牛磺酸對肉雞生長性能、肉品質及肌肉抗氧化功能的影響
2020-09-21 08:15:32韓紅麗張婧菲沈明明何家樂楊培萱
食品科學 2020年17期
關鍵詞:實驗
韓紅麗,張婧菲,沈明明,何家樂,劉 佳,楊培萱,王 恬
(南京農業大學動物科技學院,江蘇 南京 210095)
牛磺酸(taurine,TAU)是一種含硫的β氨基酸,以高濃度存在于絕大部分動物細胞中[1]。在1827年,研究者首次從牛膽汁中分離出TAU[2]。TAU已廣泛應用于動物生產中,具有促進動物生長、提高機體抗氧化能力和免疫力等多種生物學功能。日糧添加適量TAU能改善肉雞的生長性能、免疫功能、總抗氧化能力(total antioxidant capacity,T-AOC)和脂質代謝[3]。飼料中添加適宜劑量的TAU,能提高鯉魚腸道超氧化物歧化酶(superoxide dismutase,SOD)、過氧化氫酶(catalase,CAT)和谷胱甘肽過氧化物酶(glutathione peroxidase,GPx)的活性,抑制丙二醛(malondiadehyde,MDA)的生成,增強鯉魚的抗氧化能力[4]。此外,TAU的前體物質(半胱氨酸、半胱胺、亞牛磺酸等)也具有抗氧化活性,可有效清除活性氧自由基,抑制脂質過氧化反應[5]。
近幾十年來,肉雞產業發展迅速,全球肉雞消費量僅次于豬肉,但是由于肉雞肌肉中富含多不飽和脂肪酸,易氧化酸敗,引發肌肉變質。現代養殖環境中的各種應激(如熱應激、免疫應激、屠宰應激等)均會加速機體脂質氧化,引起肌肉蛋白質和脂質過氧化,影響肌肉的肉品質(如肉品失色、滴水損失增加等),從而對肉品質產生不良影響。氧化應激通過改變蛋白質結合氫和毛細管結合水分子的能力,影響肌肉的保水性[6]。因此,通過提高肌肉的抗氧化能力,能有效改善肌肉品質。然而,目前關于TAU在肉雞方面的研究多集中于從抗氧化酶活性的角度來探討TAU的抗氧化作用,但有關TAU對肉雞肌肉抗氧化功能及相關基因mRNA表達水平對肌肉品質的影響研究仍十分缺乏。因此,本實驗旨在研究日糧中添加不同水平的TAU對肉雞生長性能、肌肉品質、胸肌抗氧化功能的影響,探討肌肉抗氧化與肌肉品質之間的關系,同時篩選出TAU在肉雞日糧中的適宜添加水平,為生產中以營養調控的方式改善肉雞的肌肉品質提供新的思路和方法,同時也為TAU在提高肉品質健康方面的研究提供參考。
1 材料與方法
1.1 材料與試劑
TAU(純度>98.5%)購自江陰華昌食品添加劑有限公司;‘艾拔益加’(Arbor Acres,AA)肉雞購自安徽和威集團;DEPC水購自上海碧云天生物技術有限公司。T-AOC、GPx、SOD、CAT和MDA試劑盒均購自南京建成生物工程研究所;TRIzol試劑、反轉錄試劑盒、聚合酶鏈式反應(polymerase chain reaction,PCR)試劑盒均購自寶生物工程(大連)有限公司。
1.2 儀器與設備
Nanodrop ND-2000c分光光度計、Muliskan Go全波長酶標儀、Q u a n t S t u d i o 5 熒光定量P C R 儀美 國T h e r m o S c i e n t i f i c 公 司;C R-4 0 0 型 色 度儀日 本K o n i c a M i n o l t a 公 司;H I 9 0 2 5 型p H 計意大利Hanna公司;C-LM3B嫩度儀 東北農業大學;5804R離心機 德國Eppendorf公司;Secura125-1CN電子天平 賽多利斯科學儀器(北京)有限公司。
1.3 方法
1.3.1 實驗分組及設計
實驗采用隨機單因素試驗進行分組設計,選擇1 日齡初始體質量相近的健康AA肉雞384 只,隨機分為4 組,每組12 個重復,每個重復8 只肉雞。對照組(CON組)肉雞飼喂玉米-豆粕型基礎日糧,實驗組肉雞分別在基礎日糧的基礎上添加0.25%(TAU1組)、0.5%(TAU2組)、0.75%(TAU3組)的TAU。實驗周期為42 d,分為前期(1~21 d)和后期(22~42 d)。實驗肉雞采用3 層疊籠飼養。在實驗期間,自由采食和飲水,每日清糞,保持舍內的通風換氣。實驗過程中,嚴格按照常規程序進行免疫處理。基礎日糧參考National Research Council(1994)《肉雞飼養標準》完成配制[3],配方及營養水平見表1。
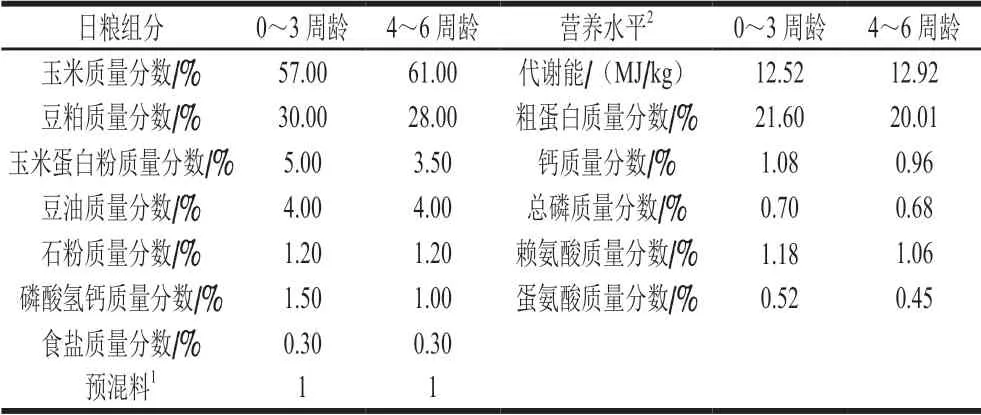
表 1 日糧配方及營養水平Table 1 Ingredients and nutrient contents of experimental diets used in this study
1.3.2 樣品采集
于實驗第42天,從每個重復中隨機選1 只體質量接近于該重復平均體質量的肉雞,此前禁食12 h。于頸動脈處放血,待其死亡后立即解剖,在每只肉雞的同一部位取適量胸肌樣品,迅速置于-80 ℃冰箱保存備用,同時取另一側胸肌用于測定肉品質。
1.3.3 指標測定
1.3.3.1 生長性能測定
于實驗第21天和第42天分別禁食12 h后空腹稱體質量,以每個重復為單位記錄采食量,并以此為基礎計算平均日采食量、平均日增體質量和料重比(平均日采食量與平均日增體質量之比)[3]。
1.3.3.2 肉品質測定
pH值測定:于實驗第42天肉雞屠宰后,迅速取下一側胸肌置于冰上,45 min時測定1 次肉樣的pH值,然后置于4 ℃冰箱,24 h后再次測定肉樣的pH值。每個樣品于不同位置測量3 次,結果取其平均值[7]。
肉色測定:胸肌樣品在4 ℃存放24 h后取出,使用色度儀測定樣品的明度(L*)、紅度(a*)和黃度(b*),每個樣品于3 個不同位置進行測定,結果取其平均值[8]。
滴水損失率測定:于實驗第42天,肉雞屠宰后,將肌肉修整為3 cm×2 cm×1 cm的肉塊,稱質量(m1/g),然后用細鐵絲鉤住肉樣,使肌纖維垂直向下,懸吊于4 ℃冰箱,24 h后取出肉樣后稱質量(m2/g),按式(1)計算24 h滴水損失率,再過24 h后取出肉樣稱質量(m3/g),按式(2)計算48 h滴水損失率[9]。
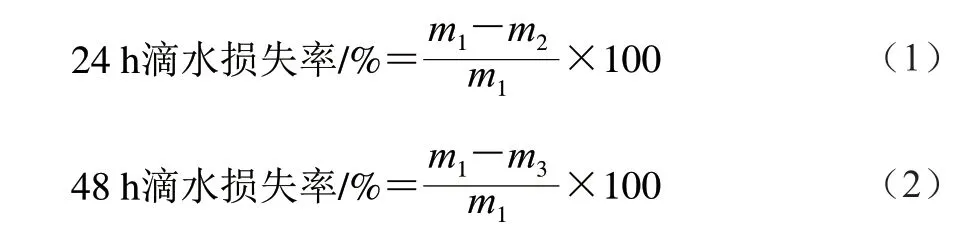
蒸煮損失率測定:于實驗第42天肉雞屠宰后,用取樣器切取肉塊,去除結締組織,用塑料袋包裝,在0~4 ℃冰箱熟化24 h,稱質量(m1/g)。把肌肉放到水浴鍋中加熱,加熱到肉中心溫度為70 ℃,水浴溫度在75~80 ℃,取出肉樣冷卻后稱質量(m2/g)[9]。按式(3)計算蒸煮損失率。

剪切力測定:將測定蒸煮損失率后的樣品順著肌纖維方向切為截面為1 cm×1 cm的3 個肉條,再以垂直肌纖維方向用嫩度儀對其進行剪切,每個肉條剪切1 次,結果取3 次剪切力讀數的平均值[8]。
1.3.3.3 肌肉抗氧化能力測定
稱取約0.5 g的肌肉樣品,加入4.5 mL生理鹽水(質量分數0.75%的NaCl溶液),在冰浴條件下充分勻漿,然后4 ℃條件下4 000×g離心10 min,吸取分裝上清液,置于-20 ℃條件下儲存待測[10]。測定肌肉(1 mg蛋白中)T-AOC、GPx活力、SOD活力、CAT活力和MDA含量。
1.3.3.4 肌肉抗氧化相關基因mRNA的表達測定
總RNA的提取:提前將所用試劑預冷,取出-80 ℃條件下保存的肌肉樣品,切取約50 mg肌肉,采用TRIzol試劑從肌肉中提取總RNA,使用分光光度計測定OD260nm和OD280nm,通過觀察兩者的比值即OD260nm/OD280nm來評估總RNA的濃度和純度,比值介于1.8~2.1表示RNA的純度較高、未被污染。
RNA反轉錄:將不同樣品的RNA濃度用DEPC水統一稀釋成500 ng/μL,利用反轉錄試劑盒合成cDNA。反轉錄的反應條件為:37 ℃下反應15 min,85 ℃下變形15 s,最后降溫至4 ℃。
熒光定量PCR:利用實時熒光定量PCR試劑盒進行。PCR程序為:95 ℃、30 s,95 ℃、5 s,60 ℃、31 s,40 個循環。目的基因的相對表達量以β-Actin作為內參基因,采用比較2-ΔΔCt法進行計算。所有基因序列均從GenBank中獲取,引物序列見表2,由生工生物工程(上海)股份有限公司合成。

表 2 目的基因及內參基因引物信息Table 2 Primer sequences information for target and reference genes
1.4 數據處理與分析
實驗數據采用Excel 2010軟件進行初步匯總整理后,采用SPSS Statistics 22軟件進行統計,采用單因素方差分析進行差異顯著性檢驗,并采用Duncan’s法進行多重比較,結果用平均值±標準誤差表示,當P<0.05時表示差異顯著。
2 結果與分析
2.1 添加不同水平的TAU對肉雞生長性能的影響
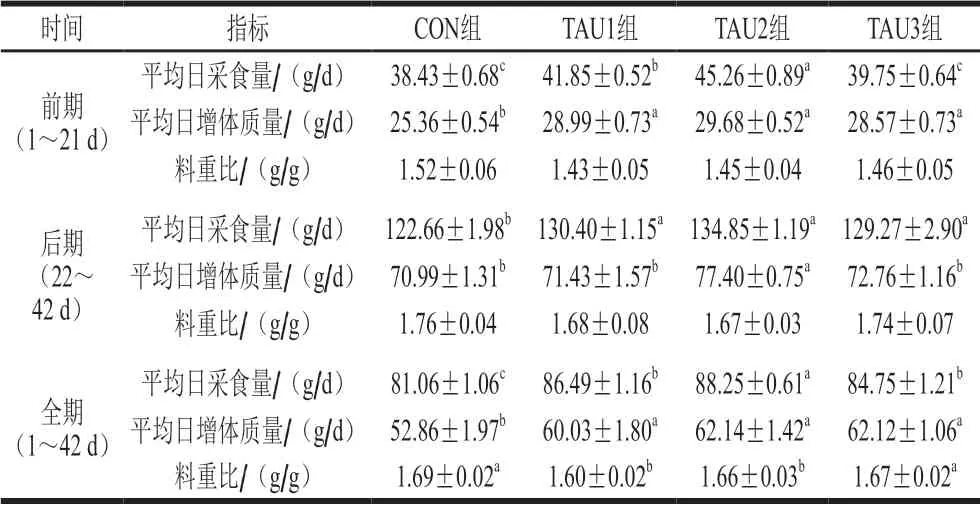
表 3 添加不同水平TAU對肉雞生長性能的影響Table 3 Effects of dietary TAU supplementation at different levels on growth performance of broilers
由表3可知,實驗前期,與CON組相比,各TAU處理組均能顯著提高肉雞的平均日增體質量(P<0.05)。其中,TAU1和TAU2組平均日采食量較CON組分別顯著提高了8.90%和17.77%。實驗后期,與CON組相比,各TAU處理組均能顯著提高肉雞的平均日采食量(P<0.05),其中TAU2組平均日采食量較CON組相比提高了9.94%。從實驗全期來看,與CON組相比,各TAU處理組均能顯著提高肉雞的平均日增體質量和平均日采食量(P<0.05)。其中,TAU1和TAU2組的料重比相較CON組分別顯著降低了5.33%和1.78%(P<0.05)。
2.2 添加不同水平的TAU對肉雞胸肌肉品質的影響
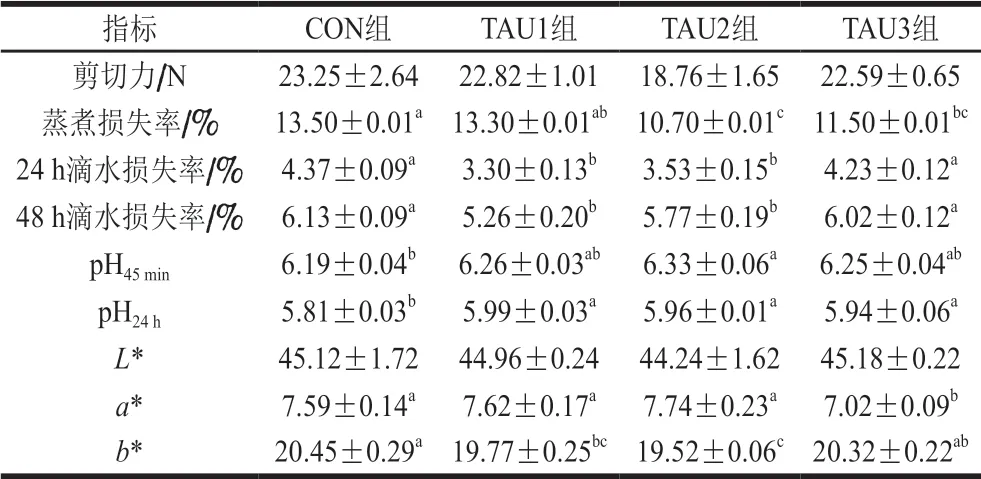
表 4 添加不同水平的TAU對肉雞胸肌肉品質的影響Table 4 Effects of dietary TAU supplementation at different levels on breast muscle meat quality of broilers
由表4可知,與CON組相比,TAU2組胸肌的滴水損失率和蒸煮損失率顯著降低(P<0.05)。各TAU處理組pH24h均顯著高于CON組(P<0.05),但TAU處理組之間差異不顯著(P>0.05)。就胸肌的b*值而言,TAU1和TAU2組顯著低于CON組(P<0.05)。各組間的胸肌剪切力和L*值差異均不顯著(P>0.05)。
2.3 添加不同水平的TAU對肉雞胸肌肉抗氧化指標的影響

表 5 添加不同水平的TAU對肉雞胸肌肉抗氧化指標的影響Table 5 Effects of dietary TAU supplementation at different levels on the antioxidant capacity of breast muscle in broilers
由表5可知,與CON組相比,各TAU處理組均能顯著提高肉雞胸肌中GPx活力(P<0.05),降低MDA含量(P<0.05)。TAU1組SOD活力顯著高于CON組(P<0.05)。
2.4 添加不同水平的TAU對肉雞胸肌抗氧化相關基因mRNA相對表達量的影響
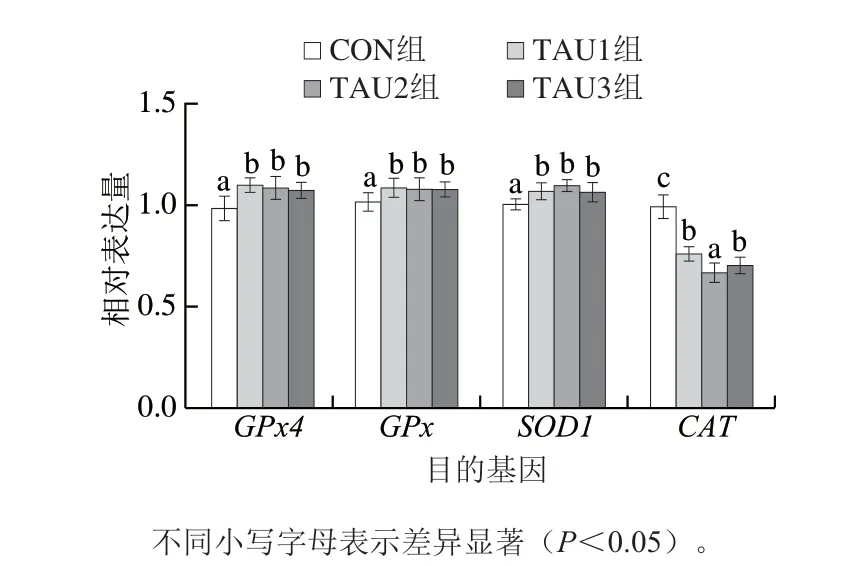
圖 1 TAU對肉雞胸肌抗氧化相關基因mRNA相對表達量的影響Fig. 1 Effects of dietary TAU supplementation at different levels on antioxidant enzyme mRNA expression in broilers
由圖1可知,與CON組相比,各TAU處理組均能顯著提高肉雞胸肌中GPx4、GPx及SOD1mRNA相對表達量(P<0.05),但TAU1、TAU2和TAU3各處理組之間差異不顯著(P>0.05)。與CON組相比,各TAU處理組均能顯著降低CAT的相對表達量(P<0.05)。
3 討 論
3.1 TAU添加水平對肉雞生長性能的影響
目前,關于TAU對動物生長性能的影響已有一定的研究。李麗娟等[10]的研究結果表明,在生長前期日糧中添加TAU可不同程度地提高肉雛雞的平均日增體質量,降低料重比。黃春喜等[11]研究發現,日糧添加TAU可以顯著降低肉仔雞早期(1~7 d)的料重比。Liu Yue等[12]發現飼糧添加TAU能提高仔豬的生長性能,促進腸道發育。本實驗結果表明,從全期來看,日糧添加TAU能顯著提高肉雞的平均日增體質量和平均日采食量,其中添加0.25%和0.5%的TAU能顯著降低肉雞的料重比,這與上述研究結果基本一致。TAU的促生長作用可能與它的抗氧化作用有關,TAU可緩解活性氧對機體脂質的氧化損傷,提高動物體對脂肪的利用,進而促進動物的生長。也有研究表明,在植物蛋白的飼糧中添加TAU對斑馬魚的生長性能沒有顯著影響[13]。因此推測,造成這種差異的原因可能與日糧類型、物種、生長階段、飼養環境和TAU的添加量等因素有關。
3.2 TAU添加水平對肉雞肉品質的影響
影響肌肉品質的因素有很多,包括pH值、系水力、剪切力和肉色等。剪切力是評價肌肉嫩度的重要指標,剪切力越小,說明肌肉的嫩度越好[14]。另外,肌原纖維的狀態、肌肉中結締組織的含量、結締組織中膠原蛋白的種類和交聯程度均是影響肉嫩度的重要因素[15]。本實驗結果表明,在日糧中添加適量的TAU能降低肌肉的剪切力、提高肌肉的嫩度、降低肌肉的滴水損失,其中以0.25%和0.5%的添加量效果最佳,這與臧素敏等[16]的研究結果一致。鈣蛋白酶的氧化可以抑制蛋白質降解,而相關研究表明,動物體內抗氧化防御系統的提高會影響鈣蛋白酶活性、蛋白質水解,從而改善肌肉的系水力和嫩度[17],表明TAU的添加可能是通過上述途徑提高肌肉的系水力和嫩度。
pH值是影響肌肉系水力的重要因素,肌肉pH值的升高在一定程度上會減少蛋白質的變性,使肌原纖維蛋白質偏離其等電點,減弱蛋白質之間的相互作用,進而降低滴水損失和蒸煮損失,提高肌肉的系水力[9]。此外,pH值的變化能夠在一定程度上反映肉雞屠宰后機體糖原酵解的速度。肉雞屠宰后,肌肉中乳酸菌對糖原進行無氧酵解從而產生乳酸,乳酸積累會導致肌肉酸度增加,pH值下降[18]。本實驗發現,肉雞屠宰后pH45min和pH24h實驗組均高于CON組,其中以0.5%的TAU添加量效果最佳。魏源等[19]的研究結果表明,服用TAU能降低運動大鼠乳酸含量,由此推測TAU可能通過減少肌肉中乳酸的積累來提高pH值,延緩肌肉變質的速度,改善肉雞的肌肉品質。但Huang Chunxi等[20]研究結果表明日糧中添加不同水平的TAU對肌肉pH值和肉色均未產生顯著影響,這種差異的原因可能與物種、飼料組成以及添加劑量等因素有關。
肉色是肌肉外觀評定的重要指標,是肌肉的生理學、生物化學和微生物學變化的外部體現[21]。實驗結果顯示,在日糧中添加TAU能顯著降低肉雞胸肌的b*值,b*值表示肉樣的黃度,b*值越低肉色越好。曾得壽等[22]發現在肉仔雞日糧中添加0.2%TAU可以顯著提高肉雞胸肌的肉色。肉的色澤主要受肌紅蛋白含量的影響,肌紅蛋白含量越多,肉色越深。研究表明肌肉組織內的氧化會導致高鐵肌紅蛋白的生成,使得肉的表觀色澤變差,其機制涉及不飽和脂肪酸的過氧化,而抗氧化劑的加入可以保護脂質不被氧化,并穩定氧肌紅蛋白的含量[23]。由此推測添加適量的TAU可能通過其抗氧化作用抑制脂質過氧化,在一定程度上穩定氧肌紅蛋白的含量,進而改善肌肉胸肌的肉色[24]。
3.3 TAU添加水平對肉雞肌肉抗氧化功能的影響
TAU具有多種生物學功能,其中抗氧化功能是TAU最為重要的生物學功能之一。肉品質與抗氧化屬性密切相關。脂質過氧化是導致肉品質惡化的重要原因之一,它可以降低肌肉的營養價值、影響風味和質地、改變肌肉的外觀,因此提高肌肉的抗氧化功能,對于改善肌肉品質具有重要的意義。MDA是脂質過氧化代謝的代表性產物之一,其含量的高低直接反映脂質過氧化損傷的程度。在正常生理狀態下,體內的抗氧化酶(如GPx、SOD)可發揮協同作用,中和超氧化物陰離子自由基和過氧化氫,減輕氧自由基鏈式反應對機體的損害[25]。相關研究已經證明,在日糧中添加TAU能通過提高機體抗氧化酶的活性,降低MDA的含量,從而提高機體的抗氧化功能[26-27]。本實驗結果表明,在日糧中添加TAU能提高GPx和SOD活力,降低MDA的含量,與上述研究結果一致。研究表明,抑制肌肉的脂質過氧化,可減弱蛋白質降解,降低肌原纖維間的水分儲備,從而降低肌肉的系水力[28]。實驗所觀察到的胸肌肌肉蒸煮損失的減少可能是因為肌肉的系水力與蛋白質水解存在一定的關系[29],TAU的添加降低了肌肉蛋白質和脂質的過氧化,進而提高肌肉的保水性。GPx4是機體內酶類抗氧化系統中的重要成員,主要作用是清除膜或脂蛋白中的脂質過氧化物。有研究表明,細胞膜的完整性與滴水損失有關,由于肌肉細胞膜中含有大量不飽和脂肪酸和磷脂,易受自由基的攻擊而誘發脂質過氧化,從而破壞細胞膜的結構,增加雞肉水分的滲出。由此推測,TAU提高肌肉系水力可能是通過上調肌肉中GPx4基因的表達,降低肌肉中MDA的含量來實現的[30]。同時,使蒸煮損失減少,細胞失水皺縮減少,從而改善肌肉的色澤。此外,機體內脂質過氧化產物含量的減少會減緩氧合肌紅蛋白轉化為高鐵肌紅蛋白,進而改善肌肉顏色[31],最終緩解肌肉氧化變質,提高肌肉嫩度,但具體機制有待進一步研究。本實驗發現TAU可以上調肌肉中GPx和SOD1基因的表達,這個結果表明TAU可能通過調控抗氧化酶的基因表達,從分子水平來提高肉雞胸肌的抗氧化能力,這也與TAU能提高GPx和SOD活力結果一致。
綜上,在日糧中添加TAU可以提高肉雞的生長性能、改善肉雞胸肌的肉品質和抗氧化功能。添加TAU可能與其抑制脂質過氧化、提高胸肌抗氧化基因mRNA的表達水平有關。本實驗條件下,肉雞日糧中TAU添加量以0.5%效果最佳,表明TAU對于改善肉品質健康具有重要的作用,為其在食品科學領域的研究提供了參考。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
