貴州特有植物黔中杜鵑種群結構及其動態分析
2020-09-28 08:15:16姜順邦袁叢軍余德會戴曉勇龍海燕
浙江林業科技 2020年4期
姜順邦 ,袁叢軍,余德會,戴曉勇,楊 冰,李 鶴,龍海燕
(1.貴州省林業學校,貴州 貴陽 550201;2.貴州省林業科學研究院,貴州 貴陽 550005;3.貴州雷公山國家級自然保護區管理局,貴州 雷山 557100)
種群是物種進化的基本單位,種群結構是種群最基本的特征,它既可反映種群內不同大小個體數量的分布和配置狀況,也能反映種群的數量動態及其未來發展與演變趨勢[1]。種群生命表與其構建的存活曲線以及生存分析理論不僅可以反映種群目前的存活狀態,而且可以反映植物與環境之間的適合度[2],結合種群動態量化分析和時間序列預測還可以闡明種群的受干擾狀況及未來的發展趨勢。因此,研究植物種群結構與動態有助于了解種群的生活史和物種的更新動態,對野生植物的保護及開發利用具有重要意義。
黔中杜鵑Rhododendron feddei為杜鵑花科Ericaceae 杜鵑花屬Rhododendron常綠灌木或小喬木,生長于海拔1 120~ 1 740 m 的叢林中,為貴州特有種[3],該種雖然發現較早,但在《貴州植物志》和《中國植物志》中都沒有收錄,而是在《FOC》中才有記載[4]。經過多年的尋找,我們在其模式產地的貴定縣云霧山一帶再次發現該種,在雷公山保護區也發現了該種,有關報道在貴州百里杜鵑(百里杜鵑國家級森林公園)發現的該種經過查證后并不存在[4]。由于杜鵑花科植物具有很高的利用價值和科研價值,近年來,關于杜鵑花科植物種群結構的研究比較多,如莫羅堅對東莞蓮花山毛棉杜鵑花Rh.moulmainense種群結構進行了動態分析,認為毛棉杜鵑花種群徑級分布呈倒“J”型,屬于增長型種群[5];鄧賢蘭對井岡山杜鵑Rh.jinggangshanicum種群與群落特征進行了研究,認為井岡山杜鵑種群徑級結構為金字塔型,生命期望值基本隨徑級增加而呈遞減趨勢,存活曲線為近直線型,種群屬于增長型種群[6];蔡鑫對華頂杜鵑Rh.huadingense的種群結構和種間聯結進行了研究,認為華頂杜鵑種群呈衰退趨勢,幼苗和幼樹儲備不足[7]。這些學者的研究在很大程度上揭示了許多杜鵑花科植物的種群結構及其數量動態變化特征。黔中杜鵑作為貴州特有植物,種群數量少,具有一定的研究價值,然而,目前有關該物種的研究尚未見報道。本文以雷公山國家級自然保護區內仙女塘的黔中杜鵑種群為研究對象,對其種群結構及數量動態變化進行了分析,揭示該區黔中杜鵑種群數量變化動態特征、種群動態變化趨勢,以期為該種的保護和進一步研究提供重要理論支撐。
1 材料與方法
1.1 研究區概況
研究區位于貴州雷公山國家級保護區仙女塘,地理坐標為108°11′37.98″ E,26°22′18.87″ N,屬于中亞熱帶季風氣候,年均溫在13.9~ 15.2℃,極端最低氣溫為-6.2℃,最高氣溫為21.7℃,≥10℃年積溫在4 098.0~ 4 531.2℃,年日照時數為1 513.2 h,無霜期在281~ 290 d,年均降水量在1 323.7~ 1 577.2 mm,雨期4-9 月降水量在1 065.1~ 1 249.7 mm,占全年降水的80.0%。
調查群落位置均在中坡,土壤類型主要為黃壤,群落為常綠落葉闊葉混交林,其中,喬木層樹種主要有白辛樹Pterostyrax psilophyllus,檫木Sassafras tzumu,楓香樹Liquidambar formosana,云南榿葉樹Clethra cavaleriei,花楸樹Sorbus pohuashanensis,交讓木Daphniphyllum macropodum,老鼠矢Symplocos stellaris,毛棉杜鵑花,青岡Cyclobalanopsis glauca,樹參Dendropanax dentiger,水青岡Fagus longipetiolata,甜櫧Castanopsis eyrei,銀木荷Schima argentea,云貴鵝耳櫪Carpinus pubescens,錐栗Castanea henryi等;灌木層植物有川桂Cinnamomum wilsonii,川楊桐Adinandra bockiana,吊鐘花Enkianthus quinqueflorus,光葉山礬Symplocos sumuntia,黃丹木姜子Litsea elongata,凱里杜鵑Rh.kailiense,柃木Eurya japonica,毛果杜鵑Rh.seniavinii,毛棉杜鵑花,石木姜子Litsea elongatavar.faberi,茶莢蒾Viburnum setigerum,狹葉衛矛Euonymus stenophyllus,紫金牛Ardisia japonica等;草本植物分布較少。
1.2 研究方法
1.2.1 樣地設置與調查方法 在野外實地踏查的基礎上,選取黔中杜鵑天然群落為研究對象,于2019 年5 月進行調查,分別設置具代表性的樣地3 個(表1),統一設在在中坡位置,每個樣地大小為20 m×30 m,總樣地面積為1 800 m2,樣地間的距離大于100 m,每個樣地以10 m 為間隔設置成6 個10 m×10 m 的樣方,共18 個樣方。詳細記錄每個樣地的海拔、坡向、坡度等立地因子;對樣地內喬木樹種進行每木調查,記錄各樹種的株高、地徑(胸徑)、冠幅、枝下高等指標;同時,在每個樣方的左下角設置1 個2 m×2 m 樣方進行灌木層調查、1 個1 m×1 m 樣方進行草本層調查。
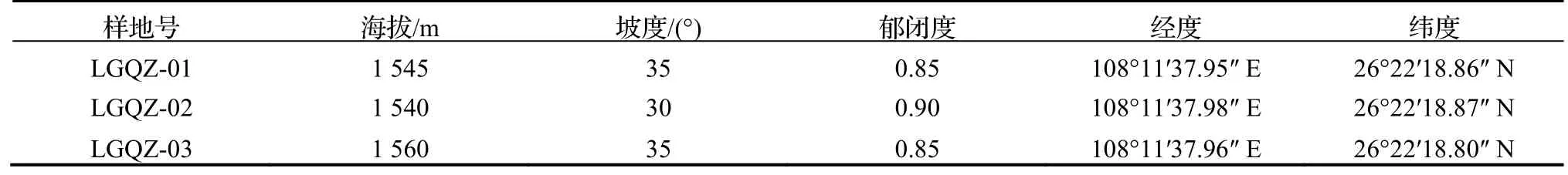
表1 群落樣地基本情況Table 1 Basic information of sample plots
1.2.2 徑級劃分 為避免破壞黔中杜鵑野生植物資源,研究采用“空間替代時間”的方法,即將林木依地徑大小分級,以立木徑級結構代替年齡結構分析種群動態,“Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ,Ⅵ,Ⅶ,Ⅷ徑級”分別對應“1,2,3,4,5,6,7,8 齡級”。關于種群大小結構立木徑級的劃分標準,許多學者提出了不同的標準[8-12]。根據該種的生物學特性,本試驗以灌叢為單株,采用植株地徑(BD)作為個體大小的指標研究其種群大小結構。徑級劃分方法:Ⅰ級(BD<2.0 cm),Ⅱ級(2.0≤BD<4.0 cm),Ⅲ級(4.0≤BD<6.0 cm),Ⅳ級(6.0≤BD<8.0 cm),Ⅴ級(8.0≤BD<10.0 cm),Ⅵ級(10.0≤BD<12.0 cm),Ⅶ級(12.0≤BD<14.0 cm),Ⅷ級(14.0≤BD<16.0 cm)。
采用陳曉德[13]的數學模型推導種群年齡結構的動態指數對黔中杜鵑種群動態進行定量描述,其計算方法為:
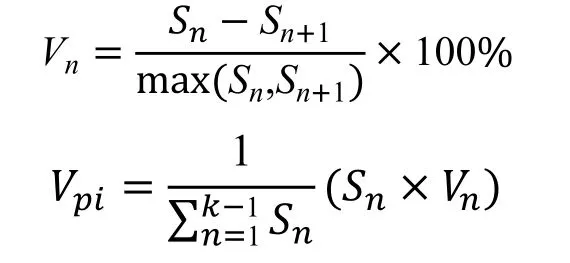
式中,Vn為種群從n級到n+1 級的個體數量變化指數;Vpi為整個種群結構的數量變化動態指數;Sn為第n個齡級種群的個體數;Sn+1為第n+1 個齡級種群的個體數。當考慮未來的外部干擾時:
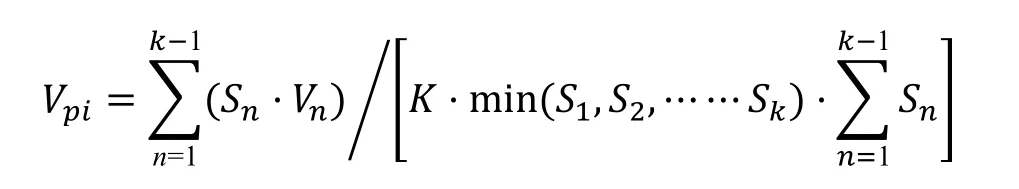
式中,K為種群齡級數量。Vpi與Vn的數值取正、負、零的意義分別反映了種群或相鄰齡級個體數量的增長、衰退及穩定的動態關系。
1.2.3 生命表編制 靜態生命表又稱特定時間生命表,主要用于木本植物種群的統計研究,其主要參數如下:a0為種群開始時的個體數量,Ax為在x齡級內的現有個體數;ax為勻滑后x齡級內的現存個體數;lnlx為在x齡級開始時的標準化存活個體數;dx為從x到x+1 齡級間隔期內的標準化死亡數;qx為從x到x+1 齡級間隔期間的死亡率;lx為從x到x+1 齡級間隔期間還存活的個體數;Tx為從x齡級到超過x齡級的個體總數;ex為進入x齡級個體的生命期望或平均期望壽命;Kx為消失率(損失度)。表中各項都是相互關聯的,可以通過實測值求得,其關系為:
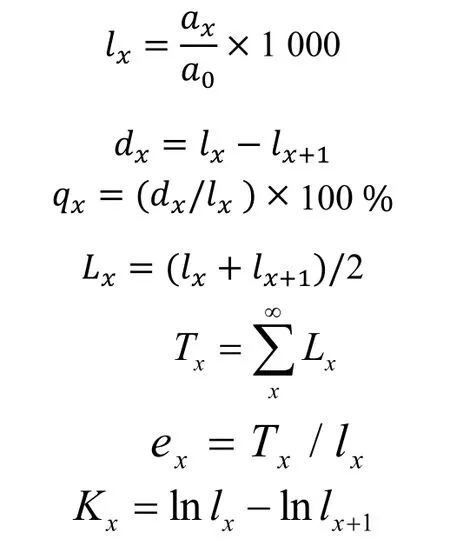
由于靜態生命表是反映了多個世代重疊的年齡動態歷程中的一個特定時間,而不是對這一種群的全部生活史的追蹤,并且調查中存在系統誤差,在生命表中會出現死亡率為負的情況,因此,本研究采用勻滑技術對數據進行處理[14]。
1.2.4 生存函數 采用生存率函數(Si)、累積死亡率函數(Fi)、死亡密度函數(fi)和危險率函數(λi)[15-18]等指標對黔中杜鵑自然種群的生存規律及其種群動態進行分析,其計算公式如下:
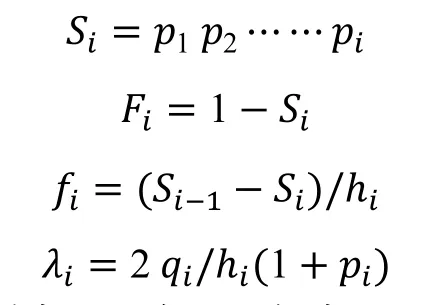
式中,hi是第i間隔期的時間長度,pi為存活率,qi為死亡頻率。
1.2.5 種群數量動態的時間序列預測 本研究采用時間序列分析中的一次移動平均法[19-20]對黔中杜鵑種群的年齡結構進行預測,其計算公式如下:
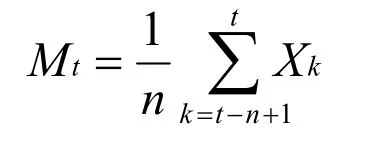
式中,n表示需要預測的時間(本研究為齡級時間),t為齡級,Xk為k齡級內的個體數量,Mt表示經過未來n個齡級時間后t齡級的種群大小。本文對未來經過2,4,6,8 和10 齡級時間后的種群各齡級的個體數量進行預測。
2 結果與分析
2.1 黔中杜鵑所在群落樹種組成
調查結果表明,在研究區域的樣地中,共有維管束植物23 科36 屬43 種,其中,蕨類植物有2 科2 屬2 種,被子植物有21 科34 屬41 種。群落結構主要分為喬木層、灌木層和草本層三個層次。構成喬木層的樹種共17種,其物種組成及重要值見表2;構成灌木層的樹種共22 種,其物種組成及重要值見表3;黔中杜鵑在喬木層中的重要值為0.298,在灌木層中的重要值為0.099,雖然黔中杜鵑在喬木層和灌木層中的重要值均為最大,但其他物種的重要值占比也相對較大,所以,黔中杜鵑群落為混交林。草本層種類及數量均較少,其種類、頻度等見表4。
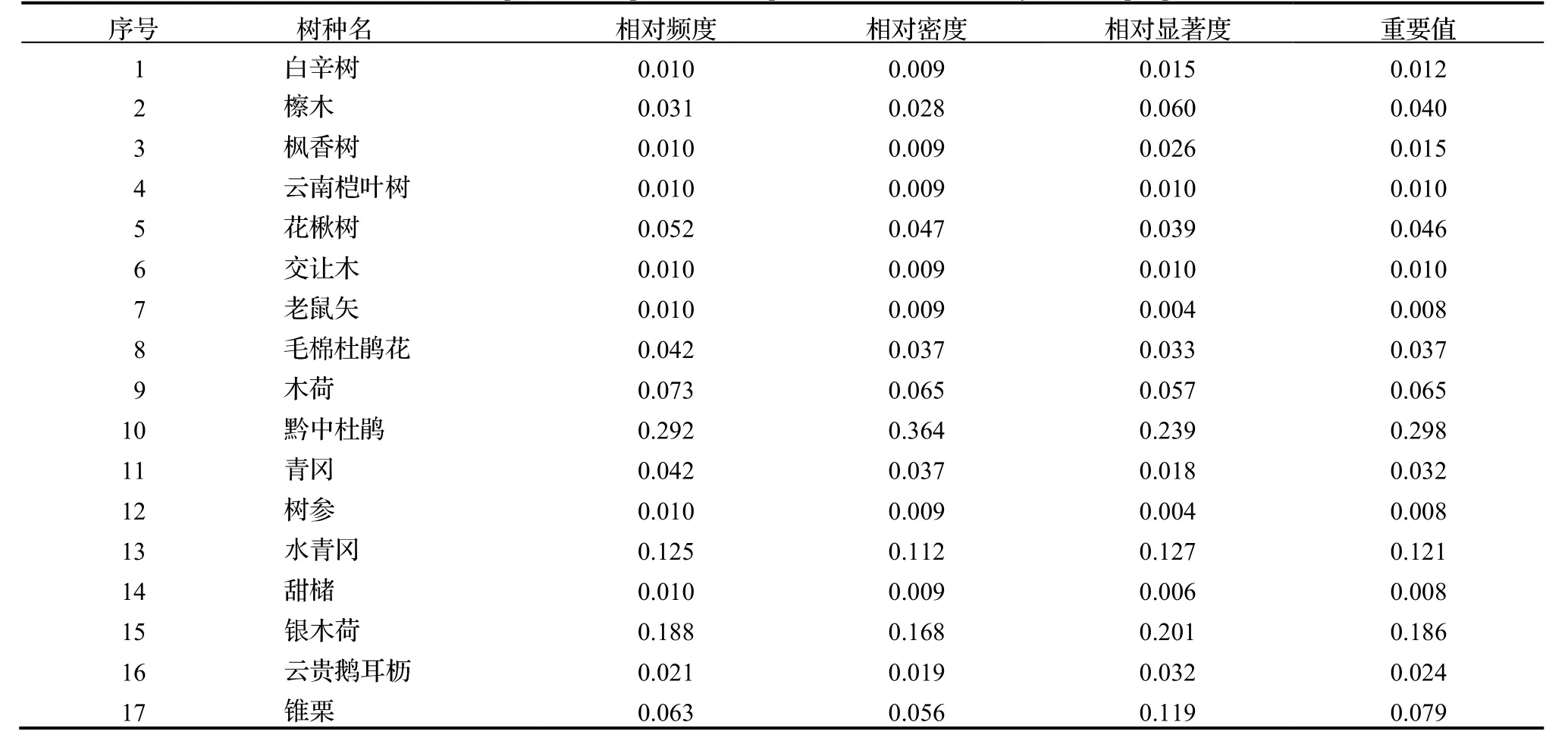
表2 黔中杜鵑群落喬木層種類組成及重要值Table 2 Composition of species and importance values at tree layer in sample plots
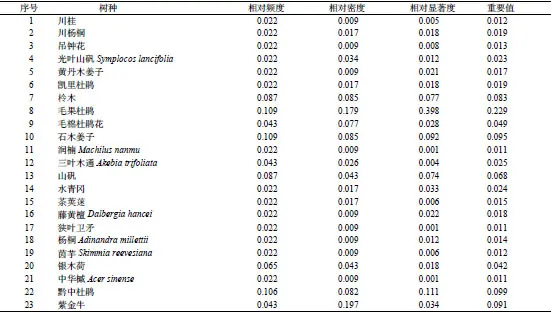
表3 黔中杜鵑群落灌木層種類組成及重要值Table 3 Composition of species and importance values at shrub layer in sample plots
表4 黔中杜鵑群落草本層種類組成及重要值Table 4 Composition of species and importance values at herb layer in sample plots
2.2 黔中杜鵑種群分布現狀及結構特征
黔中杜鵑種群結構呈現出不規則金字塔形(圖1),種群個體數量主要集中分布在Ⅲ~Ⅴ徑級,這三個徑級的個體數量占種群總數量的69.23%,說明該黔中杜鵑種群的中幼齡個體數很充足,但幼齡個體數較少,僅為種群總數量的11.54%,嚴重缺乏幼齡個體,中老齡個體數量總體呈現出下降趨勢。
采用種群動態量化方法對黔中杜鵑種群相鄰大小級的結構動態變化進行分析,以便對其結構特征進行更準確的評價。根據黔中杜鵑種群結構(圖1)計算得出,種群從Ⅰ級至Ⅷ級各徑級間的Vn分別為-80.00%,-37.50%,-20%,-41.18%,58.82%,71.43%,50%;Vpi為14.92%,考慮種群外部干擾,Vpi為0.67%。由此可見,雖然黔中杜鵑種群結構存在較大波動變化過程,但目前仍處于增長型。
2.3 黔中杜鵑靜態生命特征
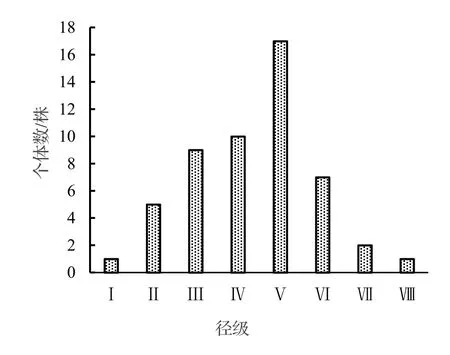
圖1 黔中杜鵑種群徑級結構Figure 1 Diameter class structure of Rh.feddei population
靜態生命表不僅可以反映種群從出生到死亡的數量動態,還可用于預測種群未來發展的趨勢[21]。由表5 可知,種群數量隨著徑級結構的增加呈現出先增大后減小的趨勢,而種群個體存活數lx和標準化存活數lnlx隨著徑級的增加逐漸減小,從x到x+1 徑級間隔期內標準化死亡數dx呈現出先下降后上升的趨勢,其dx在Ⅵ徑級時出現最大值,為170;種群死亡率qx從Ⅰ徑級至Ⅷ徑級隨演替的進行呈現出先減小后增大的趨勢,qx在Ⅵ和Ⅶ兩個徑級中出現“突變”,其qx明顯大于其它徑級,其次,qx較高的為Ⅰ徑級,說明黔中杜鵑種群在演替過程中幼年個體和老年個體最容易死亡而被淘汰;從x到x+1 徑級存活個體數Lx呈現出隨齡級的增加而減小的趨勢,個體期望壽命ex隨著年齡的增加逐漸降低,這與其生物學特性相一致;損失度Tx整體表現為先急劇下降再急劇上升再下降最后劇烈上升的趨勢,其損失度Tx在Ⅵ和Ⅶ徑級相對較高,其次為Ⅰ徑級和Ⅳ徑級。
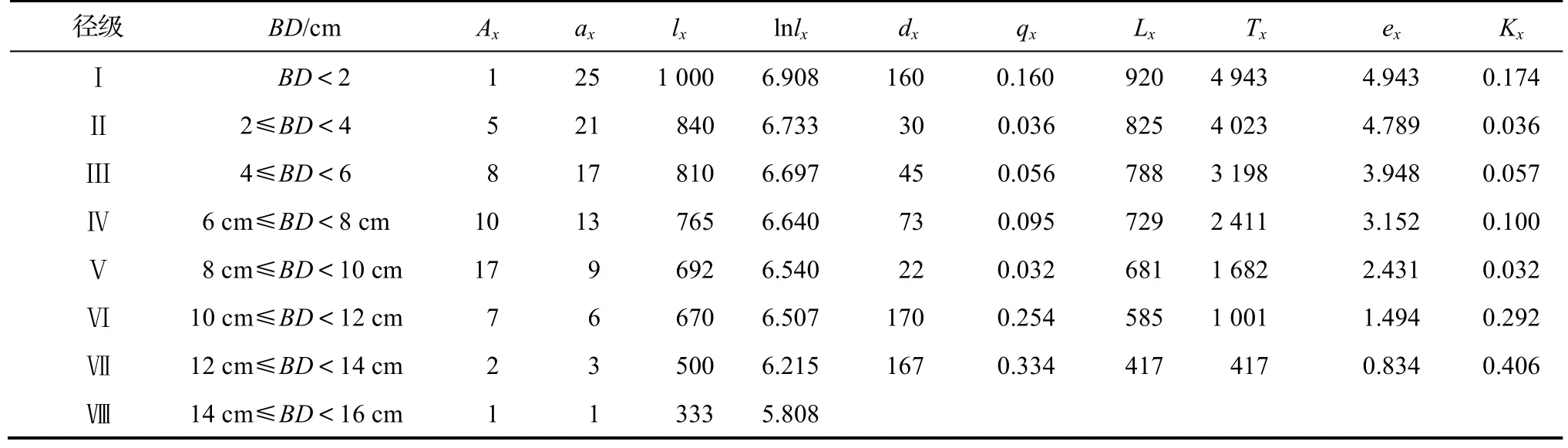
表5 黔中杜鵑種群靜態生命表Table 5 Static life table of Rh.feddei population
2.4 黔中杜鵑種群存活曲線特征
存活曲線可以有效地反映種群個體在各年齡級的存活狀況[22]。本研究以徑級(相對齡級)為橫坐標,以lnlx為縱坐標做出了黔中杜鵑種群存活曲線(圖2),依據Hett 和Loucks 的數學模型得出的檢驗方程及參數見表6。由模型檢驗結果顯示,兩種模型的擬合結果均達到顯著水平,但指數模型的R2值大于冪函數模型,且P值更小,說明黔中杜鵑的存活曲線趨近于Deevey-Ⅱ型。

表6 黔中杜鵑種群存活曲線檢驗模型Table 6 Test models for survival curve of Rh.feddei population
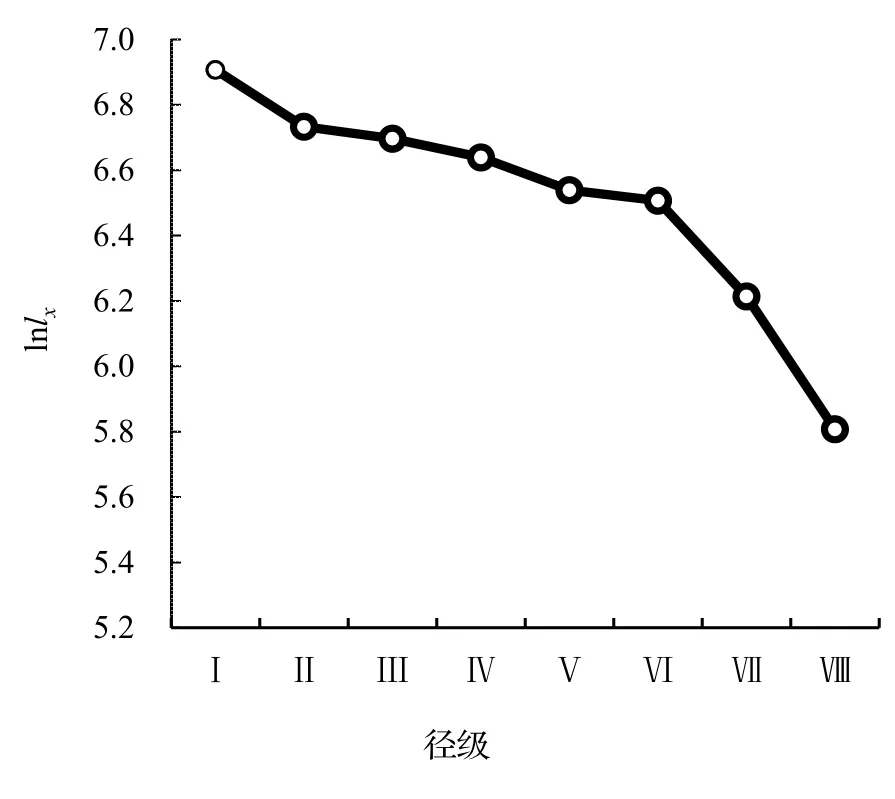
圖2 黔中杜鵑種群存活曲線Figure 2 Survival curve of Rh.feddei population
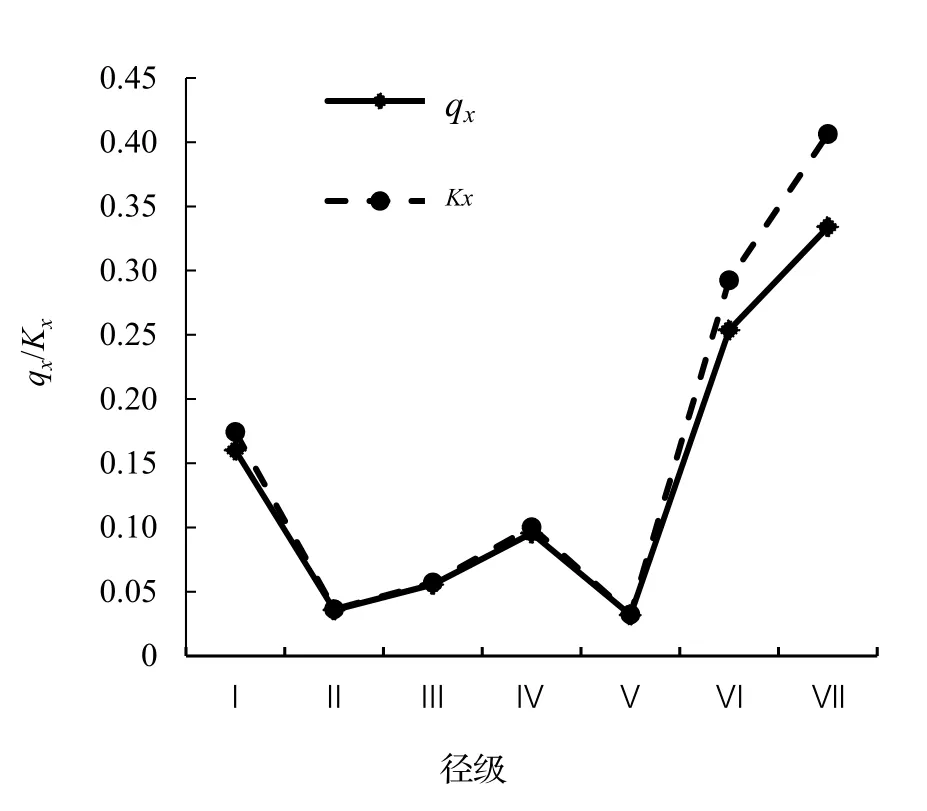
圖3 黔中杜鵑種群死亡率和虧損率曲線Figure 3 Mortality and vanish rate of Rh.feddei population
2.5 死亡率和虧損率曲線
以徑級為橫坐標,以各齡級的死亡率和虧損率為縱坐標做出的死亡率和虧損率曲線如圖3 所示。黔中杜鵑死亡率qx和虧損率Kx曲線變化趨勢一致,均呈現出先降低后升高再降低最后急劇增長的趨勢(圖3),認為黔中杜鵑種群數量具有前期短暫增加后逐漸減少,中期存在短暫的增長,后期急劇減少的特點。
2.6 黔中杜鵑種群生存分析
以徑級為橫坐標,函數值為縱坐標,繪制了黔中杜鵑野生種群生存率Si、累計死亡率Fi(圖4),死亡密度fi和危險率λi函數(圖5)。從圖4 可以看出,黔中杜鵑種群生存率和累計死亡率呈互補狀態,即隨著徑級的增加,黔中杜鵑種群的生存率下降,累計死亡率逐漸上升;種群在Ⅳ徑級達到平衡,即種群生存率和累計死亡率持平。黔中杜鵑死亡密度曲線總體呈現出先緩慢上升后緩慢下降的趨勢,整體趨勢較為平緩,而危險率曲線總體表現為先緩慢上升后有一個較小的下降趨勢,之后急劇上升再急劇下降,波動性比較大,表明黔中杜鵑種群數量受環境影響較大。
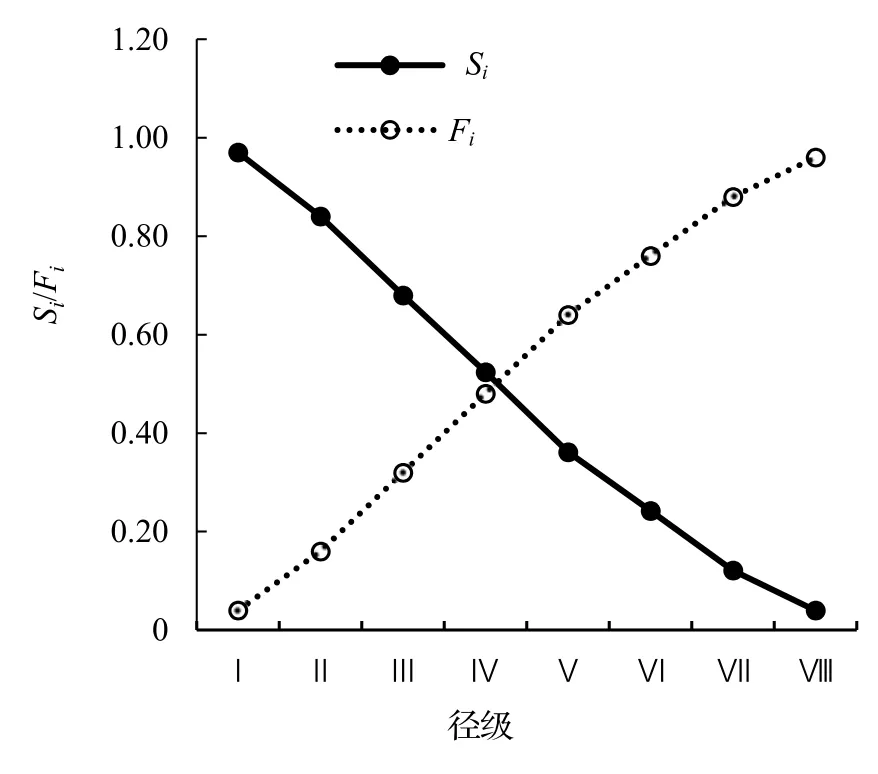
圖4 黔中杜鵑種群生存率和累計死亡率曲線Figure 4 Survival rate and cumulative mortality rate of Rh.feddei population
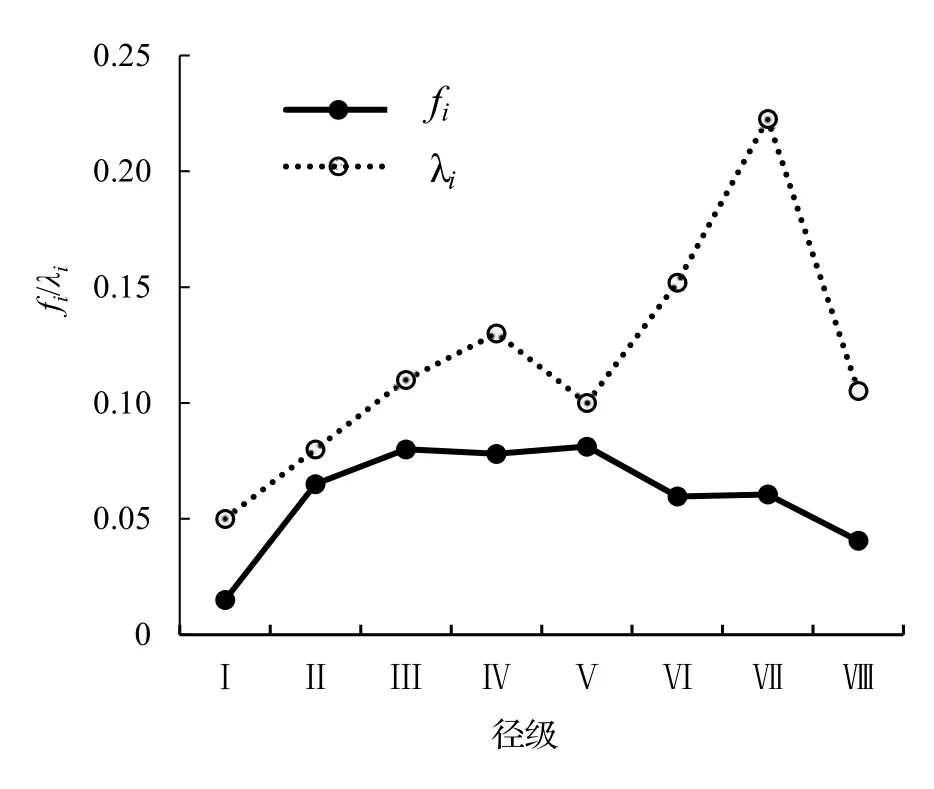
圖5 黔中杜鵑種群死亡密度和危險率曲線Figure 5 Mortality density and hazard rate of Rh.feddei population
2.7 黔中杜鵑種群數量的時間序列分析
以黔中杜鵑種群各齡級株數為原始數據,按照一次移動平均法則預測出各齡級在未來2,4,6 和8 個齡級時間后的個體數量(表7)。結果顯示,經歷2,4 個齡級時間,第2至第5 齡級的黔中杜鵑種群個體數均有不同程度的減少,而種群從第6 至第8 齡級開始之后均有不同程度的增加,經過6,8 個齡級時間,種群數量逐漸趨于穩定,說明一旦過了一定的生理年齡,種群就開始逐漸走向衰退,第2 至第3 齡級經過2 個齡級時間后個體數分別下降了40.0%和25.0%,第4至第5 齡級經過4 個齡級時間后個體數分別下降了40.0%和41.2%,這表明黔中杜鵑種群嚴重缺乏幼齡個體,綜合其整個發展趨勢來看,黔中杜鵑種群正常更新難以維持,未來會不同程度趨于衰退,甚至瀕危。
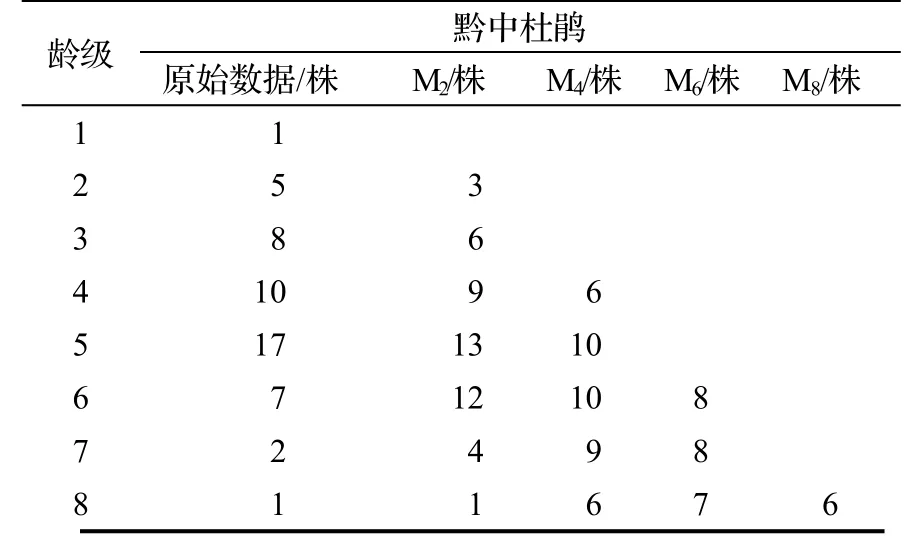
表7 黔中杜鵑種群動態變化的時間序列分析Table 7 Time sequence analysis of age structure of Rh.feddei population
3 結論與討論
植物種群結構特征不僅能夠體現種群內部個體的發展過程,而且也是植物對環境適應性的反映[23],其徑級結構能很好地反應種群動態變化[24]。本研究結果初步表明,黔中杜鵑種群結構整體呈現出不規則金字塔形;種群的中幼齡個體數充足,其個體數量占種群總數量的69.23%;嚴重缺乏幼齡個體數,僅為種群總數量的11.54%;中老齡個體數量總體呈現出下降趨勢;種群目前呈增長型狀態。說明黔中杜鵑種群結構總體呈相對穩定型,有向衰退轉化的趨勢,該種群雖然能夠暫時適應當地的生存環境,但由于幼齡個體數的缺乏,隨著時間的推移,種群整體的長期穩定性將難以維持。王立龍和易雪梅在裸果木Gymnocarpos przewalskii和水曲柳Fraxinus mandshurica的研究中也有相似的結論[25-26]。
黔中杜鵑種群靜態生命表分析表明:黔中杜鵑種群數量隨著齡級結構的增加呈現出先增大后減小的趨勢,而種群個體存活數(lx)隨著年齡的增加逐漸減小,個體期望壽命(ex)隨著年齡的增加逐漸降低。這與其生物學特性相一致。黔中杜鵑死亡率(qx)和虧損率(Kx)曲線變化趨勢一致,均呈現出先降低后升高再降低最后急劇增長的趨勢,表明黔中杜鵑種群數量具有前期短暫增加后逐漸減少,中期存在短暫的增長,后期急劇減少的特點。這與紅杉Larix potaninii種群的研究結論相似[27]。
黔中杜鵑種群生存率和累計死亡率呈互補狀態,即隨著齡級的增加,種群生存率下降,累計死亡率逐漸上升。黔中杜鵑種群在Ⅳ徑級達到平衡,即種群生存率和累計死亡率持平;死亡密度曲線總體呈現出先緩慢上升后緩慢下降的趨勢,整體趨勢較為平緩,而危險率曲線總體表現為先緩慢上升后有一個較小的下降趨勢,之后急劇上升再急劇下降,波動性比較大,表明黔中杜鵑種群數量受環境影響較大。姜在民在瀕危植物羽葉丁香Syringa pinnatifolia種群的研究中也有相似的結論[28]。
采用時間序列分析中的“一次移動平均法”對黔中杜鵑種群的年齡結構進行預測,結果表明:種群一旦過了一定的生理年齡,就會開始逐漸走向衰退。從黔中杜鵑整個發展趨勢來看,該物種種群正常更新難以維持,未來會不同程度趨于衰退,甚至瀕危。因此,建議加強黔中杜鵑自生生物學特性研究,進一步摸清其生物學特性,開展種群的更新定位觀測研究,包括不同群落枯落物對黔中杜鵑種子的自然萌發和苗木生長的影響,還有黔中杜鵑幼苗建成機制;同時,開展有性與無性繁殖方式相結合,進行回歸引種試驗,擴大種群數量,提升種群自身抵御種群衰退的風險。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
哲學評論(2021年2期)2021-08-22 01:53:34
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
