水稻抽穗期基因OsDof6 功能的初步研究
2020-09-17 06:16:06張立成李懿星王天抗邱牡丹宋書鋒董皓李磊劉建豐李莉
中國水稻科學 2020年5期
張立成 李懿星 王天抗 邱牡丹 宋書鋒 董皓, 李磊 劉建豐 李莉,
(1 湖南農業大學 農學院, 長沙 410128;2 雜交水稻國家重點實驗室 湖南雜交水稻研究中心, 長沙 410125;3 湖南大學研究生院 隆平分院, 長沙410125;*通信聯系人, E-mail: lili@hhrrc.ac.cn; liujf501@aliyun.com)
抽穗期是水稻最重要的農藝性狀之一,與產量密切相關,由主效和微效QTL/基因共同控制。近年來國內外許多科學家對水稻抽穗開花機制進行了大量深入的研究。
Dof(DNA binding with one finger)是一類植物特有的轉錄因子,它屬于鋅指蛋白超家族(zinc finger super-family),一般由200~400 個氨基酸組成[1-2],其N 末端含有由4 個Cys 殘基與Zn2+共價結合形成的單鋅指保守DNA 結合域,其C 末端含轉錄調控結構域[3]。N 末端的Dof 結構域由52 個氨基酸組成且含有保守的C2-C2型單鋅指結構域[4]。Zn2+和Cys 殘基是維持Dof 蛋白功能所必需的,對二者的替換或破壞都會使Dof 蛋白的活性喪失[5]。Dof 結構域中的單鋅指及鋅指旁邊C 側鏈狀結構中某些特定氨基酸均能與DNA 結合,這種DNA 與蛋白質雙功能結合活性,使Dof 蛋白可以調控多種基因的表達[6]。Dof 蛋白在植物生長發育過程中參與了激素響應、貯藏蛋白表達和光周期響應等生物學過程。如煙草的Dof 蛋白NtBBF1 可調節受生長素誘導表達的植物癌基因rolB的表達[7];水稻中的Dof 蛋白RPBF 可與RISBZ1 相互作用調節胚乳特異性貯藏蛋白[8],OsDof12過量表達植株在長日條件下能夠通過上調表達Hd3a和OsMADS14基因來縮短水稻抽穗時間[9]。擬南芥中的AtDof5.1可以調節葉片的軸向卷曲[10];OBP1在水楊酸和氧化脅迫下調控植物防御基因表達[11];OBP3通過調節光敏色素與隱花素來影響植物發育過程中的光形態[12]。
Yanagisawa 等[13]在玉米中鑒定了第一個Dof蛋白MNB1 后,Dof 家族進入研究者的視界,目前在擬南芥和水稻基因組中共預測存在36 個和30 個Dof基因[14],在小麥、大麥和大豆中分別預測存在31、26、28 個[15-17]。Dof 家族廣泛存在于整個植物界[18],但其中大部分基因的功能尚未明晰。
我們前期通過穗發育芯片篩選到大量可能與穗發育相關的基因,本研究選取其中的OsDof6,采用反向遺傳學的方法,利用CRISPR/Cas9 技術對該基因進行定點敲除,期望通過對突變體性狀進行觀察來獲得該基因功能,并通過煙草瞬時表達與實時定量PCR 等方式獲得該基因的表達模式,為深入了解該基因所參與的生命進程奠定基礎。
1 材料與方法
1.1 實驗材料
實驗所用粳稻品種9522(武運粳7 號),所用大腸桿菌菌株DH5α、農桿菌菌株EHA105 均保存于雜交水稻國家重點實驗室;CRISPR/Cas9 載體系統由華南農業大學劉耀光課題組提供。將T1突變體和9522 野生型種植于湖南省長沙市湖南雜交水稻研究中心試驗基地,種植密度為16 cm×20 cm,每行8 株,每個群體種植3 行,2019 年6 月5 日播種,6 月25 日移秧,常規大田管理。
1.2 生物信息學分析及進化樹構建
在TIGR(http://rice.plantbiology.msu.edu/)數據庫中檢索OsDof6,獲得該基因的基因組、CDS、ATG 上游1500 bp 啟動子序列及蛋白序列。將該基因蛋白序列輸入 SMART ( http://smart.emblheidelberg.de/)獲得該蛋白的預測結構,將啟動子序列輸入至PlantCARE ( http://bioinformatics.psb.ugent.be/webtools/plantcare/html/),獲得該啟動子中可能的調控元件,在 PlantTFDB (Plant Transcription Factor Database, planttfdb.cbi.pku.edu.cn)數據庫中檢索Dof,并根據周淑芬[19]、紀劍輝等[20]的研究,確定了30 個Dof 家族成員的基因,利用ClustalX 軟件對這30 個基因的序列進行多重序列比較,制作水稻Dof 家族進化樹。
1.3 基因組織表達分析
利用TRIZOL,分別提取水稻根、莖、葉、穗(穗長1~2 mm、5~10 mm、15~50 mm、50~100 mm、穗變綠及即將抽穗等6 個時期)各個部位的總RNA,逆轉錄cDNA,利用在線工具(http://www.oligoarchitect.com/LoginServlet)設計OsDof6的qPCR 引物qPCR-Dof6-F 和qPCR-Dof6-R(表1),以各個部位cDNA 為模板,以Ubq(Os03g13170)為內參,對OsDof6各個時期的表達水平進行檢測。
1.4 OsDof6 亞細胞定位載體構建及煙草瞬時表達
利用同源重組的方式,設計引物1300-Dof6-F和1305-Dof6-R(表1)擴增OsDof6去掉終止密碼子的CDS 序列并連接到pCAMBIA1305 載體上,獲得1305-Dof6-GFP 表達載體,將該表達載體轉入大腸桿菌涂板,挑取單菌落進行PCR 并測序,選擇測序結果正確的表達載體轉化入農桿菌中,將含有1305-Dof6-GFP 載體、1305-GFP 載體和p19 載體的農桿菌單克隆搖菌過夜,次日取出,4 000 r/min 下離心10 min,調OD 值至0.6 左右,加入1/50 體積0.5 mol/L MES(pH=5.6)和1/500 體積的0.1 mol/L乙酰丁香酮,混勻后室溫靜置3 h。將含有表達載體的農桿菌菌液和p19 農桿菌菌液按照1∶1 體積比混合,從煙草葉片背軸面注射,避光培養36~48 h,取葉片在激光共聚焦顯微鏡下觀察熒光信號。
1.5 CRISPR/Cas9 特異性靶點選擇及載體構建
將OsDof6的基因序列輸入E-CRISP (www.ecrisp.org)靶標位點設計網站中,對PAM 及5′端堿基要求選擇medium 選項,選擇靠近起始密碼子或保守結構域處NGG 上游20 bp 序列,將所有結果通過NCBI 進行BLAST,選擇特異性較強的一個,即CCGGTACAAGCAGGGCGCCGAGG,靶位點標記為T1,設計靶位點引物Dof6-U3-F 和Dof6-U3-R。載體構建、鑒定與農桿菌轉化參考鄧堯的方法[21]。

表1 引物序列Table 1.Primer sequence.
1.6 農桿菌介導水稻愈傷組織轉化及轉基因陽性植株基因型檢測
將成熟的9522 種子置于誘導培養基上誘導形成愈傷組織,用上一步的農桿菌菌液侵染愈傷組織,置于共培培養基20℃下暗培養3 d,之后將共培養后的愈傷組織用無菌水沖洗3~5 次,置于含潮霉素、頭孢噻肟鈉和氨芐青霉素篩選培養基上暗培養2 周后更換新的篩選培養基暗培養2 周,再經過分化培養、生根培養后,得到幼苗植株。將植株移入大田種植,提取T0代轉基因植株葉片基因組DNA。以該 DNA 為模板,用檢測引物 JC-Dof6-F 和JC-Dof6-R(表1)進行PCR 擴增,產物大小為491 bp。將PCR 產物送測序公司測序,Chromas 讀取測序峰圖,利用NCBI 網站BLAST 工具對序列進行對比,分析T0轉基因植株靶位點序列變化。
1.7 OsDof6突變體表型鑒定
參考趙金成[22]的方法,從見穗日起至齊穗為止,每天9:00-10:00 調查T1株系內所有植株的抽穗情況,主穗穗頂端伸出劍葉1 cm 記該單株抽穗,株系內50%單株抽穗為整個株系抽穗期,80%單株抽穗為齊穗期,同時統計株系內所有單株抽穗水平繪制抽穗動態圖。此外,調查突變體與對照的株高、分蘗數、穗長、每穗粒數、結實率、千粒重等。用DPS 7.5 數據處理系統對產量性狀等進行方差分析。
1.8 抽穗期相關基因差異表達分析
提取突變體與野生型穗發育初期葉片總RNA,利用實時熒光定量PCR 的方法,檢測抽穗期相關基因的表達量(引物見表1),分析這些基因在突變體與野生型中的表達水平差異。
2 結果與分析
2.1 OsDof6生物信息學特征
OsDof6位于水稻第1 染色體上,該基因全長993 bp,CDS 總長963 bp,含有1 個外顯子,編碼一個含有320 個氨基酸的蛋白(圖1-A)。利用PlantCARE 在線工具對該基因起始密碼子上游1.5 kb 啟動子序列進行預測分析,發現該基因的啟動子中包含大量的光響應元件,茉莉酸甲酯、赤霉素等植物激素響應元件,植物組織表達相關元件及逆境響應元件(表2)。表明該基因可能參與了植物光周期、激素調節、組織發育等相關生物學過程。利用SMART 對該基因的功能結構域進行預測,發現在其22~80 aa 處存在一個zf-Dof 保守結構域(圖1-B)。
高度保守的C2-C2 類型鋅指結構是植物Dof 轉錄因子家族的核心結構域。此外,在不同Dof 亞家族中,除C2-C2 類型鋅指結構域外,還存在一些相對保守的基序,然而序列的保守性在一定程度上能夠反映其在功能上的相似性[23]。為了預測OsDof6的功能,從PlantTFDB 上檢索了30 個水稻Dof 家族的候選基因,進一步對其氨基酸序列進行多重對比,構建出水稻Dof 家族的系統進化樹,發現OsDof6與OsDof20處于同一分支(圖1-C),對OsDof20的表達模式進行預測,發現該基因主要在花藥及穎殼的發育過程中表達。OsDof20的具體功能及其與OsDof6的關系有待進一步研究。
2.2 OsDof6的表達模式及編碼蛋白的亞細胞定位
利用RiceXpro 表達數據庫對OsDof6的時空表達模式進行分析。結果顯示,OsDof6在葉鞘、營養生長期的根部和生殖生長期的莖部表達量較高,在內外稃發育中后期表達量最高(圖2-A)。進一步利用實時定量PCR 對OsDof6在水稻根、莖、葉、穗P3 期(穗長1~2 mm)、穗P4 期(穗長5~10 mm)、穗P5 期(穗長15~50 mm)、穗P6 期(穗長50~100 mm)、穗P7 期(穗變綠)、穗P8 期(即將抽穗)等組織器官中的相對表達量進行檢測,發現OsDof6屬于組成型表達,在根、莖、葉和穗中均檢測到轉錄本,然而在穗發育的P7 期和P8 期表達水平明顯高于其他部位(圖2-B)。

圖1 OsDof6基因核苷酸序列(A)、蛋白結構(B)與家族進化分析(C)Fig.1.OsDof6 nucleotide sequence(A) and its protein structure(B) and phylogenetic analysis of OsDof family(C).
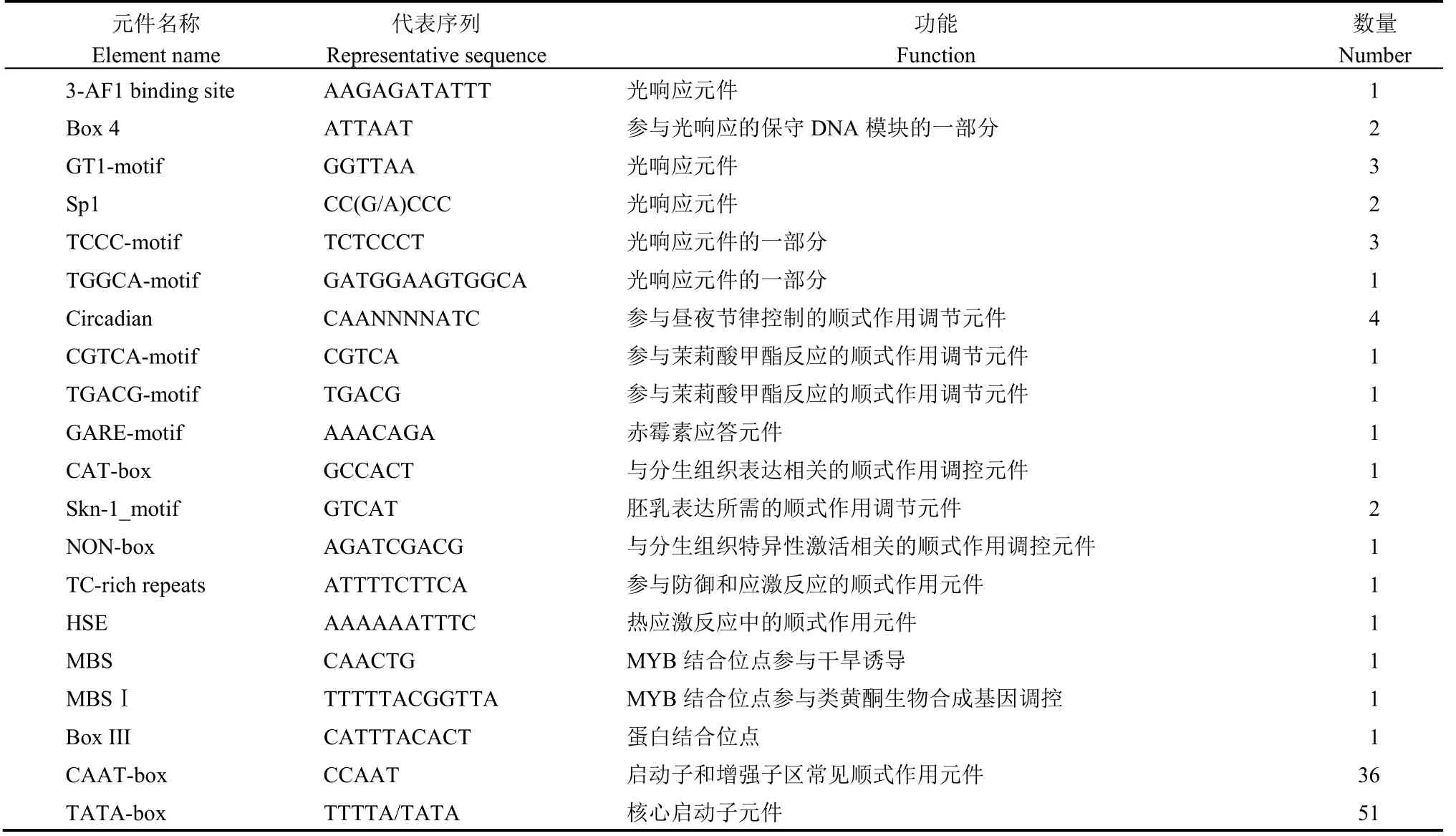
表2 OsDof6基因ATG 上游1.5 kb 啟動子區域功能元件分析Table 2.Functional element analysis of the 1.5 kb promoter region upstream ofOsDof6 gene ATG.
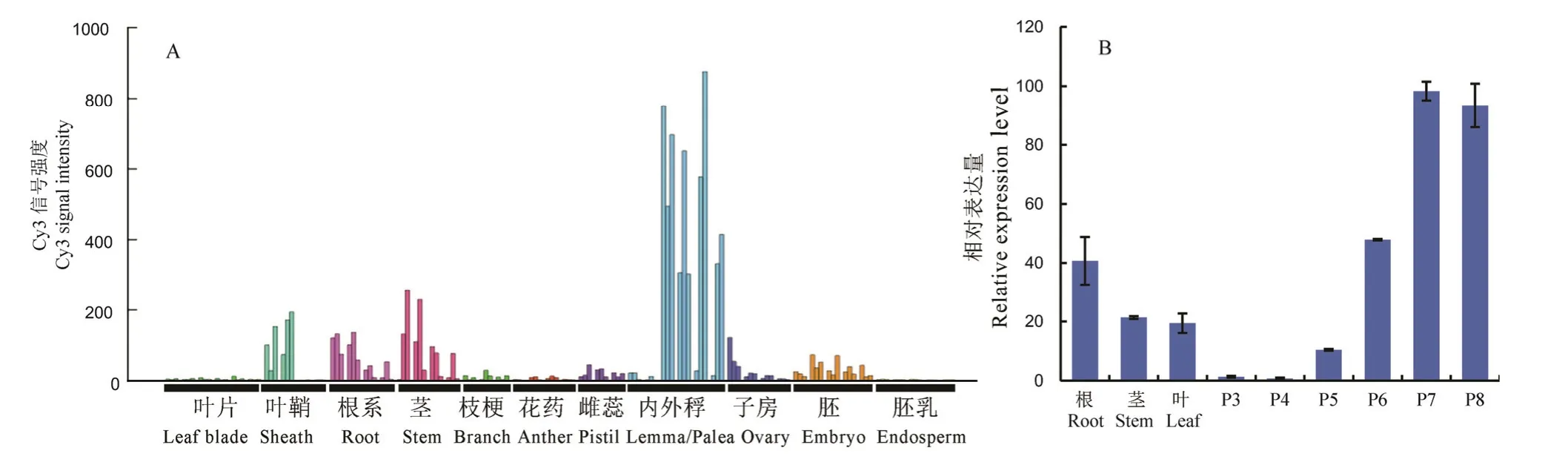
圖2 OsDof6基因時空表達分析Fig.2.Temporal and spatial expression analysis ofOsDof6 gene.
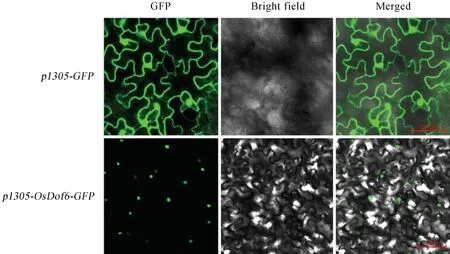
圖3 OsDof6亞細胞定位結果Fig.3.Subcellular localization result ofOsDof6.
通過同源重組的方式,將OsDof6的cDNA 序列(不包含終止密碼子TAA)整合到pCAMBIA1305-GFP載體上,獲得pCAMBIA1305-Dof6-GFP植物融合表達載體,將植物表達載體轉入農桿菌并侵染煙草葉片表皮細胞,通過激光共聚焦顯微鏡觀察Dof6-GFP 融合蛋白的熒光信號(圖3),Dof6-GFP融合蛋白主要定位在細胞核中,而在空載對照的細胞膜、細胞質和細胞核中均能檢測到熒光信號。該實驗結果表明,OsDof6 特異定位于細胞核中。
2.3 獲得OsDof6基因突變體植株
為了進一步研究OsDof6在水稻生長發育中的功能,構建了OsDof6基因編輯載體。經過PCR 鑒定與序列比對,OsDof6的靶位點成功構建入pYLCRISPR/Cas9-MT載體中,獲得了植物表達載體pYLCRISPR/Cas9-MT-Dof6(圖4-A)。將該植物表達載體轉入農桿菌菌株,通過農桿菌介導轉化水稻9522 愈傷組織并鑒定,共獲得4 株陽性轉基因植株9522Dof6-1、9522Dof6-2、9522Dof6-3和9522Dof6-4,其中9522Dof6-2、9522Dof6-3和9522Dof6-4在靶位點處出現突變,9522Dof6-1未發生突變。9522Dof6-2和9522Dof6-3具有相同的突變類型,該突變使得原基因序列的第62~69 bp 共8 個堿基缺失,直接導致閱讀框移碼致使翻譯提前終止,破壞了Dof 蛋白的結構,形成了一個與Dof蛋白完全不同的108 aa的蛋白(圖4-D)。9522Dof6-4使得原基因序列第11~313 bp 共303 個堿基丟失,即氨基酸序列第4~104 aa 共100 個氨基酸缺失,該突變使Dof 保守結構域完全缺失。
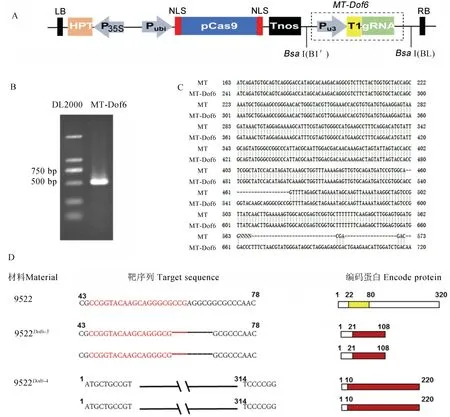
圖4 OsDof6基因編輯載體構建與突變體突變類型分析Fig.4.Construction ofOsDof6 gene editing vector and analysis of mutation types of mutants.
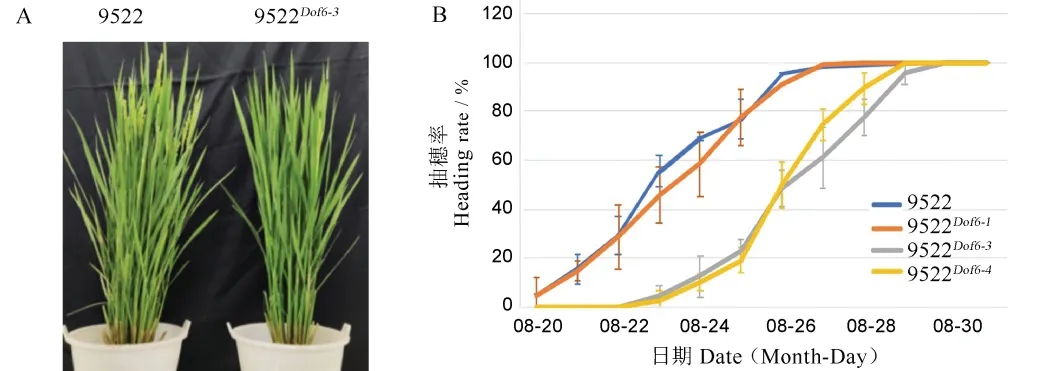
圖5 野生型9522 與Dof6 突變體的表型比較Fig.5.Phenotypic comparison between the wild type 9522 and theDof6 gene-knock-out mutant.
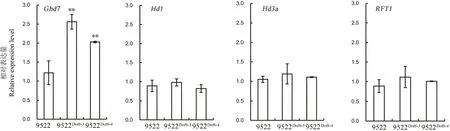
圖6 抽穗期主要調控基因差異表達分析Fig.6.Analysis of expression levels of main genes for heading date.
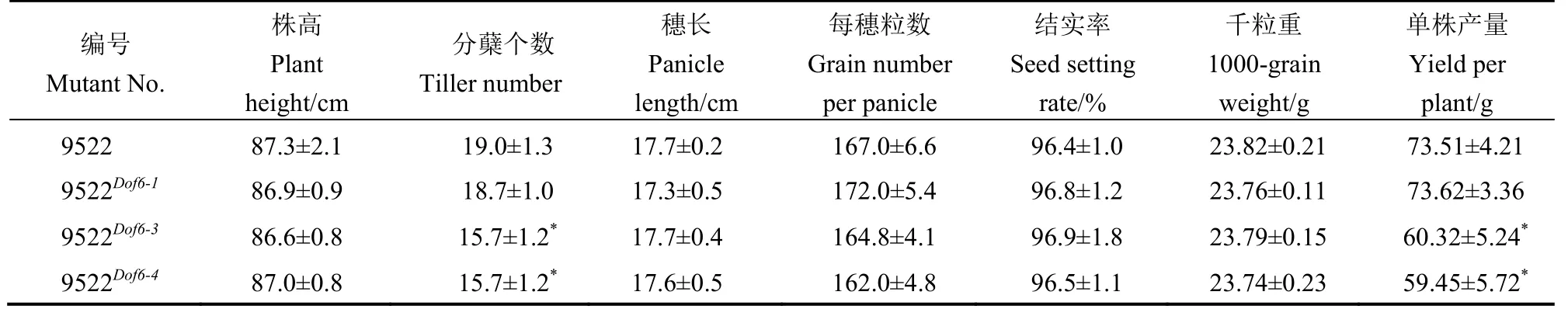
表3 突變體與對照的產量性狀調查Table 3.Investigation of yield traits in mutants and controls.
2.4 OsDof6突變體延遲抽穗
對9522Dof6-1(陰性對照)、9522Dof6-3、9522Dof6-4和9522 野生型的抽穗動態調查結果顯示,9522 野生型和9522Dof6-18 月20 日始穗,8 月24 日進入抽穗期,8 月26 日齊穗,播始歷期為77 d,而突變體9522Dof6-3和9522Dof6-48 月23 日始穗,8 月27 日進入抽穗期,8 月29 日齊穗(圖5-A,見上頁),播始歷期為80 d。突變體材料進入抽穗期的時間與野生型和陰性對照相比平均推遲3 d 左右(圖5-B)。但在抽穗歷期與抽穗速度上無顯著變化。對突變體與野生型9522 和陰性對照9522Dof6-1的產量性狀調查發現,突變體的平均分蘗數為15.70 和15.75 個,顯著低于陽性對照9522 野生型的19.0 個和陰性對照9522Dof6-1的18.7 個,而突變體與野生型在株高、穗長、每穗粒數、結實率、千粒重等產量相關因素上無顯著差異,突變體的單株產量由于分蘗數的下降而顯著降低(表3)。由此可知,OsDof6的功能缺失影響到突變體材料的抽穗期、分蘗數和單株產量。
2.5 OsDof6功能缺失影響了Ghd7 的表達
為了探索OsDof6基因所參與的分子調控途徑,選取了抽穗期調控網絡中的幾個重要的基因Hd1、Ghd7、Hd3a及RFT1。以幼穗發育初期葉片總RNA為模板,在突變體與野生型中進行了差異表達分析,結果顯示,在突變體中Ghd7基因的表達量顯著上調,Hd1基因的表達量沒有顯著變化;而成花素基因Hd3a及RFT1的表達量無顯著變化(圖6)。
3 討論
抽穗期是水稻一個重要的農藝性狀,它主要由品種的感溫性、感光性和基本營養生長性決定[22]。目前已有大量的水稻抽穗期相關QTL被精細定位并克隆。如開花促進基因DTH3[24]、Ehd4[25],開花抑制基因Hd16[26],成花素基因Hd3a[27]和RFT1[28]等水稻Dof家族部分成員也是具有調控抽穗期的功能。其中,OsDof12基因過量表達可在長日條件下上調Hd3a和OsMADS14基因的表達從而縮短抽穗期,但其過量表達在短日條件下對抽穗期無影響[9]。OsDof4的過量表達也會導致長日條件下抽穗提前,短日條件下抽穗延遲[29]。
本研究利用CRISPR/Cas9技術,對水稻品種9522的OsDof6基因進行定點敲除,獲得了兩種不同突變類型的純合功能缺失突變體。對這兩份突變體材料的T1代生育期觀察發現,突變體材料與野生型及陰性對照相比抽穗推遲3 d。而在產量性狀中,突變體材料除分蘗數與野生型相比顯著下降外,其他性狀均無明顯差異。并且,抽穗動態調查發現分蘗數的減少對抽穗歷期與抽穗速率無顯著影響。Dof家族中Dof功能結構域在不同物種間均表現出高度保守,其功能可能也存在著一定的保守。水稻Dof家族中OsDof12與OsDof4的功能均有報道為開花相關。對水稻Dof家族進行進化樹分析,發現與OsDof6親緣關系最近的成員是OsDof20。我們對其進行的時空表達模式預測結果顯示其主要在花藥發育與穎殼發育過程中表達,其具體功能有待進一步研究。值得注意的是,Tsuji等[30]發現成花素基因Hd3a在葉片韌皮部合成之后,除了會運輸到莖尖分生組織影響水稻抽穗外,還會在腋生分生組織中積累,以促進水稻分蘗。對OsDof6的時空表達進行研究發現該基因在水稻根、莖、葉中的表達量較高,在內外稃發育后期表達量最高,與預測結果吻合。對OsDof6序列分析得知在其啟動子中含有大量的光響應元件,少量組織表達元件、非生物脅迫響應元件以及激素響應元件,推測其可能具有響應光周期的功能。在抽穗期主要調控基因差異表達實驗中,Ghd7基因的表達量極顯著上調。Dai等[31]的研究表明,在長日照的環境下,Ghd7的表達量上升可以抑制Hd3a與RFT1的表達使水稻的抽穗期延遲,與Dof6突變體的表型一致。成花素基因Hd3a與RFT1表達量沒有顯著變化可能是由于抽穗期變化幅度較小、Dof家族基因功能冗余[32]所致。綜上所述,OsDof6在長日條件下可能參與Ghd7-Hd1-Hd3a/RFT1通路,通過光周期調節或調節穗部發育的方式來影響抽穗期。
后續將對該突變體進行過量表達、功能互補等研究對該表型進行進一步驗證。并對該基因的互作蛋白進行挖掘,以探究該基因在影響抽穗期過程中的分子機制及調控網絡。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
