三裂葉薯NBS-LRR類抗病基因的篩選鑒定與結構分析
2020-10-09 14:32:00胡韻卓石媛媛黃小芳a畢楚韻a周麗香梁才曉黃碧芳許明林世強a陳選陽a
熱帶亞熱帶植物學報 2020年5期
胡韻卓, 石媛媛, 黃小芳a,, 畢楚韻a,, 周麗香, 梁才曉, 黃碧芳,許明, 林世強a,*, 陳選陽a,*
(福建農林大學, a. 作物生物技術福建省高校重點實驗室;b. 植物保護學院;c. 農學院;d. 生命科學學院,福州 350002)
Flor提出“基因對基因”假說,認為對于病原的每個無毒基因(Avr),宿主都有1個相應的抗性基因(R基因),這2個基因產物互作激活宿主的防御反應, 如超敏反應(hypersensitive response, HR)[1]。根據結構域基序,R基因的編碼產物R蛋白分為5類[2],第一類含有絲氨酸-蘇氨酸激酶結構域和十四烷基化結構域;第二類包含LRR (leucine-rich repeat)結構域、NBS (nucleotide-binding site)結構域和位于N端的亮氨酸拉鏈(leucine zipper)或卷曲螺旋(coiled coil)序列;第三類和第二類相似,但N端序列為TIR(toll and Interleukin 1 receptor)結構域;第四類具有跨膜結構域和胞外LRR結構域,但是缺失NBS; 第五類含有跨膜結構域、胞外LRR結構域和絲氨酸-蘇氨酸激酶結構域。在這些結構域中,NBS結構域是這類蛋白家族的關鍵結構域,可能通過與核苷酸結合或水解來影響R蛋白的功能。
三裂葉薯(Ipomoea triloba)是栽培甘薯(I.batatas)的二倍體近緣野生種,主要分布于我國臺灣、廣東和美洲熱帶等地,具有抗旱、抗病等優良性狀[3-4]。研究表明[5-6],野生種三裂葉薯可作為克服種間雜交不親和性,通過胚胎培養獲得雜種的重要材料。三裂葉薯基因組測序于2018年完成[7],本研究基于測序結果對其進行CDS (coding domain sequence)區域預測,檢索其中的NBS-ARC家族基因并進行結構分析、染色體定位、保守結構域分析和進化樹構建,為三裂葉薯的抗病研究及甘薯的抗性育種提供參考。
1 材料和方法
1.1 材料
三裂葉薯(Ipomoea triloba)全基因組序列從https://www.ncbi.nlm.nih.gov/genome/?term=Ipomoea triloba L.下載,共16條染色體的DNA序列。
1.2 基因注釋
通過snap程序應用擬南芥(At.hmm)、線蟲(Ce.hmm)和水稻(Os.hmm)的HMM模型對三裂葉薯全基因組序列CDS區域進行檢索,所得作為訓練三裂葉薯特異HMM模型的文件(Itl.hmm), 后以Itl.hmm作為檢索工具預測屬于三裂葉薯基因組的CDS區域,并得到其全基因組蛋白質序列。
1.3 基因預測
通過pfam下載含有NB-ARC家族序列信息的HMM文件(PF00931),作為檢索序列對三裂葉薯的全基因組蛋白質序列進行NB-ARC結構域搜索。從中選取可用序列(E-value≤1E-60),用Sam- tools構建三裂葉薯的NB-ARC蛋白本地數據文件,同時,用Clustal Omega進行比對并用hmmbuild建立三裂葉薯特異HMM模型,并再次檢索全基因組蛋白序列預測NB-ARC結構域,得到三裂葉薯高特異的NB-ARC序列信息。最后導入NCBI Conserved Domains Tool和Interproscan進行篩選,剔除結構域缺失嚴重序列,得到三裂葉薯NB-ARC基因家族蛋白質序列數據庫。
1.4 保守結構域分類
從Pfam中下載TIR HMMs、RPW8 HMMs和LRR HMMs模型,合并后用hmmpress程序建立TRL.hmm模型,隨后通過hmmscan對三裂葉薯的NBS-LRR家族基因信息庫進行結構域預測,并結合Interproscan查看結果對TIR/RPW8/LRR結構域進行類型分類和統計。運用Pepcoil與Paircoil2對三裂葉薯NBS-LRR家族基因編碼的蛋白質序列中的CC結構域進行預測和檢測。
1.5 染色體定位
對三裂葉薯16條染色體上的NBS-LRR家族基因位置信息進行統計,根據其在染色體上的位置分成上、中、下3個部分,每個部分按照N、NL、CN、CNL、TN、TNL和RN共7個類型進行分類統計。
1.6 基序分析
將三裂葉薯NBS-LRR家族基因進行分類整理后,從中篩選出CC-NBS-LRR、TIR-NBS-LRR和RPW8-NBS共3種NBS亞家族類型的氨基酸序列。保守基序的分析與識別通過MEME進行,并將運行后得到的.xml文件利用TBtools進行處理。
1.7 進化樹構建
將篩選得到的3種NBS-LRR亞家族類型編碼的CC-NBS-LRR、TIR-NBS-LRR和RPW8-NBS氨基酸序列利用Clustal Omega程序進行多序列比對,隨后使用Gblocks提取保守序列,進一步用Jalview手動矯正。以比對結果為基礎,應用MEGA X軟件,Maximum Likehood法模式為WAG with Freqs. (+F)model,校驗參數Bootstrap=500運行生成三裂葉薯NBS-LRR基因家族系統進化樹。
2 結果和分析
2.1 NB-ARC基因鑒別及其分類
采用snap程序對三裂葉薯全基因組序列進行CDS區域識別預測,經篩選后獲得282個NB-ARC家族基因,占全基因組基因數(98025)的0.288%。根據TIR/RPW8/LRR結構域和CC結構域,這些基因可進一步分為N (NBS)、NL (NBS-LRR)、CN (CCNBS)、CNL (CC-NBS-LRR)、TN (TNL-LRR)、TNL(TNL-NBS-LRR)和RN (RPW8-NBS)共7個亞家族類型(表1),分別有80、83、28、57、10、23和1個基因。不同基因的NBS/LRR結構域重復數不同, 在染色體中的分布位置也存在差異。此外,還含有馬鈴薯抗X病毒和類似蛋白的RX-CC_like結構域(coiled coil domain of the potato virus X resistance protein and similar proteins),即Cx結構域,與Pepcoil程序得到的CC結構域不同。根據基因結構域的特點和差異,將NBS/LRR基因類型進一步分為40個小類(表1)。
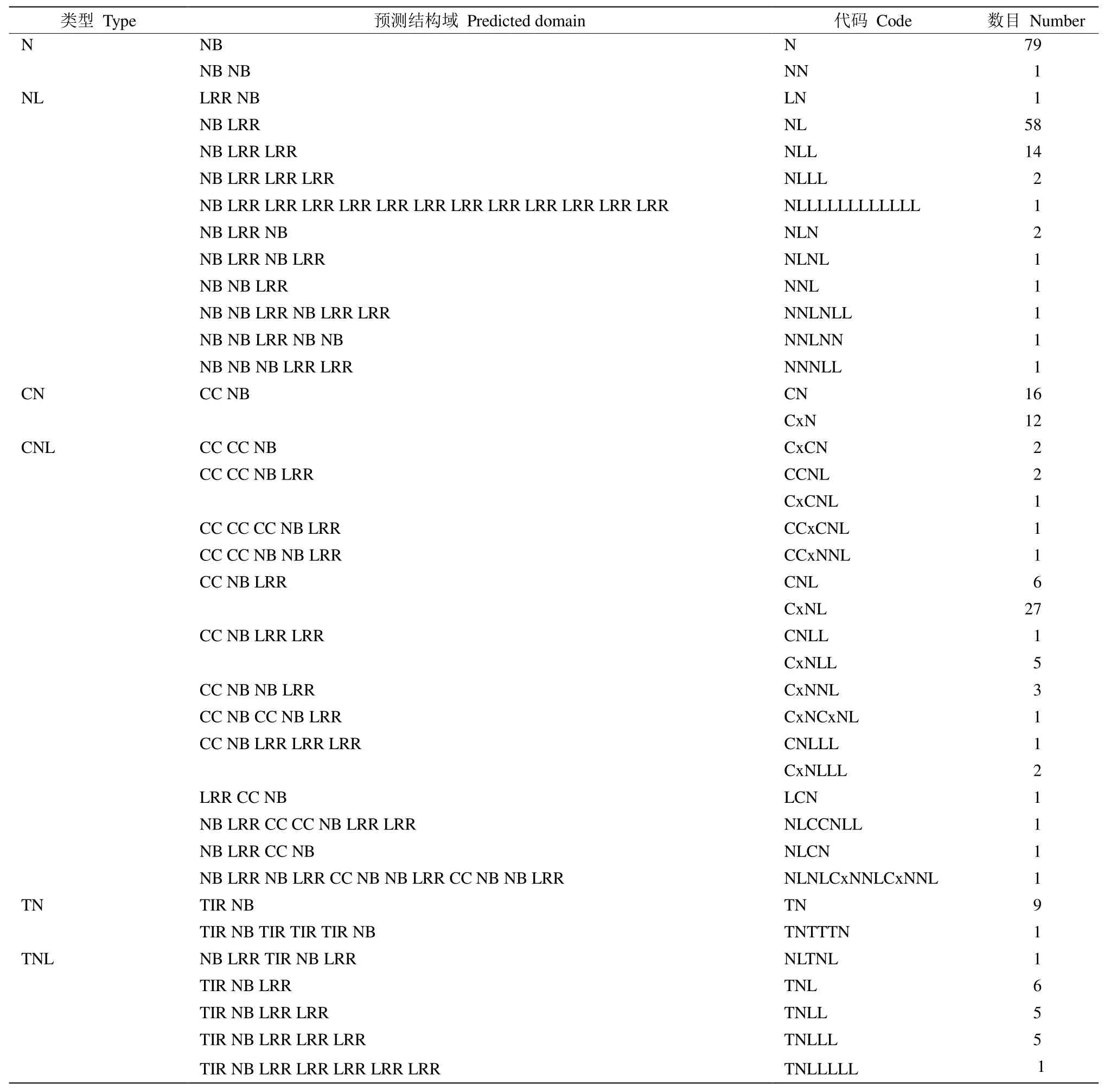
表1 三裂葉薯NBS-LRR基因的數量和分類Table 1 Number and type of NBS-LRR genes of Ipomoea triloba

續表(Continued)
2.2 NBS-LRR家族基因的染色體分布
NBS-LRR家族基因在三裂葉薯16條染色體上的分布差異明顯(表2),不同的亞家族類型,CN型、CNL型、N型、NL型、TN型、TNL型和RN型在各染色體上的數量和位置較不均勻。在第16號染色體僅有1個NBS-LRR家族基因,而在第15號染色體多達65個。NBS-LRR家族基因在染色體的位置也有差異,第4、13和14號染色體的NBS-LRR家族基因主要位于染色體的中部和上部,而在第7、11和15號染色體則以中部和下部居多(表2)。
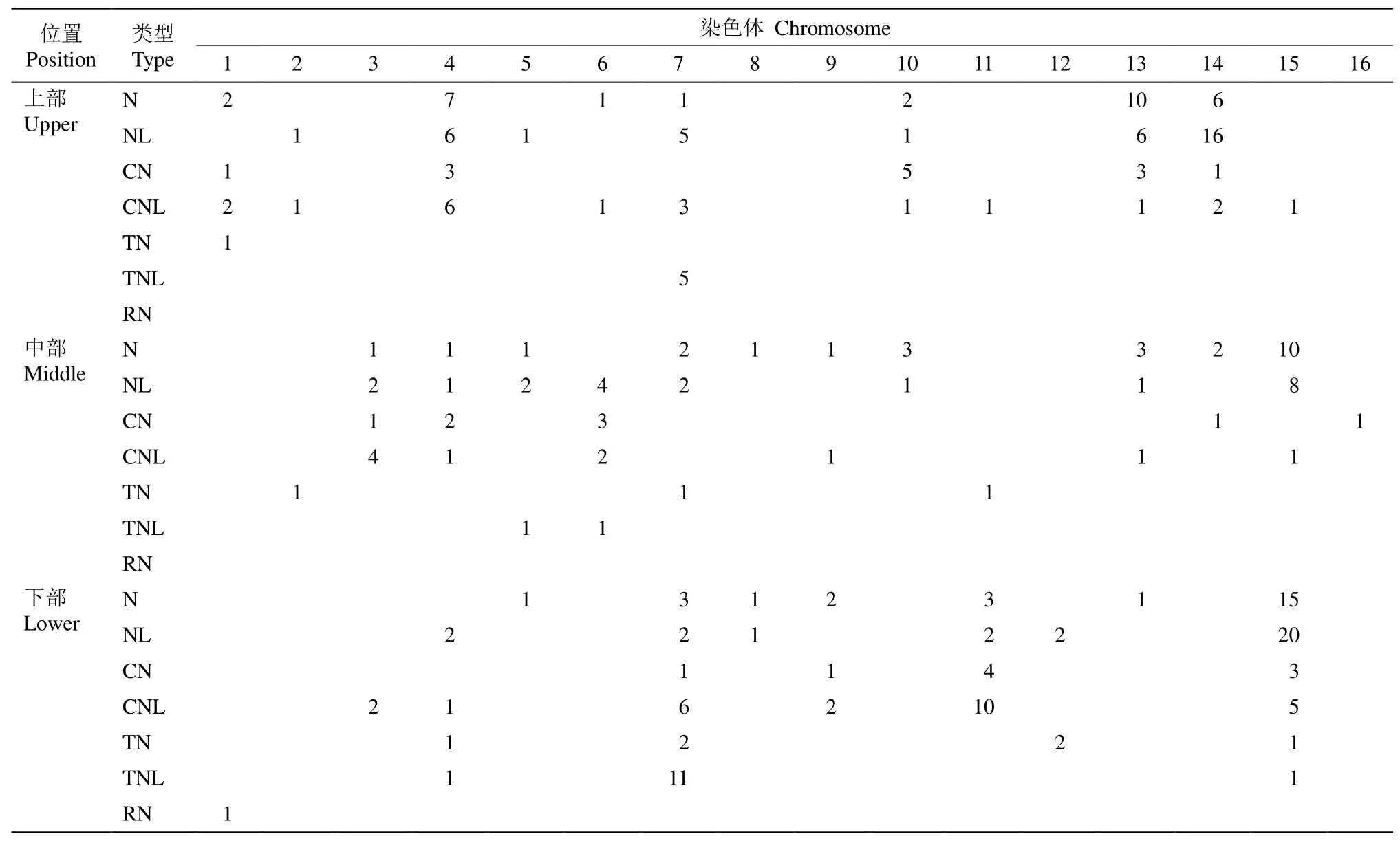
表2 三裂葉薯NBS-LRR家族基因在染色體上的分布Table 2 Chromosomal location of NBS-LRR family genes of Ipomoea triloba
NBS-LRR家族基因在各染色體上分布形式也存在明顯不同,有些是以單基因分布于染色體,而有些則以基因簇的形式存在。三裂葉薯每條染色體上的基因簇數量也存在較大差異,如第15號染色體的基因簇最多,有10個;第16號染色體則沒有基因簇(表3)。所有染色體總共有55個基因簇,包含了179個NBS-LRR基因,占NBS-LRR家族基因總基因數(282)的63.5%。
2.3 基序分析
分別將三裂葉薯NBS-LRR蛋白家族中CNL和TNL亞家族氨基酸序列信息利用MEME程序檢測,得到15個相似度較高的結構域。CNL亞家族氨基酸序列(共57條)結構域的分布(圖1)有以下規律:motif 9-motif 10-motif 1 (P-loop)-motif 4 (RNBS-A)-motif 2 (kinase 2)-motif 15 (RNBS-C)-motif 6(GLPL)-motif 8-motif 3 (RNBS-D)-motif 12-motif7-motif 11-motif 14-motif 11-motif 5-motif 13。TNL亞家族氨基酸序列(共22條)結構域的分布有以下規律(圖2):motif 11 (TIR-1)-motif 3 (TIR-2)-motif 1(TIR-3)-motif 13 (TIR-4)-motif 5 (P-loop)-motif 10-motif 2 (kinase 2)-motif 9 (RNBS-B)-motif 4 (RNBSC)-motif 14 (GLPL)-motif 6-motif 8 (RNBS-D)-motif 7 (MHDV)-motif 12-motif 15-motif 12-motif 15。
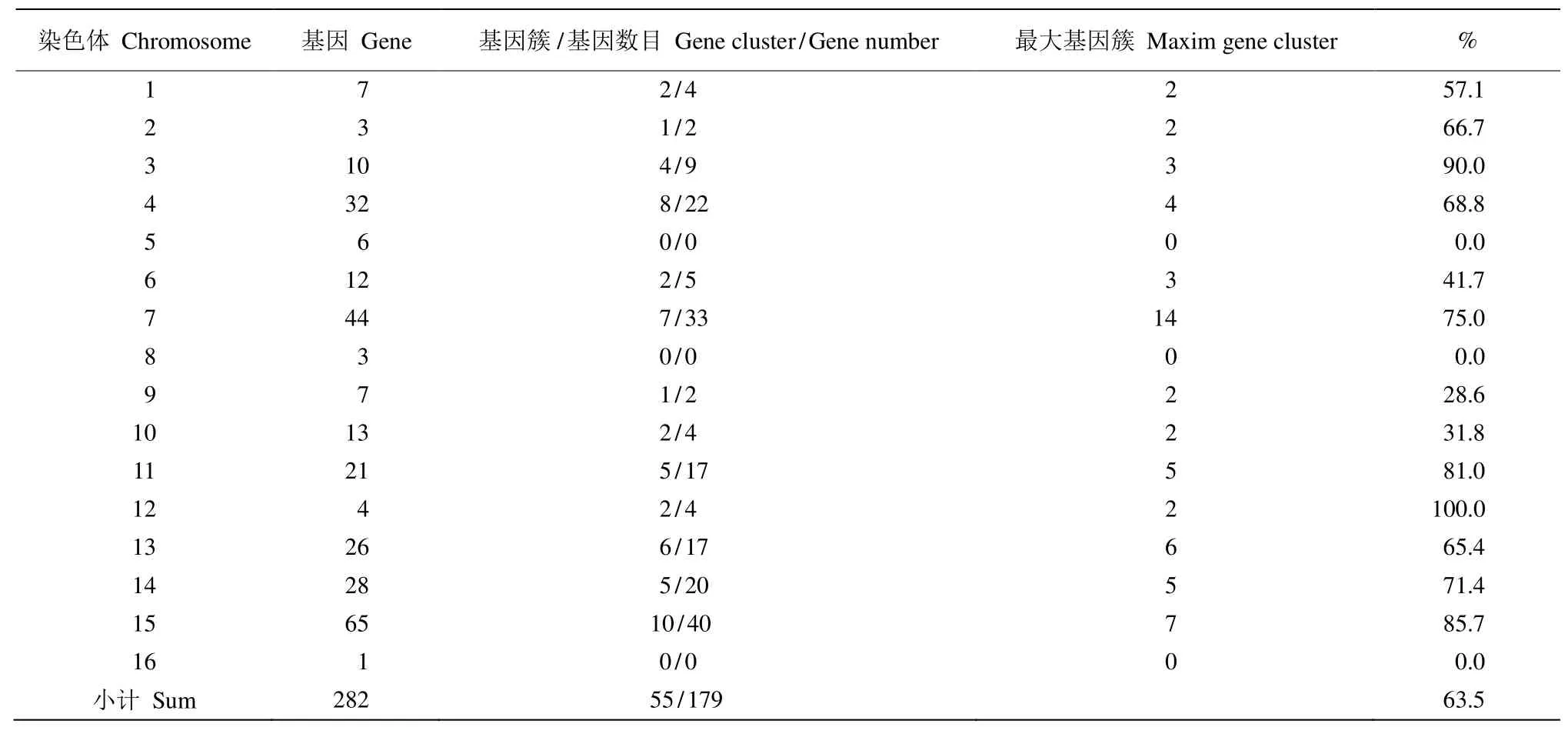
表3 三裂葉薯NBS-LRR基因家族基因簇統計表Table 3 Statistics of NBS-LRR family gene clusters in Ipomoea triloba
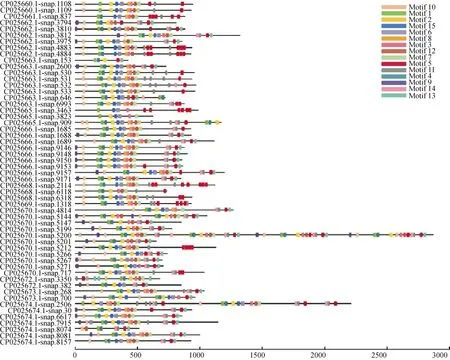
圖1 三裂葉薯NBS-LRR基因家族中CNL亞家族蛋白保守結構域分布Fig. 1 Distribution of conservative domains of CNL subfamily proteins in NBS-LRR gene family of Ipomoea triloba
2.4 結構域保守性分析
在植物的NBS-LRR蛋白家族中NB-ARC區域相對保守,通常有8個保守基序[8]: P-loop、RNBS-A、Kinase2、RNBS-B、RNBS-C、GLPL、RNBS-D和MHDV。對三裂葉薯NBS-LRR家族氨基酸序列利用Clustal Omega篩選出20條序列進行多重比對,結果表明,具有5個保守性較高的結構域[9-11](圖3),分別為P-loop (Kinase 1)、Kinase 2、RNBS-B、GLPL和MHDV。其中P-loop、Kinase 2、RNBS-B、GLPL、MHDV最保守的氨基酸序列分別為SIV GMGGI GKTT+AR++F (下劃線為氨基酸保守性較強序列)、R+LLVLDDV、GS+I+LTTR++++L、AGGLPLAL+V+G、GKMHDLV++MG。
根據MEME結構域搜索結果,對CNL和TNL亞家族基因結構域進行基因識別和定位[12]。從表4可見,TNL和CNL亞家族中分別有11和7個保守結構域,其中TNL亞家族NB-ARC結構域中保守基序分別為P-loop、Kinase 2、RNBS-B、RNBS-C、GLPL、RNBS-D和MHDV,缺失RNBS-A保守基序。CNL亞家族NB-ARC結構域保守基序分別為P-loop、RNBS-A、Kinase 2、RNBS-C、GLPL、RNBSD和MHDV,缺失RNBS-B保守基序。TNL和CNL亞家族中共有的保守基序分別為P-loop (Kinase 1)、Kinase 2、RNBS-C、GLPL、RNBS-D和MHDV。此外,在TNL亞家族中TIR結構域檢測到4個保守基序(TIR 1~TIR 4)[13]。
2.5 NBS-LRR亞家族系統進化樹分析
將三裂葉薯的NBS-LRR家族蛋白中CC/TIR/RPW8/NBS/LRR結構域序列分別提取,使用MEGA X構建進化樹(圖4),可見有3個明顯的分支,分別為CNL型(藍色)、TNL型(紫色)和RN型(黃色)。進化樹分支中有些基因分支路徑很長,如CP025666.1-snap.8798、CP025670.1-snap.5147等,推測這類基因的“祖先”基因在較早時即已發生分化,相應的基因序列也發生了較大的變化。有些基因序列的分支距離較近, 如CP025666.1-snap.1140和CP025666.1-snap.1141、CP025663.1-snap.531和CP025663.1-snap.532,說明這些變化發生的時間較晚。
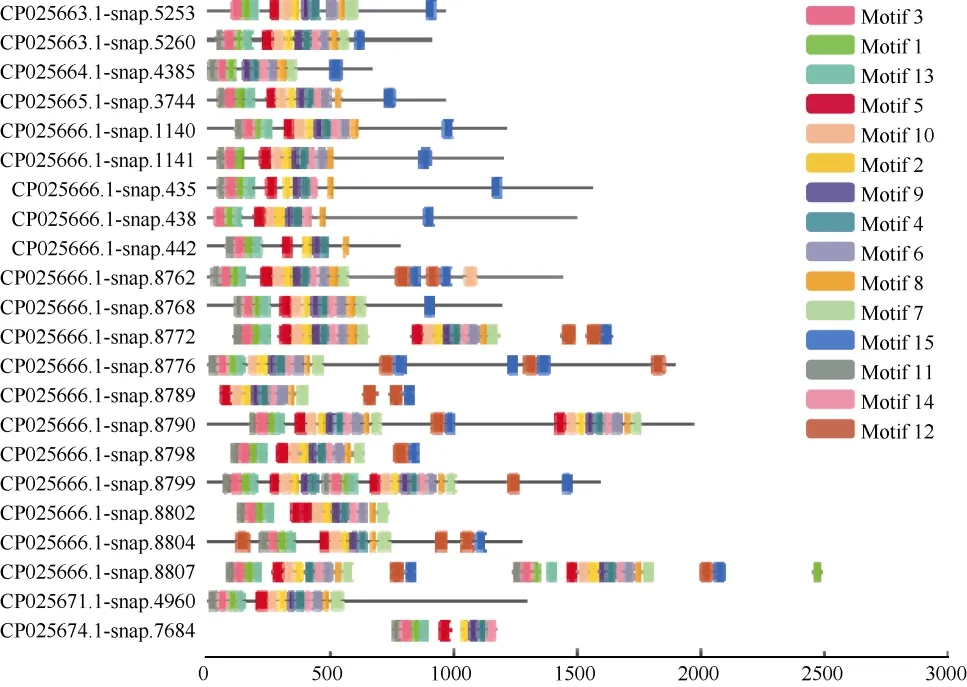
圖2 三裂葉薯TNL亞家族蛋白保守結構域分布Fig. 2 Distribution of conservative domain of TNL subfamily proteins in Ipomoea triloba
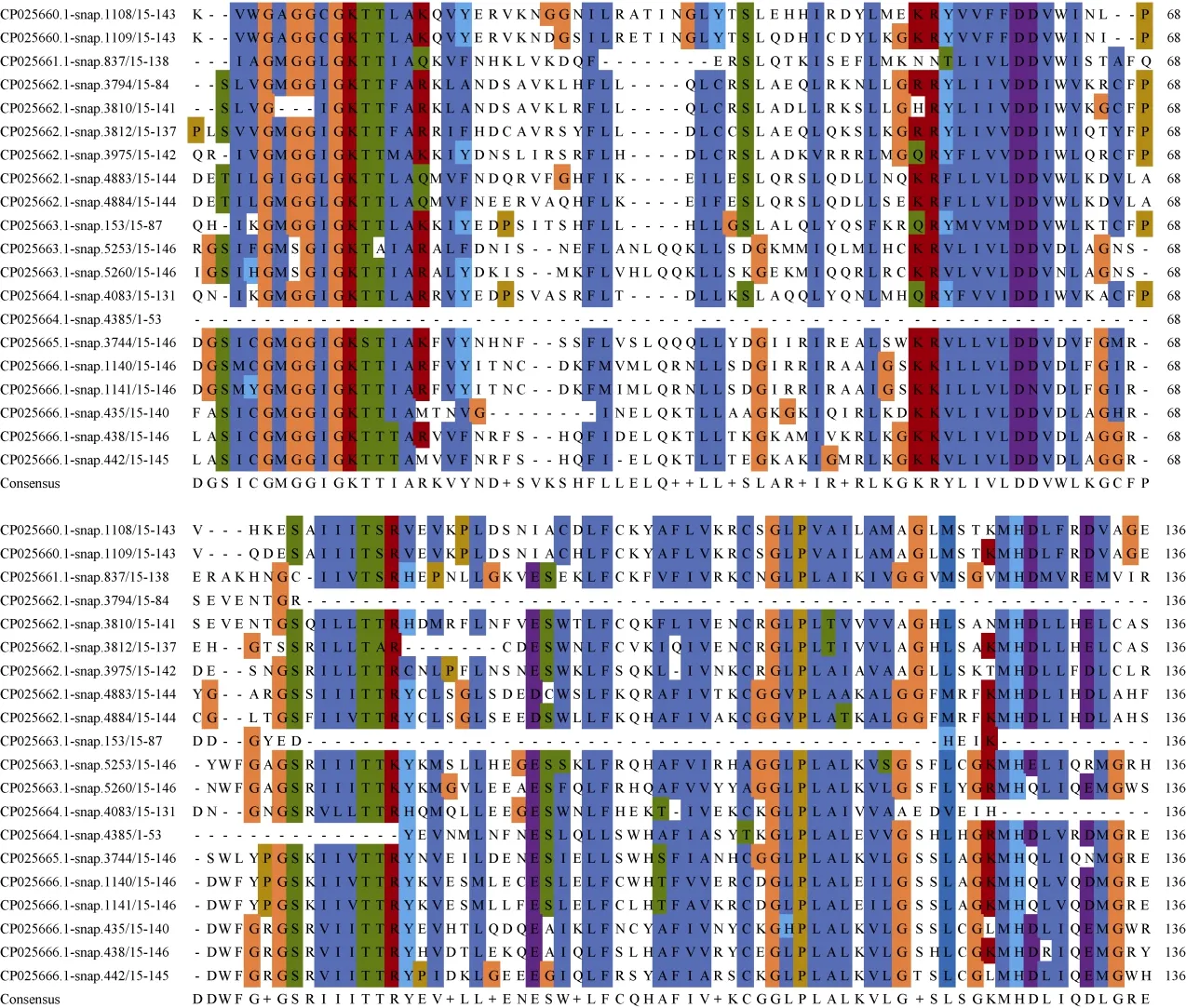
圖3 三裂葉薯CNL和TNL亞家族NB-ARC保守性分析Fig. 3 Conservation analysis of the NB-ARC in CNL and TNL subfamilies in Ipomoea triloba

表4 三裂葉薯NBS-LRR家族蛋白保守結構域和氨基酸保守性分析Table 4 Conservative domains and amino acids of NBS-LRR family proteins in Ipomoea triloba
3 結論和討論
植物R基因在長期的抗病過程中監控識別病原微生物產生的無毒基因(Avr),從而引發植物細胞過敏性壞死,產生免疫反應。R基因與Avr基因間互作模式包括直接作用、間接作用和轉錄調控[14]。通過克隆植物體內相關的抗病R基因,研究其與病原物的互作模式與機制,或將其遺傳轉化到目標作物中, 增強作物的抗病能力,有利于提高作物產量,保障生產安全。
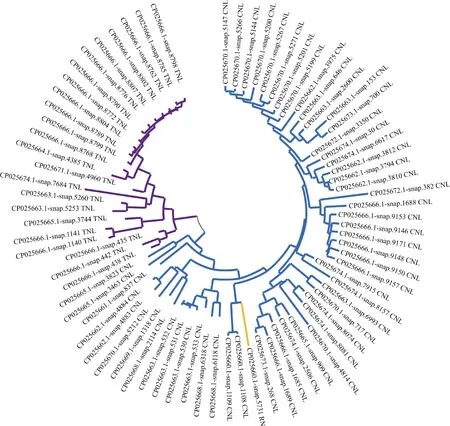
圖4 三裂葉薯NBS-LRR家族基因系統進化樹Fig. 4 Phylogenetic tree of NBS-LRR family genes in Ipomoea triloba
甘薯在生產上面臨著許多病害的威脅,如南方薯區的蔓割病、薯瘟病,北方薯區的莖線蟲病、黑斑病、根腐病等,以及最近時有發生的莖腐病等病害,都嚴重影響甘薯生產的安全。甘薯抗性是產量和品質的堅實保證[15],因而抗性育種是甘薯育種的重要方向。甘薯種質資源是甘薯育種工作的基礎[16],然而,甘薯種質資源在新品種育種工作中存在利用率低、育成品種遺傳背景狹窄、優秀資源篩選困難等問題[17-18]。因此,如何充分發掘、利用野生種質資源,拓寬甘薯遺傳背景,對于選育具有突破性的甘薯新品種具有重要意義。研究表明,甘薯與其近緣野生種三裂葉薯雜交不親和,無法直接利用其抗性基因,但應用原生質體細胞融合法可以獲得三裂葉薯和甘薯的種間體細胞雜種植株,或者可以通過克隆其抗性基因,通過遺傳轉化提高栽培種甘薯的抗性水平,以充分發揮三裂葉薯基因資源在甘薯育種中的巨大潛力[5,19-20]。
值得注意的是,甘薯栽培種為同源異源六倍體,三淺裂野牽牛(I. trifida)存在二倍體、四倍體和六倍體,一套染色體均為15條,但數據庫中三裂葉薯基因組的染色體數為16,本文即以三裂葉薯16條染色體的全基因組序列為研究對象開展了相關的研究,研究結果有待于與甘薯和三淺裂野牽牛基因組進行進一步的比較,分析它們之間的異同之處,將有利于甘薯的進化研究和育種工作。
對作物全基因組NBS-LRR基因進行鑒定分析,有利于其抗性資源的應用[21],本研究對三裂葉薯全基因組蛋白序列進行了預測,并鑒定、篩選了NBS-LRR類抗病基因,獲得NBS-LRR家族基因的數量、在染色體上定位與分布,分析了基因結構域特點,構建了進化樹,為進一步發掘其抗性基因資源,研究甘薯抗病分子機制,提高甘薯抗性育種水平提供了參考。
