miR-181b-5p靶向PDCD4基因調控高糖誘導血管內皮細胞增殖和凋亡的機制
2020-10-12 03:07:36吳源白淑榮任紅梅尹彩君魏述軍
中國老年學雜志 2020年19期
關鍵詞:檢測
吳源 白淑榮 任紅梅 尹彩君 魏述軍
(寧夏回族自治區人民醫院 1心血管中心,寧夏 銀川 750021;2呼吸科;3心內科;4內分泌科)
糖尿病是一種常見的內分泌疾病,致病因素很多,長期糖尿病可引起動脈硬化、冠心病、高血壓、腎病、足潰瘍等多種慢性并發癥,嚴重危害患者生命〔1〕。血管內皮細胞(VEC)是位于血管最內層的一層細胞,作為功能異常活躍的內分泌器官可合成及分泌大量的血管活性物質,維持血管正常生理功能〔2〕。糖尿病血管病變是糖尿病人最常見的并發癥,是糖尿病致死、致殘的主要原因〔3〕。程序性細胞死亡因子(PDCD)4是一個細胞凋亡因子,可以抑制細胞凋亡的發生,位于染色體10q24〔4〕。研究表明,PDCD4蛋白在高濃度葡萄糖誘導的VEC中上調表達,參與高糖誘導的VEC凋亡過程,并影響細胞的增殖〔5〕。微小RNA(miRNA)是在生物體內發現的一類由19~25個堿基構成的內源性小分子RNA,通過連接于靶基因mRNA的3′UTR,降解靶基因mRNA或阻止其蛋白翻譯〔6〕。高糖環境可上調人臍靜脈內皮細胞(HUVEC)中miRNA-503表達,通過抑制胰島素樣生長因子1的表達,減少HUVEC的增殖,促進其凋亡〔7〕。在血栓素的刺激下,HUVEC中miR-181b的表達迅速下調,保護VEC損傷〔8〕。miR-181b-5p在VEC的研究尚未清楚,因此,本研究采用體外培養人血管臍靜脈內皮細胞的方法,探究miR-181b-5p對高糖誘導的HUVEC增殖和凋亡的影響及調控機制。
1 材料與方法
1.1材料 HUVEC購自美國ATCC公司;胎牛血清、DMEM培養液、胰蛋白酶購自Gibco公司;葡萄糖購自北京百歐博偉生物技術有限公司;LipofectamineTM2000轉染試劑購于美國Invitrogen公司;噻唑藍(MTT)、二甲基亞砜(DMSO)購于美國Sigma公司;Trizol試劑、聚偏氟乙烯(PVDF)膜、電化學發光液、雙熒光素酶報告基因檢測試劑盒購自北京Solarbio公司;放射免疫沉淀法(RIPA)裂解液、十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)試劑盒、二喹啉甲酸(BCA)試劑盒購自上海碧云天生物技術有限公司;Annexin V-異硫氰酸熒光素(FITC)/碘化丙啶(PI)凋亡檢測試劑盒購自美國Coulter公司;反轉錄試劑盒、實時熒光定量試劑盒購于TaKaRa公司;兔多克隆抗體PDCD4購自美國Bioworld公司;兔多克隆抗體GAPDH、辣根過氧化物酶(HRP)標記的山羊抗兔免疫球蛋白(Ig)G購自Abcam公司;control mimic、miR-181b-5p mimic、control inhibitor、miR-181b-5p inhibitor由上海吉瑪公司設計合成;pcDNA-PDCD4過表達載體由金瑞斯生物科技公司構建;引物合成由上海生工生物工程股份有限公司合成。
1.2細胞培養 HUVEC細胞使用含10%胎牛血清的DMEM培養基,于37℃ 5%CO2濕度飽和的條件下培養,每隔24 h換液1次,待細胞融和至60%~ 70%時,加入胰蛋白酶進行消化傳代。選取對數生長期的細胞進行實驗。
1.3細胞轉染與分組 收集對數生長期的HUVEC,加入胰酶消化并重懸,以1×105個/孔接種至24孔板,常規培養24 h。參照LipofectamineTM2000轉染試劑盒說明書進行轉染,miR-NC組轉染control mimic,miR-181b-5p組轉染miR-181b-5p mimic,anti-miR-NC組轉染control inhibitor,anti-miR-181b-5p組轉染miR-181b-5p inhibitor,si-PDCD4組轉染PDCD4 siRNA,miR-181b-5p+pcDNA組共轉染miR-181b-5p mimic和pcDNA,miR-181b-5p+pcDNA-PDCD4組共轉染miR-181b-5p mimic和pcDNA-PDCD4,轉染至HUVEC細胞。之后對miR-181b-5p組、si-PDCD4組、miR-181b-5p+pcDNA組和miR-181b-5p+pcDNA-PDCD4組給予含33 mmol/L葡萄糖的培養基培養,標記為高糖組。其余用濃度5.5 mmol/L葡萄糖培養的細胞標記為對照組。
1.4實時熒光定量-聚合酶鏈反應(qRT-PCR)檢測 取上述各組對數生長期的細胞,Trizol試劑提取各組HUVEC細胞總RNA,檢測RNA純度和濃度后使用TaKaRa反轉錄試劑盒將RNA反轉錄成cDNA,按照TaKaRa 熒光定量試劑盒使用說明配制反應體系,置于實時熒光定量儀上進行PCR擴增,每個樣品重復3次。采用F=2-△△Ct法進行結果分析。以GAPDH作為內參,引物如下:GAPDH正義鏈:5′-GCACCGTCAAGGCTGAGAAC-3′,GAPDH反義鏈:5′-TGGTGAAGACGCCAGTGGA-3′。
1.5Western印跡檢測 取上述各組對數生長期的細胞加入RIPA裂解液,置于冰上裂解細胞,4℃,26 750 r/min離心20 min,收集上清。將蛋白樣品進行SDS-PAGE電泳后轉至PVDF膜上,5%脫脂奶粉封閉液室溫封閉1 h。分別加入PDCD4抗體(1∶1 000)和GAPDH抗體(1∶1 000),4℃孵育過夜,TBST洗膜3次,每次10 min;加入HRP標記的二抗(1∶5 000)室溫孵育2 h,TBST洗滌;電化學發光顯影,每個蛋白樣品重復3次。
1.6MTT檢測細胞增殖 取上述各組對數生長期的細胞,將其制備成1×104個/ml的單細胞懸液后,每孔100 μl接種至96孔板中,培養于37℃、5%CO2飽和濕度條件下。在各組細胞培養至24 h、48 h、72 h時加入20 μl(5 g/ L)的MTT溶液,培養4 h,棄上清,每孔加入150 μl DMSO,震蕩使結晶溶解,在490 nm波長下檢測細胞吸光度值(OD)。細胞活性與吸光值呈正比。
1.7流式細胞儀檢測細胞凋亡 取上述各組細胞,加入預冷的PBS洗滌細胞,胰蛋白酶消化細胞,調整細胞濃度為 1×106個/ml,使用Annexin V-FITC/PI細胞凋亡檢測試劑盒依次加入5 μl Annexin V-FITC和10 μl PI,輕輕混勻,室溫避光孵育15 min,流式細胞儀檢測細胞凋亡。
1.8雙熒光素酶報告基因實驗 Targetscan在線預測發現PDCD4 3′-非翻譯區(UTR)上存在miR-181b-5p的結合位點,為進一步驗證PDCD4是否是miR-181b-5p的靶基因,按照LipofectamineTM2000使用說明書,將野生型PDCD4表達載體(3′-UTR-WT)和突變型PDCD4表達載體(3′-UTR-MUT)分別與miR-181b-5p mimic共轉染至HUVEC細胞,培養48 h后檢測熒光素酶活性,相對熒光強度=螢火蟲熒光強度/海腎熒光強度。
1.9統計學分析 采用SPSS20.0軟件進行獨立樣本t檢驗、單因素方差分析。
2 結 果
2.1高糖對HUVEC增殖和凋亡的影響 與對照組相比,高糖組HUVEC在培養至48 h、72 h時活性顯著降低,細胞凋亡率顯著升高(P<0.01、P<0.001)。見表1。

表1 高糖對培養不同時間HUVEC增殖和凋亡的影響
2.2高糖對HUVEC中miR-181b-5p和PDCD4蛋白表達的影響 與對照組相比,高糖組的miR-181b-5p呈顯著低表達,PDCD4 mRNA和蛋白表達水平顯著升高(P<0.05)。見圖1,表2。

圖1 高糖對HUVEC中PDCD4蛋白表達的影響
表2 高糖對HUVEC中miR-181b-5p和PDCD4蛋白表達的影響
2.3過表達miR-181b-5p對高糖誘導的HUVEC增殖、凋亡的影響 與高糖組相比,高糖+miR-181b-5p組miR-181b-5p的表達量顯著升高(P<0.001),HUVEC在培養至48 h和72 h時細胞活性顯著增加(P<0.05,P<0.01),細胞凋亡率顯著降低(P<0.01)。見表3。
表3 過表達miR-181b-5p對高糖誘導的培養不同時間HUVEC增殖、凋亡的影響
2.4抑制PDCD4蛋白表達對高糖誘導的HUVEC增殖、凋亡的影響 與高糖組相比,高糖+si-PDCD4組的PDCD4蛋白表達量顯著降低(P<0.01),48 h、72 h HUVEC的細胞活性顯著增加(P<0.05,P<0.01),凋亡率顯著降低(P<0.01)。見表4,圖2。
表4 抑制PDCD4蛋白表達對高糖誘導的培養不同時間HUVEC增殖、凋亡的影響

圖2 抑制PDCD4蛋白表達對高糖誘導的HUVEC中PDCD4蛋白表達的影響
2.5miR-181b-5p靶向調控PDCD4蛋白的表達 PDCD4 3′-UTR上存在miR-181b-5p的結合位點(圖3);將miR-181b-5p mimic與PDCD4 3′-UTR-WT質粒共轉染后細胞熒光素酶活性顯著下降(P<0.001);與PDCD4 3′-UTR-MUT質粒共轉染后細胞熒光素酶活性無明顯變化(P>0.05),見表5。與miR-NC組(0.45±0.05)相比,miR-181b-5p組HUVEC的PDCD4蛋白表達(0.21±0.03)顯著下調(P<0.05),與anti-miR-NC組(0.41±0.04)相比,anti-miR-181b-5p組HUVEC的PDCD4蛋白表達(0.76±0.08)顯著上調(P<0.05)。見圖4。
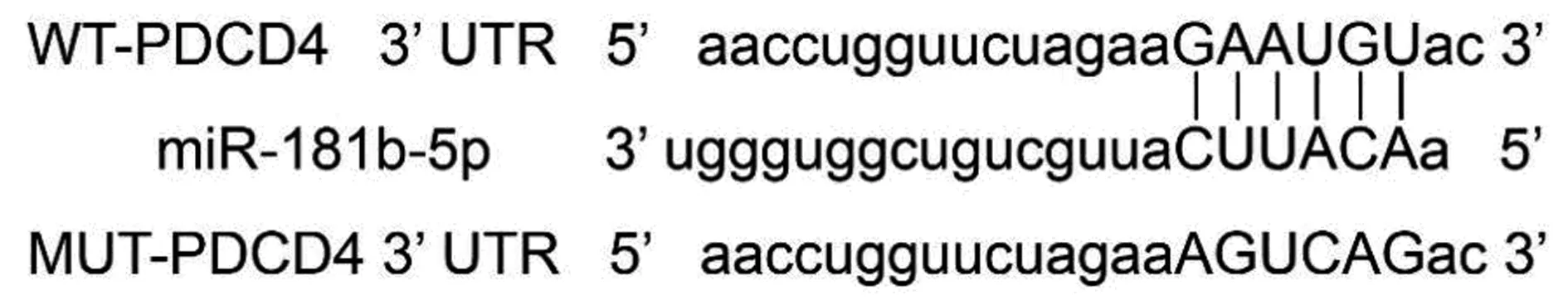
圖3 PDCD4的3′UTR中含有與miR-181b-5p互補的核苷酸序列
表5 雙熒光素酶報告實驗

1~4:miR-NC組,miR-181b-5p組,anti-miR-NC組,anti-miR-181b-5p組圖4 miR-181b-5p調控PDCD4蛋白的表達
2.6過表達PDCD4逆轉了過表達miR-181b-5p對高糖誘導的HUVEC細胞凋亡的影響 將miR-181b-5p和PDCD4共轉染至HUVEC細胞,并用葡萄糖濃度為33 mmol/L的培養基培養。與高糖組相比,培養48 h、72 h時高糖+miR-181b-5p組HUVEC細胞的細胞活性顯著提高,細胞凋亡率顯著降低(均P<0.05);與高糖+miR-181b-5p+pcDNA組相比,培養48 h、72 h時高糖+miR-181b-5p+pcDNA-PDCD4組HUVEC細胞的細胞活性顯著降低,細胞凋亡率顯著升高(均P<0.05)。提示過表達PDCD4逆轉了上調miR-181b-5p表達對高糖誘導的HUVEC細胞增殖和凋亡的影響。見表7。
表7 過表達PDCD4逆轉過表達miR-181b-5p對高糖誘導的培養不同時間HUVEC細胞增殖及凋亡的影響
3 討 論
糖尿病患者的死亡與其并發癥和合并疾病直接相關,據統計〔9〕,80%的糖尿病患者死于心血管疾病。研究表明,糖尿病患者心血管疾病的發病機制與血管內皮損傷有關,血管內皮損傷是糖尿病血管病變的重要機制〔10,11〕。因此,尋找有效的基因靶點保護VEC對預防糖尿病血管病變具有重要意義。
miRNA與靶基因之間的關系是復雜的,一個miRNA可以調控許多靶基因的表達,一個靶基因也可以被許多miRNA所調控,miRNA在各個領域的功能不斷被認識〔12〕。研究表明,miRNA參與調節VEC的增殖、遷移和凋亡等過程,進而影響VEC介導的血管生成,在心血管疾病的發生過程中起到調控作用〔13~15〕。在腫瘤細胞生物學功能上關于miR-181b-5p的研究較多,如miR-181b-5p在卡波西肉瘤組織中的表達水平高于癌旁組織,miR-181b-5p可促進卡波西肉瘤細胞增殖,抑制其凋亡〔16,17〕。在胃癌細胞中miR-181b-5p的表達呈顯著升高水平,促進細胞增殖,抑制細胞凋亡,miR-181b-5p在胃癌中可能起到癌基因的作用〔18〕。miR-181b在動脈粥樣硬化患者血清中低表達,可能通過阻斷VEC中核轉錄因子-κB信號途徑和抑制血管平滑肌細胞的增殖和遷移起到抗動脈粥樣硬化的作用〔19〕。本研究顯示,高糖誘導的HUVEC中miR-181b-5p表達明顯降低,細胞增殖能力降低,細胞凋亡率明顯升高。上調miR-181b-5p的表達,可促進HUVEC的增殖,降低細胞凋亡率。
PDCD4是一種細胞凋亡基因,在正常組織中廣泛表達,起到調節細胞程序性死亡的功能,也作為一種抑癌基因,通過抑制基因的轉錄和翻譯來抑制腫瘤的生長〔20〕。有研究發現,血小板源性生長因子刺激的血管平滑肌細胞中PDCD4顯著下調,過表達PDCD4可抑制血管平滑肌細胞增殖〔21〕。研究發現,PDCD4是VEC中miR-155的重要靶基因,miR-155通過調節PDCD4表達參與了高糖誘導的VEC的凋亡過程〔22〕。本研究提示,過表達PDCD4逆轉了上調miR-181b-5p表達對高糖誘導的HUVEC增殖和凋亡的影響。
綜上,miR-181b-5p在高糖誘導的HUVEC中呈低表達,上調miR-181b-5p可促進HUVEC增殖,抑制其凋亡,其機制可能與miR-181b-5p靶向調控PDCD4有關,為高血糖引起的血管病變提供了新的治療靶點。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
