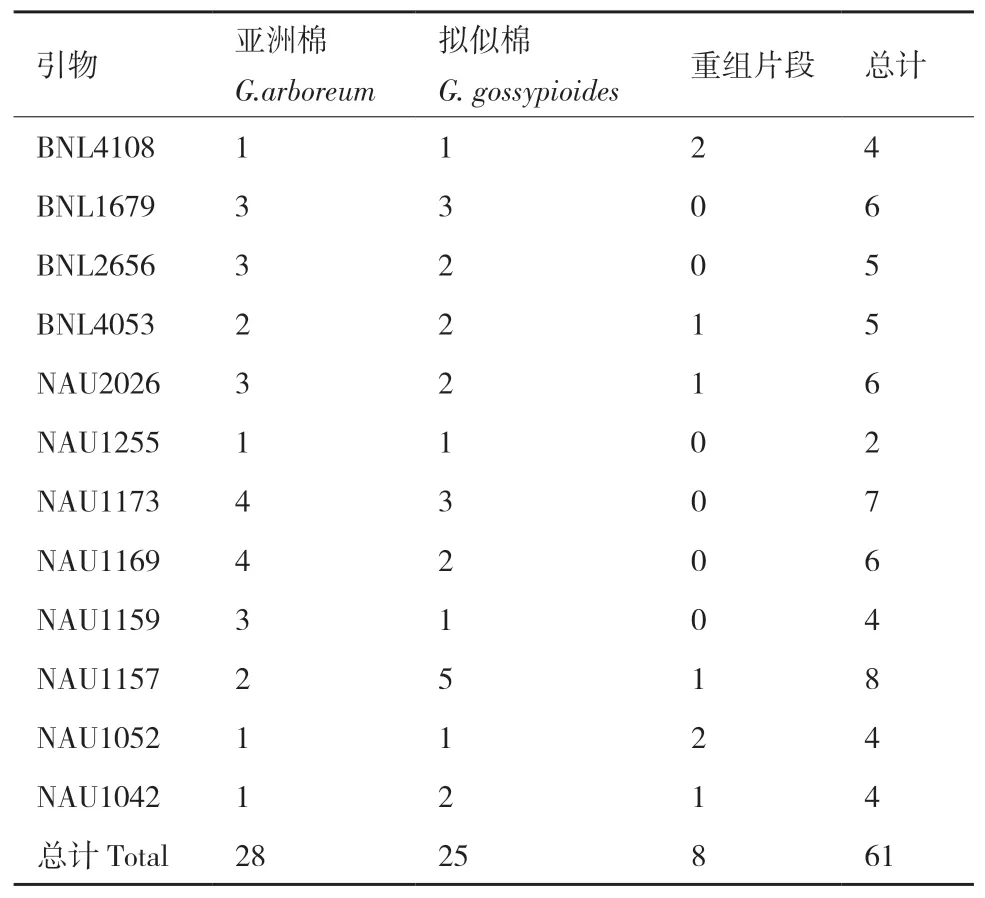
表4 亞擬雜種的SSR擴增結果表
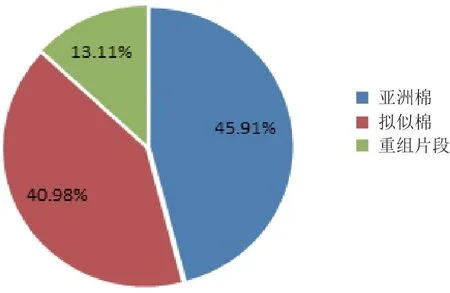
圖3 亞擬雜種的SSR多態(tài)性來源

表5 亞擬雜種的SSR多態(tài)性來源
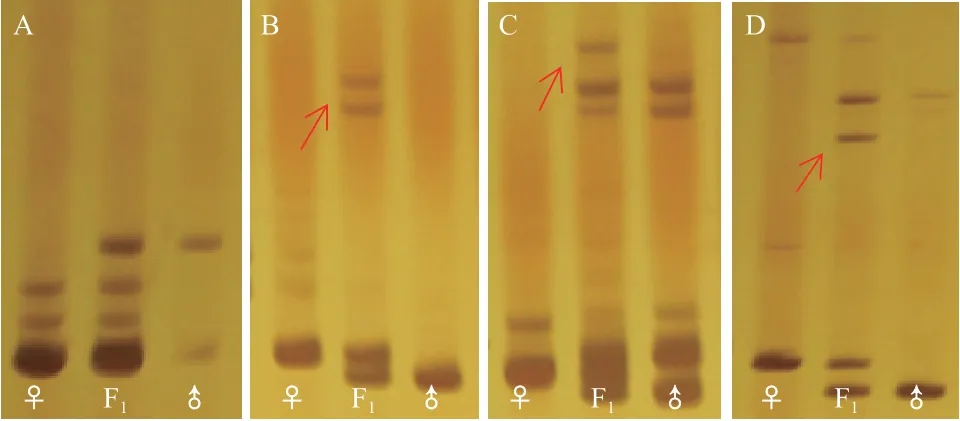
圖4 SSR引物在亞擬雜種及父母本中的擴增結果
3 討論
3.1 遠緣雜交與基因組互作
遠緣雜交是拓寬種質(zhì)遺傳基礎的重要途徑之一[10]。為了擴大棉花的遺傳多樣性,需要引進野生棉的潛在基因,通過遠緣雜交手段,培育出適應性強、綜合性狀優(yōu)良的棉花新種質(zhì)。然而,通過遠緣雜交之后的種質(zhì)并不能馬上應用到生產(chǎn)上,需要經(jīng)過進一步的選育才能成為生產(chǎn)應用的新品種。肖松華等[11]通過將栽培棉與野生棉雜交培育出了209份有陸地棉遺傳背景的種質(zhì)材料,并利用其中65份材料,創(chuàng)制出了抗黃萎病的新品種。郭寶生等[12]對陸地棉與海島棉、瑟伯氏棉雜交后代進行改良,經(jīng)過多年連續(xù)定向選擇,獲得類型豐富的改良材料493份。并不是所有材料都能成功的進行遠緣雜交,由于棉屬不同種在形成進化過程中有不同的遺傳背景,所以常會伴隨著雜交不親和、雜交不育或雜種敗育、雜交后代出現(xiàn)瘋狂分離等障礙[13]。想要克服這些障礙,最重要的就是消除種間生殖隔離。生殖隔離包括合子前(如花粉不能在異種柱頭上萌發(fā))與合子后(雜種不育及敗育)隔離[14]。本研究中選取的親本都是二倍體,雖然部分雜交出現(xiàn)合子前不親和的現(xiàn)象,但最終成功獲得了雜種,產(chǎn)生合子后不親和現(xiàn)象即亞擬雜種幼苗致死,這與基因的互作有關。基因互作是造成亞擬雜種不能成活的重要原因,不同位點基因互作不親和(Bateson-dobzhanskymuller incompatibilities,BDM模型)認為由于物種在長期的選擇進化過程中積累了大量的遺傳變異,在雜交重組后這些基因會產(chǎn)生不良的互作效應,造成雜種的敗育等不良現(xiàn)象[15]。
亞擬雜種中不僅結合了雙親的條帶,還出現(xiàn)了新的帶,這是雜交過程中發(fā)生了基因的互作及染色體重組的結果。雜種生長過程中可以通過植物激素、染色體加倍、嫁接、強制回交等方法來調(diào)節(jié)其生理狀況或恢復其育性。王志龍等[16]研究了海島棉與擬似棉F1不育原因,并通過植物激素克服了一些雜交不親和及雜種不育。基因組互作是促成生物進化的重要原因之一,本研究為探究擬似棉與亞洲棉基因組互作提供了理論基礎,解釋了亞擬雜種敗育的重要原因。
3.2 雜種鑒定方法探討
棉花屬常異花授粉作物,所以在雜交過程中需要仔細處理去雄,去雄不徹底會造成假雜種,因此,鑒定雜種的真假具有十分重要的意義。一般鑒定雜種的方法有形態(tài)學鑒定、細胞學鑒定以及分子鑒定等。形態(tài)學鑒定直觀、簡單,對雜種初步鑒定起到重要作用。細胞學鑒定能夠通過父母染色體數(shù)不同時,鑒定染色體數(shù)目判斷真假雜種,但當父母本染色體數(shù)一致時,這種方法鑒定具有局限性[17]。分子標記鑒定由于其可靠性高,所以廣泛應用于鑒定雜種中,其中SSR因其操作簡單、穩(wěn)定準確被廣泛應用于真假雜種的鑒定及雜種純度的鑒定[18]。付小瓊等[19]利用SSR對親本未知的F1雜種及其產(chǎn)生的F2群體進行了種質(zhì)真實度與純度的鑒定,并且構建了F1的SSR指紋圖譜,最終驗證了F1雜種純度高,F(xiàn)2出現(xiàn)了分離現(xiàn)象且雜合程度高。申狀狀等[9]對雜交育成的亞瑟雜種進行SSR分子鑒定,進一步印證了雜種的真實性。石建斌等[20]采用篩選出的26對SSR引物,對保存的58份棉花種質(zhì)資源進行純度檢測和遺傳多樣性分析,證明了品種資源的DNA聚類與品種的親緣關系相關性較高,表明棉花種質(zhì)資源遺傳的多樣性與復雜性。
3.3 雜種致死及半致死
雜種致死一般也稱為雜種劣勢,指生長旺盛的親本在雜交后產(chǎn)生的F1往往會有黃化、變色、生長矮小以及生長勢降低等現(xiàn)象,最終導致植株死亡[21]。戴寶生[22]報道了陸地棉與海島棉雜種節(jié)間縮短、植株矮小最終花葉脫落導致死亡;Phillips等[23]報道了陸地棉與擬似棉雜種植株矮小葉片失綠、莖稈變脆導致植株死亡,又發(fā)現(xiàn)了克勞茨基棉與戴維遜氏棉雜種在幼株或胚胎時期形成遺傳瘤,新葉的生長速度與老葉的壞死速度相等,即半致死現(xiàn)象。本研究亞洲棉與擬似棉的雜種同樣是節(jié)間縮短、植株矮小、葉片從下方開始變紅最終植株無法進行光合作用導致死亡。本研究通過遠緣雜交形成亞擬雜種,為研究擬似棉遺傳機理及致死原因提供新種質(zhì),并且后續(xù)可以通過測序及生信分析等手段,確定致死的遺傳模式,從分子水平解釋擬似棉遺傳規(guī)律及致死遺傳機制。
3.4 棉屬多倍化
棉屬的四倍體棉種的形成來源于A、D二倍體基因組,Wendel等[24]、Li等[25]認為四倍體棉種中A基因組由亞洲棉提供,而Wang等[26]、梅磊等[27]支持四倍體中D基因組的供體種是雷蒙德氏棉。但A基因組的供體種(亞洲棉)證據(jù)要比D基因組供體種(雷蒙德氏棉)的證據(jù)更明確,所以學術界一直對四倍體供體種的起源爭論不斷。郭旺珍等[4]利用SSR標記對10個二倍體棉種進行了聚類,結果表明,擬似棉是D基因組最原始的棉種。吳玉香[28]通過RAPD+SSR標記對異源四倍體棉種與A、D基因組中可能的供體種進行了遺傳相似系數(shù)的分析,結果表明,擬似棉的平均遺傳相似系數(shù)僅次于雷蒙德氏棉。因此,研究亞擬雜種的基因表達可以為研究棉屬多倍體化提供一定的方向,為尋找D亞基因組供體種提供證據(jù),揭示棉屬多倍體進化規(guī)律。以期對該新種質(zhì)進行加倍,對探究異源四倍體棉種的起源也具有十分重要的意義。
4 結論
克服了亞洲棉×擬似棉生殖隔離,從分子水平證明了F1雜種的真實性,同時在雜交過程中發(fā)生了基因互作及染色體重組。
