除草劑草甘膦和氯磺隆對植物毒性效應的研究進展
2020-10-16 10:33:28王淑楠任浦慧解靜芳
山西農業科學 2020年10期
王淑楠,任浦慧,解靜芳
(山西大學環境與資源學院,山西太原030006)
除草劑在農業生產過程和其他領域使用廣泛,且種類繁多。目前常見的種類有化學、生物和轉基因除草劑。其中,化學除草劑的歷史可追溯到20世紀40年代初,2,4-D是其中最早使用的,此后其他除草劑的使用也逐漸增多,施用面積遠超其他類型農藥(如殺蟲劑和殺菌劑),截至2000年,化學除草劑在我國的施用面積已達到種植面積的40%[1]。化學除草劑的種類繁多,主要類型有:苯氧羧酸類、芳氧苯氧丙酸酯類、二硝基苯胺類、環己烯酮類、三氮苯類、酰胺類、取代脲類、二苯醚類、磺酰脲類、氨基甲酸酯類以及有機磷類等。
草甘膦是一種高效、廣譜、殘留低、非選擇性的芽后除草劑。自20世紀70年代美國孟山都公司開發以來,現已成為全世界使用量最大的除草劑之一;氯磺隆是一種內吸、超高效、用量少的磺酰脲類除草劑。它是在20世紀80年代初由美國杜邦公司開發的,因其高生物活性對后茬作物藥害嚴重。
除草劑的施用對于農作物有沒有危害,對植物的光合作用、抗氧化系統和代謝過程等有哪些具體影響,對人體健康是否有不良影響,已成為公眾關注的熱點。本文在概述2種除草劑的性質、作用機制以及使用現狀基礎上,就其對植物尤其是農作物的生態毒性效應的研究進展進行了綜述,以期為除草劑的安全管理提供科學依據。
1 草甘膦、氯磺隆的除草作用機制
草甘膦(Glyphosate,PMG)用于農田除草,其每公頃施用量為6.7~13.4 g。主要除草機制是通過抑制植物葉綠體中的5-烯醇丙酮莽草酸-3-膦酸合成酶(EPSP)的活性從而誘導莽草酸的合成。同時EPSP的抑制使得參與植物細胞合成的芳香族氨基酸(色氨酸、酪氨酸和丙氨酸)和次生代謝物(類黃酮、木質素、泛醌、植物保護素等)減少,并且能夠引起植物機體內代謝紊亂以及蛋白質合成受阻,最終導致綠色植物的生長受到抑制。但EPSP只存在于綠色植物中,因此草甘膦只對綠色植物有作用,對其他生物基本不會造成影響[2-4]。草甘膦還能夠快速進入植物根部并對其造成損傷,通過降低根部蛋白質和核酸的合成,導致ATP水平增高從而抑制生物合成過程[5]。除此之外,草甘膦還會影響植物葉肉細胞葉綠體的亞顯微結構,造成葉綠體和內質網發生腫脹,最終加速植物的老化[6]。
氯磺隆(Chlorsulfuron,CSF)作為一類超高效的除草劑,每公頃施用量為0.268~4.690 g[7],僅是草甘膦用量的4%~35%。氯磺隆的除草機制是通過抑制植物體內的乙酰乳酸合成酶(ALS),使支鏈氨基酸BCAAs(纈氨酸、亮氨酸、異亮氨酸)的合成受到抑制,從而導致細胞分裂期間DNA的合成受到破壞,干擾植物細胞的有絲分裂過程,使分裂細胞數量下降,最終導致植物枯萎死亡[8]。草甘膦和氯磺隆2種除草劑的基本性質如表1所示。
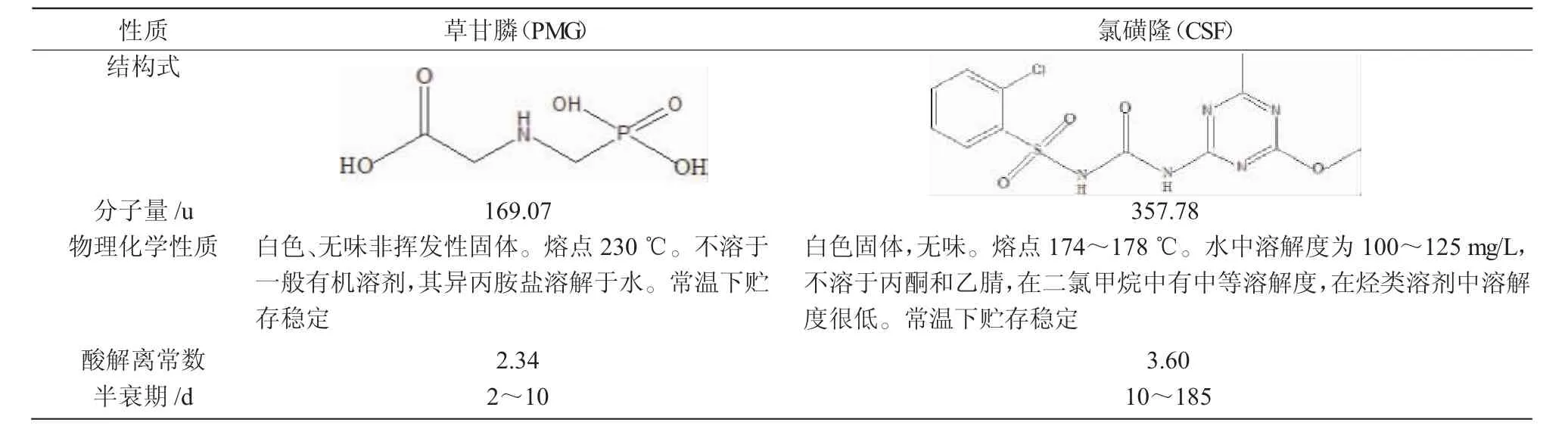
表1 草甘膦和氯磺隆的基本性質
本研究將從草甘膦和氯磺隆2種除草劑的使用現狀、土壤殘留現狀及對植物的毒性影響等方面展開綜述,并分析二者作用于植物的毒性效應、特別是導致植物代謝物變化及代謝途徑失衡的異同。
2 草甘膦和氯磺隆的使用現狀
2.1 草甘膦
草甘膦屬于有機磷除草劑,是美國最常用的除草劑之一[9]。據美國農業部門統計,1974—2014年美國共使用了137萬t草甘膦,其農田單位面積平均施用量約為全球的2倍[10]。草甘膦使用范圍廣,不僅適用于玉米、油菜和棉花等農作物,還可用于果園、工業園區、公路及鐵路旁等。我國1995年在非農田地區草甘膦使用量超過16 000 t,而農田用量是非農田的3倍多,高達51 000 t[11]。據統計,2012年國內草甘膦生產量約占全球總量的70%,2012—2014年產能由65萬t躍升為85萬t,且每年以8%~10%速度增加[12]。
2.2 氯磺隆
磺酰脲類除草劑因其活性高、用量低、殺草譜廣和選擇性強等優點在大麥和小麥等農作物生產地區被廣泛應用[13]。磺酰脲類除草劑品種繁多,包括甲磺隆、煙嘧磺隆、氟嘧磺隆和氯磺隆等。其中,氯磺隆是磺酰脲類除草劑中首先被開發利用的除草劑。氯磺隆所屬的磺酰脲類除草劑在美國使用最廣泛,其次是中國、歐洲和日本。在美國,每年約有30 t磺酰脲類除草劑用于小麥田除草[14],2003年歐洲共使用了221 t磺酰脲類除草劑[15]。2007年我國有10 t的氯磺隆用于小麥田除草[16],2009年國內磺酰脲類除草劑銷售量占所有除草劑的12%,僅次于草甘膦[16]。考慮到氯磺隆對農產品生產安全以及生態環境安全的影響,我國農業部規定自2015年12月31日起禁止銷售和使用氯磺隆[17]。
3 草甘膦和氯磺隆在土壤中的殘留現狀
3.1 草甘膦
草甘膦屬于一種有機酸,長期殘留會造成土壤酸化等一系列問題[18]。草甘膦在土地中的利用率只有30%,其殘留給土壤造成了許多潛在的風險[19]。研究表明,草甘膦施用后的林場土壤pH值下降,容重降低,結構發生改變,有機質、含氮量和自然含水率等增加[20]。并且低劑量的草甘膦還會造成土壤各項鹽化和堿化指標的上升[21-22]。施用過草甘膦的西班牙森林,在其20~35 cm深的土壤中均檢測出草甘膦殘留,并發現草甘膦在30 cm深處的土壤降解速率顯著低于表層土壤降解速率[23]。在1999—2002年,丹麥研究人員發現草甘膦在土壤中有吸附能力,在深層土壤中依然會發生移動和淋溶的現象[24]。在北京昌平區某蘋果園測得土壤中草甘膦施用后第28 d殘留量為0.073 mg/kg,低于我國規定的草甘膦最大殘留限量標準值(MRL)0.1 mg/kg[25]。
3.2 氯磺隆
氯磺隆因其較強的水溶性,進入土壤后會造成地下水污染。氯磺隆雖然在田間使用量不高,土壤中殘留微量,但因其殘留期和半衰期較長,尤其在堿性土壤中降解十分緩慢[26],容易對一些敏感的當茬和后茬作物造成危害,影響糧食安全和產量。研究表明,一般農藥僅有10%~30%被作物所利用,其余將進入土壤環境中,繼而改變土壤微生物群落數量和組成[27-28],但氯磺隆主要存在于表層土壤[29]。國外許多地區要求在施用氯磺隆1 a后才可種植作物[30]。美國規定磺酰脲類除草劑在水稻中的MRL為0.05 mg/kg,日本規定磺酰脲類除草劑的MRL為0.02mg/kg[31],我國制訂的食品安全國家標準中該農藥在農作物中的MRL為0.1 mg/kg[32]。
4 草甘膦和氯磺隆對植物的毒性效應
4.1 草甘膦和氯磺隆對植物生長的影響
國內外研究顯示[33-38],草甘膦和氯磺隆對一些農作物生長的影響,經常用萌發率、根長、芽長及生物量等來指示,在一定濃度下對植物的生長多呈抑制作用。
4.1.1 草甘膦 草甘膦在除草的同時,會導致作物的生長受抑。國內外諸多研究顯示,植株的總高度、總生物量、幼苗的根長和萌發率均隨草甘膦劑量的增加而逐步降低,植株死亡率上升。例如,經草甘膦處理后的品種為GR的大豆,其總高度和總生物量降低[33];洋蔥(Amaryllidaceae)在 500 mg/L草甘膦的脅迫下,毒性最強,其發芽率、根長和總生物量均有不同程度的下降[34]。
4.1.2 氯磺隆 土壤中的氯磺隆對植物根部起顯著抑制作用,通過抑制作物的根長、芽長和根質量等,影響作物的生產安全。經408 g/kg氯磺隆處理后的品種為ARLE3的春小麥,其根長和芽長下降[35]。研究表明,施用35 mmol/L氯磺隆后的大麥幼苗(Hordeum vulgareL.),其根部干質量顯著降低[36]。氯磺隆對豌豆(Pisum sativum)、菜豆(Phaseolus vulgaris)和蠶豆(Vicia faba)幼苗根部生長均起抑制作用,主要表現在根尖細胞膜損傷和根冠脫落[37]。研究表明,我國江蘇省農田施用劑量為2 g/hm2的氯磺隆后,小麥幼苗生長受到抑制,鮮質量水平降低,最終導致作物產量下降[38]。
4.2 草甘膦和氯磺隆對植物光合作用系統的影響
光合作用是植物的基本生理代謝過程,光合作用的增加有利于農作物質量和產量的提升。經草甘膦和氯磺隆處理后的植物光合速率下降,對葉綠素熒光參數有顯著影響,且會通過多種途徑影響植物的光合作用。研究表明[39-44],2種除草劑超過一定濃度范圍后會破壞植物的光合組織,減少光合色素含量。且草甘膦和氯磺隆均會抑制對光合作用有重要影響的氣孔反應,導致光合速率下降和氣孔導度下降(氣孔阻力上升)相伴發生。
4.2.1 草甘膦 草甘膦及其代謝產物氨甲基磷酸(AMPA)均會抑制植物的光合作用,主要表現在影響葉綠素的合成、葉綠素熒光參數異常和破壞C3循環等。有研究表明,草甘膦和AMPA會導致柳樹(Salix miyabeana)品種SX64的凈光合速率下降、葉綠素熒光參數(如PSⅡ最大光量子效率(Fv/Fm)和光合電子傳遞速率(ETR))不同程度地下降,葉綠素含量也呈下降趨勢[39]。CO2同化是光合作用的一個重要過程,C3是其中最基本的一個循環途徑。草甘膦通過破壞甜菜的C3循環從而幾乎完全抑制了甜菜的光合作用[40]。抗草甘膦RR2大豆幼苗施用10%水劑的10 800 g/hm2草甘膦后,其蒸騰速率和氣孔導度明顯下降[41]。
4.2.2 氯磺隆 0.001 mg/kg的氯磺隆土壤殘留對玉米(Zea maysL.)幼苗的葉綠素含量、葉綠素熒光參數均產生負面影響[42]。葉綠素熒光成像系統的結果顯示,葉綠素熒光參數(如PSⅡ最大光量子效率(Fv/Fm)和光化學淬滅(qP)等)呈下降趨勢,非光化學淬滅(NPQ)呈上升趨勢,這些參數的變化均表明玉米幼苗的光合作用受到抑制且激發了自身的保護機制[42]。葡萄(Vitis viniferaL.)每周連續3次施用田間劑量1%的氯磺隆后,葉片嚴重損傷:葉片面積50%褪綠,葉片凈光合速率下降25%,氣孔阻力增加59%[43]。但也有研究表明,氯磺隆處理時間較短(1~6 h)時,植物的光合作用和呼吸作用不會出現抑制作用[44]。
4.3 草甘膦和氯磺隆對植物抗氧化系統的影響
植物(主要是農作物)對草甘膦和氯磺隆的脅迫均出現不同程度的氧化損傷,并通過自身活性氧(ROS)清除機制,清除自身ROS,以提高自身的耐受性[45]。超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)是植物體ROS系統中的重要保護酶。草甘膦和氯磺隆的脅迫均造成農作物中的CAT活性先升后降或短暫維持,不同的是,草甘膦會導致SOD和POD活性先升后降,而氯磺隆則會導致這2種酶活性逐漸下降。草甘膦和氯磺隆在植物體內均會引起丙二醛(MDA)的含量逐漸增加或維持一段時間后下降。除此之外,還會引起植物的其他氧化應激反應和脂質過氧化損傷。
4.3.1 草甘膦 草甘膦脅迫會導致植物體內ROS的積累,進而產生氧化損傷。膜脂過氧化致膜通透性增加、膜結構和功能受損,是造成植物氧化損傷的主要原因之一[46]。用草甘膦對玉米幼苗進行處理,隨著草甘膦劑量(1.25~20.00 mg/kg)的增加,其體內抗氧化酶(SOD、CAT和POD)活性先上升后下降,但是MDA含量逐漸上升;低劑量的草甘膦(1.25~5.00 mg/kg)誘導植物自身的防御機制發揮作用,但高劑量的草甘膦(5~20 mg/kg)會減弱植物自身保護系統,使體內活性氧增加,從而增加質膜通透性,使酶的活性下降[47]。有研究表明,水培處理時使用不同濃度(0.1、0.3、1.0 mol/L)的草甘膦處理苦蕎(Fagophyrum tataricum)幼苗后,其MDA含量增加,膜脂過氧化水平上升及超氧自由基O2-·產生速率增加[48]。經草甘膦處理后的水稻(Oryza sativaL.cv.Dongjin)幼苗葉片中,檢測到大量H2O2的積累和硫代巴比妥酸反應產物(TBARS),表明植物體內進行了氧化應激和脂質過氧化反應[49]。
4.3.2 氯磺隆 氯磺隆同樣會導致植物的氧化損傷。菠菜(Spinacia oleraceaL.)幼苗在氯磺隆劑量分別為 0.1、0.3、1.0、3.0、5.0 μg/kg脅迫下,其 SOD 和POD活性均顯著增高,且隨著濃度的增加SOD和POD活性的增速下降,CAT活性表現為先升后降趨勢,當氯磺隆劑量達到3.0 μg/kg時活性下降;與對照相比,MDA含量在0.1 μg/kg脅迫下沒有變化,0.3、1.0 μg/kg處理下逐漸上升,當氯磺隆劑量逐漸增大到5.0 μg/kg時,其含量明顯下降,表明在此濃度下菠菜幼苗受到了氧化損傷及細胞膜受損[50]。在氯磺隆劑量為0.1 μg/kg時,水稻(揚稻7號)和大豆(中黃13號)幼苗的谷胱甘肽轉移酶(GST)活性均明顯高于對照,而GST的活性增加是由于植物細胞清除體內的活性氧所致[51]。扁豆(Lens culinarisMedik)和小麥(Triticum aestivumL.)施用氯磺隆后,檢測出γ-氨基丁酸(GABA)和MDA積累,GABA是一種調節植物內部碳氮平衡和防止氧化脅迫的物質[52]。綜上所述,氯磺隆對作物抗氧化系統的影響要比對蔬菜敏感。面對不同種類和品種的植物,2種除草劑毒性效應的強度不同,或者可以說,不同植物對草甘膦和氯磺隆的敏感性各異。
4.4 草甘膦和氯磺隆對植物礦物質元素的影響
礦物質元素作為植物體的重要組成成分,用以維持其正常的生理和生命活動。植物一般通過根部從土壤中吸收礦物質元素。草甘膦和氯磺隆對不同植物的礦物質吸收影響各異,有些起促進作用,但大多數呈抑制作用。
4.4.1 草甘膦 草甘膦分別作用于作物和果樹上,二者對其營養元素的吸收影響各異。如草甘膦作用于大豆后,其葉片含氮量下降,產量降低[53-54]。而連續3 d施用4.97 g/L草甘膦后的桃樹(品種為霞暉8號),其葉片中礦物質元素N、P、K和Fe元素含量顯著升高(分別增加10.42%、11.92%、21%和27.26%)。這可能是由于草甘膦將果園的大多數雜草滅掉后避免了雜草與桃樹競爭肥力所致[55]。
4.4.2 氯磺隆 有關氯磺隆影響植物對礦物質吸收的研究較少,但有研究表明,氯磺隆同樣能阻礙植物對一些微量元素的吸收。如氯磺隆會干擾小麥根部生長從而對微量營養素運輸系統產生影響并進一步抑制對微量元素(Zn、Cu和Mn)的吸收[56]。
4.5 草甘膦和氯磺隆對植物激素分泌的影響
植物激素是調節植物生長發育的有機物質。植物激素作為一類植物體內的微量信號分子,可調節植物生長發育過程且對環境應答有重要意義[57]。赤霉素可刺激植物葉和芽的生長,草甘膦和氯磺隆均會對植物體內用于調節植物高度的赤霉素的合成有抑制作用。
4.5.1 草甘膦 植物激素分泌會受到除草劑草甘膦的干擾。研究結果顯示,草甘膦是細胞色素氧化酶P450的一種抑制劑,而P450參與植物激素赤霉素的合成,所以草甘膦是通過抑制P450最終影響植物激素赤霉素的合成,從而抑制植物生長[58-59]。也有研究表明,噴施188 g/hm2的草甘膦能夠誘導橡樹芽(品種為熱研7-33-7)中的脫落酸和玉米素含量增加[60],而大量脫落酸和玉米素會導致植物生長緩慢,最終引發植物體內細胞的死亡。
4.5.2 氯磺隆 氯磺隆可導致植物內部激素的變化,從而使植物生長受到抑制。使用0.1μg/kg的氯磺隆處理水稻(揚稻7號)和大豆(中黃13號)后,2種植物均出現脫落酸含量升高、赤霉素含量下降;脫落酸在抗逆條件下迅速增加,加快植物器官脫落,促進休眠;赤霉素的減少則抑制植物生長[51]。
4.6 草甘膦和氯磺隆對植物代謝物變化和代謝途徑的影響
從代謝組學的角度來看,代謝物處于植物系統末端,反映的是已經發生的生物學事件。通過觀察植物受到脅迫后代謝物的變化、植物體內代謝途徑的異常,可以更加深入了解植物體內生理病理的狀況。利用代謝組學分析更能反映出植物體受到環境脅迫后最終的整體生理狀態。
4.6.1 草甘膦 草甘膦作用于不同植物或作物,劑量不同,產生的效應各異。較低劑量的草甘膦會促進一些植物(如巨桉和加勒比松等)生長,并減少植物體內莽草酸的積累[60]。低劑量的草甘膦也可增強甘蔗的蔗糖積累[61-62]。但多數情況下隨草甘膦脅迫劑量的增大,導致植物或作物的代謝物變化和代謝途徑失調。如經草甘膦處理后的擬南芥(Arabidopsis thaliana),利用GC-MS對其初級代謝物進行分析,結果表明,其體內精氨酸、谷氨酰胺、莽草酸和β-丙氨酸等水平顯著增高[63]。國外研究發現,對富含類黃酮的苦蕎,施用不同濃度(0.1、0.3、1.0 mol/L)的草甘膦連續處理3 d后,其類黃酮含量分別下降了58.1%、65.8%和75.6%[48],表明次生代謝產物類黃酮代謝途徑受抑。草甘膦作用于玉米幼苗,隨草甘膦濃度不斷升高,玉米幼苗體內的莽草酸含量增加,細胞內EPSP合成受到抑制,從而導致莽草酸代謝途徑紊亂;部分有機酸(如檸檬酸和蘋果酸)含量增加,抑制三羧酸途徑(TCA循環)從而影響細胞的呼吸作用;抗逆物質(如腐胺、酪胺和天冬酰胺等)含量增加,表明草甘膦濃度較大時會對作物造成毒性作用[64]。
4.6.2 氯磺隆 氯磺隆可致蔬菜和作物幼苗代謝物發生變化、多條代謝途徑異常。例如,氯磺隆處理后的菠菜和玉米幼苗,其腐胺、肌醇均積累[65-66]。腐胺能調節植物的生長發育,提高植物的抗逆能力。肌醇是促進植物細胞壁合成的重要物質,也用于提高植物的抗逆能力。施用氯磺隆后的玉米幼苗,其葉片中游離氨基酸含量增加63%,激活蛋白質分解代謝[67];在逆境情況下,植物體內游離氨基酸的積累是用于抵抗外界環境的脅迫。玉米幼苗的多種糖類代謝物質積累(如蔗糖和鼠李糖),導致作物能量代謝異常及細胞壁合成受阻[68]。研究表明,使用氯磺隆處理向日葵(Helianthus annuusL.)和豌豆幼苗后,阿魏酸和肉桂酸含量增加[69-70]。土壤中殘留的氯磺隆影響向日葵中支鏈氨基酸(纈氨酸和異亮氨酸)的合成,且蘋果酸和檸檬酸水平發生變化,從而抑制TCA途徑達到抑制植物的呼吸作用[69]。有研究表明,氯磺隆通過抑制BCAAs合成的除草機制,會增加莽草酸代謝途徑中次級代謝物奎寧酸鹽的合成,因奎寧酸鹽既可以參與植物的生長發育和抗逆性,也在植物體內有抗氧化作用[70-71],所以氯磺隆會導致莽草酸代謝途徑紊亂。
綜上所述,2種除草劑均可導致植物或作物體內代謝物種類變化、多條代謝途徑紊亂。草甘膦主要導致莽草酸代謝途徑發生紊亂,而氯磺隆主要抑制支鏈氨基酸的合成。但2種除草劑對代謝途徑的影響也存在一些相似之處:如均能抑制與植物呼吸作用有關的TCA循環[72-74];均可導致莽草酸代謝途徑關鍵酶(DAHPS)的活性增加[70-75],次級代謝物奎寧酸鹽含量升高,導致該代謝途徑發生紊亂;用草甘膦和氯磺隆分別處理曼陀羅后,其PMT的mRNA轉錄降低,植物體內托烷生物堿(托品酮、φ-托品堿和托品堿)、苯丙氨酸含量均降低,而精氨酸和鳥氨酸的水平增加[76]。
5 小結
除草劑的生態毒性效應,是諸多研究者經常關注的熱點問題。本文在對草甘膦和氯磺隆2種除草劑的除草作用機制、結構性質、使用現狀、土壤殘留現狀進行概括的基礎上,重點對其毒性效應進行了綜述。
草甘膦和氯磺隆的結構性質不同,除草機制各異。草甘膦的除草機制主要通過抑制植物體內EPSP活性導致莽草酸和芳香氨基酸積累;氯磺隆則通過抑制ALS使BCAAs合成下降。
草甘膦和氯磺隆使用范圍廣、使用量大;氯磺隆在我國已被禁用。曾經大量使用過草甘膦和氯磺隆的土壤,會形成不同程度的殘留量,這些土壤殘留會對植物(尤其是農作物)造成各方面的毒性效應。
草甘膦和氯磺隆脅迫可導致植物各生長指標下降,如萌發率、根長、芽長和生物量等。2種除草劑均會抑制植物的光合作用,超過一定濃度范圍后可導致光合色素含量、凈光合速率和氣孔導度下降以及葉綠素熒光參數發生變化。
草甘膦和氯磺隆脅迫可造成植物體內ROS損傷,均可引起SOD、POD、CAT和MDA發生不同程度的變化。此外,還會造成植物對礦物質元素的非正常吸收,影響植物激素的合成。
2種除草劑均可致植物或作物體內多種代謝物種類變化、多條代謝途徑紊亂。但二者既有差異,也有相似之處。草甘膦主要導致莽草酸代謝途徑紊亂,氯磺隆主要抑制支鏈氨基酸的合成。相似之處是均可抑制TCA循環受阻和莽草酸代謝途徑紊亂。
6 展望
綜上所述,草甘膦和氯磺隆對植物的毒性效應十分復雜,明晰其對植物尤其是農作物生長階段的致毒機制十分重要。本綜述對國內外目前的研究結果進行歸納總結,在以下幾個方面需開展更深入的研究:一是許多研究主要以水培種植在實驗室中進行,其研究條件可能與實際污染情況相差較大,研究結果可能有一定的差異性,因此,應更加注重研究條件和環境的選擇;二是研究所選擇的毒性指示應不僅包括植物簡單的生長生理指標,還應利用基因組學、蛋白組學、轉錄組學和代謝組學等方法的結合使用,研究農作物受到除草劑脅迫時的靈敏反應,增加在分子水平上的證據,并從這些毒理學指標中尋找出污染土壤毒性的敏感生物標記物;三是許多研究只針對植物的幼苗階段,應加強研究2種除草劑對農作物整個生命周期的影響,有利于全面評價農作物的安全風險以及為人體的健康風險評價提供基礎理論依據;四是應加強植物對草甘膦和氯磺隆耐性和解毒方面的研究,以培育出含草甘膦和氯磺隆耐性基因的植物,從而提高植物的耐受力,為土壤有機污染的生態修復提供科學依據;五是對于草甘膦和氯磺隆對植物的毒性效應,在各種生理生化反應方面已經做了大量工作,但現階段土壤污染物存在多樣性,土壤污染類型復雜,僅僅關注單一有機物的污染是不夠的,應更加關注污染物的復合作用對生物的毒性效應研究。
猜你喜歡
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
現代農業(2016年5期)2016-02-28 18:42:36
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
營銷界(2015年23期)2015-02-28 22:06:18
