利用CRISPR/Cas9 技術高效創制長粒香型水稻
2020-09-17 06:16:08徐善斌鄭洪亮劉利鋒卜慶云李秀峰鄒德堂
中國水稻科學 2020年5期
關鍵詞:水稻
徐善斌 鄭洪亮 劉利鋒 卜慶云 李秀峰, 鄒德堂,
(1 東北農業大學 寒地糧食作物種質創新與生理生態教育部重點實驗室, 哈爾濱 150030;2 中國科學院東北地理與農業生態研究所, 哈爾濱 150081;3 黑龍江省種業技術服務中心, 哈爾濱 150008;*通信聯系人, E-mail: lixiufeng@iga.ac.cn; 13603609603@163.com)
水稻作為全球最重要的糧食作物之一,全世界半數以上的人口均食用稻米,水稻是保障中國糧食安全與實現農業可持續發展的主糧作物[1]。近年來,人們對高產優質稻米的需求量越來越大,特別是長粒香型稻米,如稻花香的價格是一般非香型大米的2~4 倍,長粒香米更易受到農民和消費者們的青睞。傳統的水稻品種改良過程中,主要通過雜交、回交的方式實現優良基因的聚合,但傳統育種具有育種周期長、效率低等缺點,而基因編輯技術可以為水稻品種的快速改良帶來新的契機。
目前,CRISPR/Cas9 技術作為新一代的基因編輯技術,在作物遺傳育種研究中應用較廣泛[2-7],該技術可在水稻基因組水平上進行基因定向編輯改造[8-11]。水稻中已經鑒定出許多與籽粒大小相關的數量性狀位點(QTL),如負調控因子GW2[12]、GS3[13]、GS9[14]、qW5/GW5[15]和TGW6[16],正調控因 子GL2[17]、GL3.1[18]、GS5[19]、GL7[20]、GLW7/OsSPL13[21]和GW8/OsSPL16[22]。Li 等[23]利用該技術編輯水稻中的GS3基因,T2代的粒長和粒重增加,產量得到提高。Zhao 等[14]使用CRISPR/Cas9 技術,在GS9第1 外顯子處插入1 bp,造成移碼突變,破壞GS9正常表達,使粒長增加。
香味是水稻重要的品質性狀之一,由第8 染色體上的Badh2基因控制。Badh2編碼一個由503 個氨基酸組成的蛋白產物[24]。Chen 等[25]研究發現,在香稻品種中,由于Badh2基因發生突變,使得香味的主要成分2-乙酰-1-吡咯啉(2-AP)含量增加,稻米產生香味。Shan 等[26]通過TALEN 技術,對Badh2進行基因組編輯,稻米的2-AP 含量從無增加至0.35~0.75 mg/kg,獲得具有香味的稻米。孫慧宇等[27]利用CRISPR/Cas9 技術對Badh2進行編輯,使寒地粳稻東農425 香味獲得了改良,但東農425適應積溫區較小。因此,我們擬利用CRISPR/Cas9技術對黑龍江省第二、三積溫帶品種龍粳11 進行香味改良,從而擴大香稻的種植面積。龍粳11 屬早熟品種,其抗稻瘟病較強,抗倒伏,前、后生育時期耐寒性強。同時,龍粳11 的稻米品質優良,各項指標均達到國家優質米標準。
本研究利用CRISPR/Cas9 技術,以龍粳11 為研究材料,對GS3、GS9和Badh2基因同時進行敲除,能夠高效地將圓粒水稻變成長粒香型水稻,可為加快水稻的種質資源改良提供一定的理論指導。
1 材料與方法
1.1 試驗材料
本研究以粳稻龍粳11 為試驗材料。在黑龍江哈爾濱(北緯45°)和海南(北緯19°),均以25 cm×25 cm 的株行距種植,自然長日照條件下生長。
試驗所用 pYL-U6a-gRNA 和 pYLCRISPR/Cas9Pubi-H 雙元載體[28]由華南農業大學劉耀光院士惠贈。
1.2 gRNA 靶點接頭設計及載體構建
通過NCBI(https://www.ncbi.nlm.nih.gov/)獲得GS3(Os03g0407400)、GS9(Os09g0448500)、Badh2(Os08g0424500)的基因序列,利用CRISPRGE 在線網站(http://skl.scau.edu.cn/)[29],分別在GS3第1 外顯子、GS9第1 外顯子和Badh2第2 外顯子處設計1 對靶點接頭引物,B1-F/R、B2-F/R 及B3-F/R(表1)。利用軟件CRISPR Primer Designer設計GS3、GS9和Badh2基因靶序列,比對水稻序列,檢測設計靶點是否特異,排除潛在脫靶序列。
參照Ma 等[30]的實驗方法進行載體構建,將連接好的載體通過熱激法轉入大腸桿菌Match1-T1中,菌落PCR 檢測后,選取陽性菌落搖菌后提取質粒。利用檢測引物NOSF/M13F-47(表1)對載體質粒進行PCR 擴增,對產物進行測序,將檢測無誤的質粒通過熱激法轉入EHA105 農桿菌感受態細胞中。

表1 本研究中所用的引物Table 1.Primers used in this research.
1.3 T0 代陽性植株的獲得
通過農桿菌介導法,將CRISPR/Cas9 表達載體轉入龍粳11 的愈傷組織中,經過潮霉素抗性篩選愈傷組織,分化成T0代植株。在T0代植株成熟期時,采用CTAB 的方法提取葉片的全基因組DNA,通過引物Hyg-F/R 進行檢測,PCR 產物大小為289 bp 的植株為陽性轉基因植株,篩選出陽性苗。設計引物Seq1-F/R、Seq2-F/R 和Seq3-F/R,分別擴增GS3、GS9和Badh2基因靶點及其附近的序列,使用兼并序列解碼(DSDecode)法[31]分析測序結果。
1.4 T1 代無T-DNA 元件的純合突變植株的篩選
T1代植株成熟時期,提取葉片基因組DNA,利用引物Hyg-F/R(表1)篩選出無T-DNA 元件插入的植株。再利用測序引物Seq1-F/R、Seq2-F/R 和Seq3-F/R,對無T-DNA 元件插入的植株進行PCR擴增,測序結果通過DSDecode 方法,篩選出無T-DNA 元件的三基因純合突變的植株,將其自交繁殖至T2代。
1.5 T2 代植株農藝性狀考查
在大田環境下種植龍粳11野生型和T2代植株,成熟時期分別測量粒長、粒寬、結實率、單株產量及千粒重,并采用氣相色譜-質譜聯用技術,分析水稻籽粒中的2-乙酰-1-吡咯啉(2-AP)含量。利用SPSS 18.0 軟件對農藝性狀數據進行t檢驗。
2 結果與分析
2.1 水稻GS3、GS9 和Badh2 敲除靶點設計
GS3基因位于第3 染色體,有5 個外顯子,編碼232 個氨基酸。GS9基因位于第9 染色體,有4個外顯子,編碼345 個氨基酸。Badh2基因位于第8 染色體上,有15 個外顯子,編碼503 個氨基酸。GS3、GS9和Badh2基因CDS 序列均與日本晴相同,本研究分別在GS3第1 外顯子、GS9第1 外顯子、Badh2第2 外顯子處設計1 對gRNA 靶點接引物,分別對GS3、GS9、Badh2基因進行編輯(圖1)。
2.2 GS3、GS9 和Badh2 基因表達載體的構建
根據“金門”克隆法將帶有3 個靶點的gRNA 表達盒連接到pYLCRISPR/Cas9Pubi-H 載體骨架上,構建好的載體即為 pYLCRISPR/Cas9-GS3/GS9/Badh2-gRNA 載體(圖2)。
2.3 T0 代轉基因陽性苗檢測及突變情況
16 株T0代再生苗中有8 株為陽性苗,陽性率為50%(圖3)。利用測序引物Seq1-F/R、Seq2-F/R和Seq3-F/R 擴增8 株陽性植株的靶點區域序列,并進行PCR 擴增,測序比對后,8 株陽性植株中有4株發生了突變,突變率為50%。
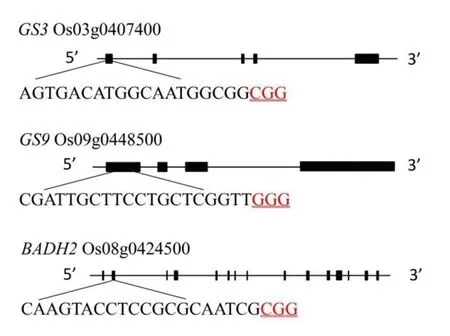
圖1 GS3、GS9 和Badh2 基因結構和靶點位置Fig.1.Gene structure and target site ofGS3,GS9 and Badh2.
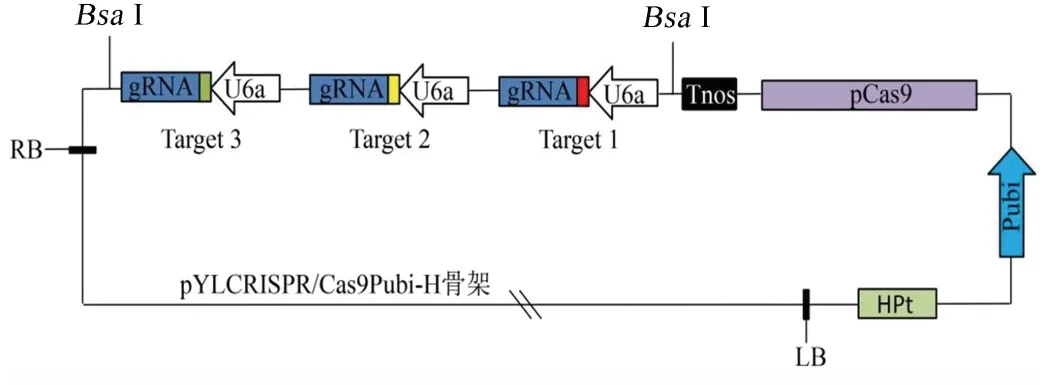
圖2 pYLCRISPR/Cas9-GS3/GS9/Badh2-gRNA 載體Fig.2.pYLCRISPR/Cas9-GS3/GS9/Badh2-gRNA vector.
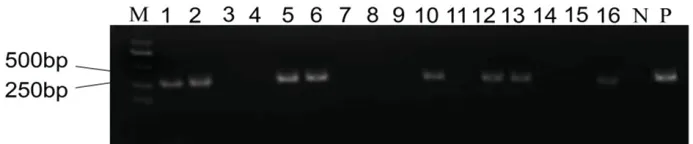
圖3 T0 代植株轉基因檢測Fig.3.Transgenic detection of T0 generation plants.
2.4 T1代無T-DNA 元件三基因純合突變植株的篩選
將三基因突變植株自交繁殖獲得T1代植株,于成熟期提取葉片基因組DNA,利用引物Hyg-F/R篩選出無T-DNA 元件插入的植株。再利用引物Seq1-F/R、Seq2-F/R 和Seq3-F/R 進行擴增,通過DSDecode 方法分析測序結果,篩選出 2 株無T-DNA 元件的三基因純合突變植株(55-2、58-2),分析GS3、GS9、Badh2基因突變的具體情況(圖4)。
2.5 T2 代植株農藝性狀分析及香味鑒定
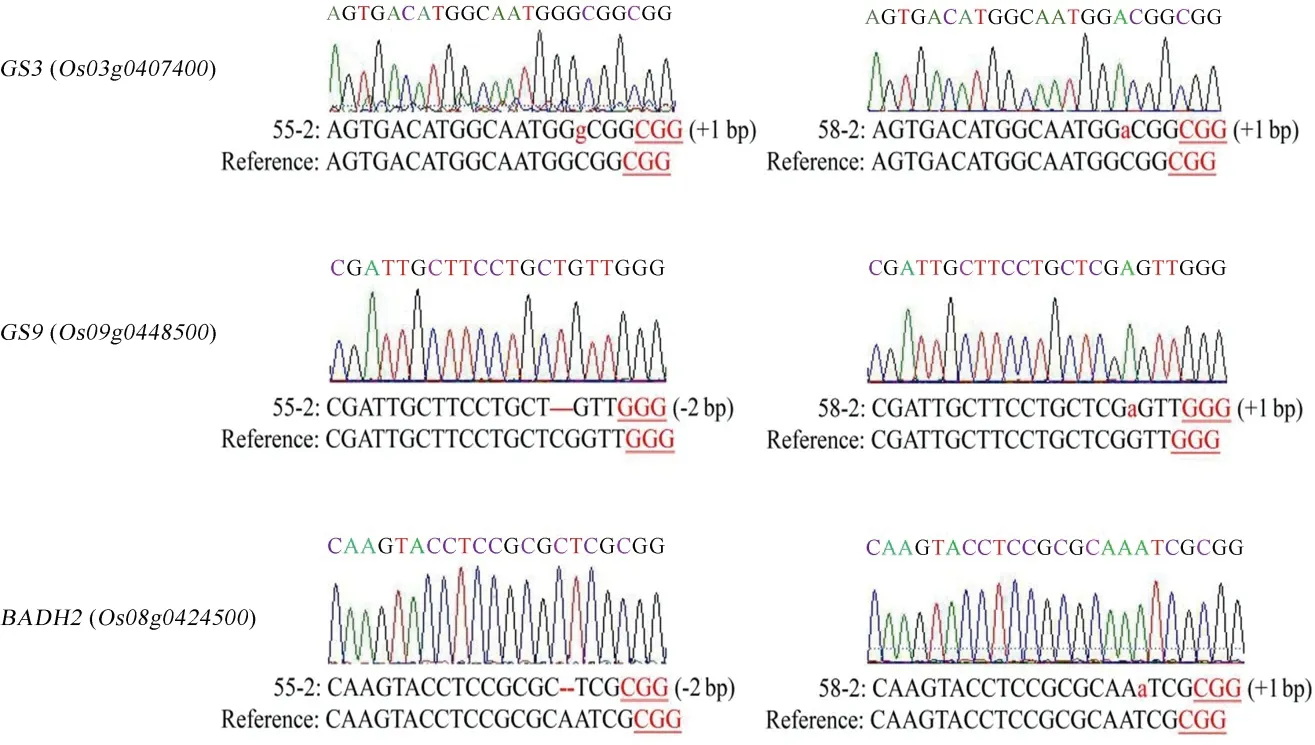
圖4 三基因純合突變植株測序結果Fig.4.Sequencing result of homozygous plants with triple gene mutation.
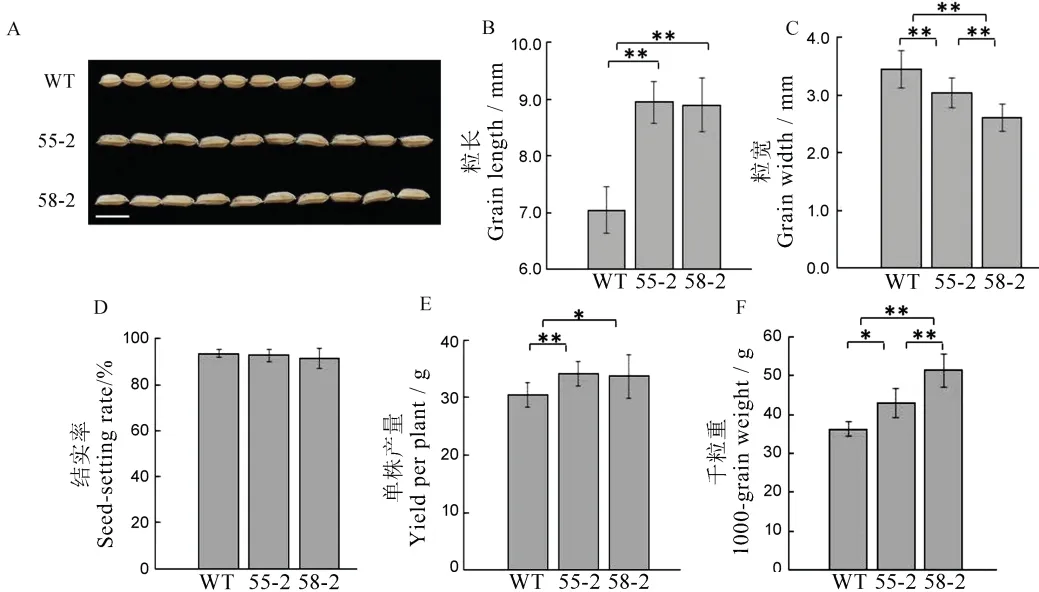
圖5 T2 代植株農藝性狀考查Fig.5.Agronomic traits of T2 generation plants.
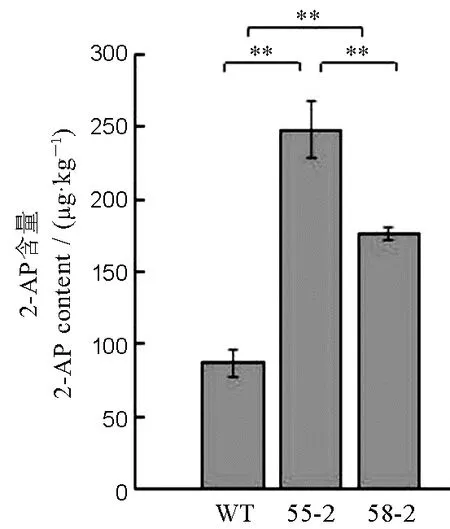
圖6 龍粳11 及突變體中香味物質2-AP 的含量Fig.6.2-acetyl-1-pyrroline(2-AP) content of Longjing 11 and mutant plants.
將野生型龍粳11 與55-2 株系、58-2 株系的農藝性狀進行比較(圖5),發現55-2 株系和58-2 株系的粒長比野生型植株分別增加27.01%、26.43%,都達到了極顯著的水平,粒寬較野生型植株降低13.4%、24.6%,結實率沒有顯著變化,單株產量分別增加10.82%、12.11%,千粒重分別增加18.34%、41.36%,58-2 株系千粒重增加比例顯著(圖5)。研究表明,通過CRISPR/Cas9 技術同時對GS3、GS9基因進行編輯,高效地將圓粒水稻變長,在育種方面加速了新品系的創制。
采用氣相色譜-質譜聯用技術,對野生型龍粳11、突變體55-2、58-2 株系中籽粒的2-AP 含量進行分析(圖6)。野生型龍粳11 中2-AP 含量約87.13 μg/kg,與龍粳11 對照相比,55-2 株系中香味物質2-AP 含量增加至248.22 μg/kg,58-2 株系中2-AP增加至176.43 μg/kg。55-2 株系和58-2 株系分別在Badh2基因第2 外顯子處缺失2 bp、插入1 bp,均產生移碼突變。結果表明,通過對Badh2基因進行編輯,能有效創制香型水稻種質新資源。
3 討論
CRISPR/Cas9 技術與傳統育種相比,因其具有操作簡單、編輯效率高和成本低等優點,正廣泛應用于水稻的種質改良中[32-35],極大地縮短了培育高產優質水稻品種的周期。本研究以粒型負調控基因GS3、GS9和香味負調控基因Badh2為目標基因,構建 pYLCRISPR/Cas9-GS3/GS9/Badh2-gRNA 載體,對GS3、GS9、Badh2基因進行敲除,篩選出兩個無T-DNA 元件的三基因純合突變株系。
GS3和GS9是粒長的負調控因子。Nan 等[36]利用具有gs3等位基因的供體GKBR 與黑龍江省優良栽培品種空育131 雜交后,以空育131 為輪回母本,經過回交3 代改善了空育131 的GS3位點,籽長增加12.05%,千粒重和單株總粒數提高,進而產量增加,但改良周期過長,效率低。通過CRISPR/Cas9 技術可以極大地縮短育種年限,還大幅度提高粒長。Li 等[23]利用CRISPR/Cas9 技術對GS3基因進行編輯,破壞GS3正常表達,粒長增加20%。Zhao 等[14]使用CRISPR/Cas9 技術,在GS9第1 外顯子處插入1 bp,使粒長增加5.5%。本研究通過將兩個基因連入同一載體,同時對GS3和GS9進行編輯。通過對GS3和GS9基因進行編輯,粒長與野生型相比差異達到極顯著水平,增幅為26.43%~27.01%,增幅大于通過CRISPR/Cas9 技術分別敲除GS3和GS9的效果,單株產量增加10.82%~12.11%,結實率沒有顯著變化,千粒重增加 18.34%~41.36%。研究結果表明,利用CRISPR/Cas9 技術可以培育出長粒型且高產的品種,加快長粒種質資源的創制。
傳統的香稻育種主要通過雜交和回交選育,但因其后代分離不穩定、試驗周期漫長、抗逆性較弱、產量較低等因素,使得培育出穩定遺傳的優良香稻品種較為困難。本研究對Badh2基因進行編輯后,通過對籽粒的2-AP 含量進行分析,Badh2基因功能缺失后香味物質2-AP 含量由87.13 μg/kg 增加至176.43~248.22 μg/kg,香味物質2-AP 顯著提高。孫慧宇等[27]通過CRISPR/Cas9 技術對Badh2基因進行編輯,在香味檢測時采用咀嚼法和氫氧化鉀浸泡法,主觀干擾性大。本研究采用氣相色譜-質譜聯用技術,定量分析水稻籽粒中的2-AP 含量,結果更客觀準確。
綜上所述,本研究以龍粳11 為材料,通過CRISPR/Cas9 技術對GS3、GS9和Badh2基因進行定點敲除并獲得了無T-DNA 元件的三基因純合突變體材料,使龍粳11 粒型變長,單株產量增加,千粒重增加,同時香味物質2-AP 含量增加,創制出長粒香型水稻品種,為培育產量與品質均提高的水稻品種提供了理論和材料基礎,加速了高產優質香稻品種的選育進程。
謝辭:中國科學院東北地理與農業生態研究所對本研究給予了極大幫助,在此表示感謝。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
