喀斯特地區造林對土壤團聚體穩定性及微生物碳代謝活性的影響
2020-10-21 07:32:38喻國軍謝曉堯
水土保持研究 2020年6期
關鍵詞:差異
喻國軍, 謝曉堯
(1.貴州大學 計算機科學與技術學院, 貴陽 550001; 2.貴州師范大學 信息與計算科學重點實驗室, 貴陽 550001)
對于全球生態環境而言,大氣、陸地及海洋是其主要生態系統,此外還包括濕地、土壤、水域等多種生態子系統,其影響要素不僅具有多樣性,還體現著明顯的復雜性[1-2]。立足于陸地生態而言,其不僅包含森林、草地等子系統,還包括土壤、微生物等多種生態子系統;由于土壤直接關乎作物生長發育[3],同時也是人類生產生活的關鍵載體,因此其重要地位不言而喻,不僅在生態系統中尤為重要,而且在生產生活中起著突出的影響作用。作為土壤的關鍵構成部分,其團聚體不僅直接決定著土壤結構的穩定性[4],在降低水土流失方面效果突出,另一方面能夠有效固定有機碳成分,進而促進土壤肥力的保持及提升,能夠明顯改善土壤質量[5-6],經過大量實證分析可知,該指標能夠作為衡量土壤質量的一個關鍵指標。就目前而言,對于團聚體分類的標準并未能完全統一,但是主要借助于濕篩法[7-8],且其大小的分界點常常為250 μm,超過該界點則稱為大團聚體,否則稱為微團聚體。團聚體的大小對土壤孔隙度的制約效果不同,對土壤結構的影響效果具有較大差別,對于土壤養分的調節效應也不盡相同[9-10],此外,對于土壤的生化反應也產生不同程度的制約效果。近些年來不少學者開展了土壤團聚體的相關研究,主要集中在土壤穩定性及有機碳等方面,研究范圍多集中在農作物方面,尤其是耕作及施肥等因素的制約方面,但是對于森林土壤的研究相對較少[11-13]。森林資源不僅能夠創造一定的經濟來源,更為重要的是具有重要的生態意義,成為陸地生態的重要一環,在為植被生長發育提供場所的同時也促進了微生物等動物生長,成為多種能量交換的場所之一;在微生物的參與下,加之酶的作用,腐殖質等有機質能夠高效地被分解轉化成養分,促進了能量循環,關乎土壤理化特性,也成為衡量土壤狀況的關鍵指標之一,在分子生物學等技術不斷發展的情況下,微生物的研究也由單純的數量分布向多樣性研究轉變,不僅能夠對其物種及功能等方面加以探索,而且在遺傳多樣性方面的研究在不斷深化,尤其是在研究過程中脂肪酸分析法PLFA以及碳素分析法的廣泛利用,此外核酸分析的方法也運用較為廣泛,這些技術的發展顯著促進了微生物研究,從多個角度、多個層面開展相應分析研究[14],也利于開展土壤研究。
貴州喀斯特地區受制于地理位置分布和高原河流等影響,喀斯特地貌分布尤為突出,近些年來該地區呈現明顯的植被退化現象,土壤肥力和活性大幅下降,土壤穩定性下降,整體呈現顯著的生態退化。為了進一步改善生態環境,人工造林及生態育林等日益受到重視,并取得了較好效果[15-16]。腐殖質等有機質的高效降解及轉化離不開微生物的參與,此外酶也起著不可忽視的作用,從而成為養分循環的關鍵參與者之一,在土壤肥力和活性方面施加關鍵影響,促進植被對養分的吸收利用,在評價土壤狀況方面效果突出。不少學者對于該地區的植被演替與土壤之間的關系進行了探究,對于喀斯特地貌而言,人工育林在土壤養分保持方面效果突出,能夠有效改善土壤理化特性,通過土壤結構的改善來促進其質量的提升[17]。基于此,本文連續4 a研究喀斯特地區造林對土壤團聚體穩定性及微生物碳代謝活性的影響,以期有利于評估我國重大工程的生態效應。
1 材料與方法
1.1 研究區概況
選取貴州睛隆縣(東經105.18°,北緯25.44°),境內50%以上的面積屬于喀斯特地貌,高原峽谷均有廣泛分布,山高、谷深的特點尤為突出,山巒起起伏伏且坡度較大,形成了海拔差(1 400 m)。受地理位置制約,其亞熱帶季風氣候尤為突出,呈現明顯的雨熱同期分布特點,其氣候也具有突出的垂直分布差異,海拔較低的區域明顯干熱,而對于高海拔區域而言較為寒冷。該區域年均降雨不足1 700 mm,年均溫度約為15℃,年均日照1 600 h,雨熱同期;受喀斯特地貌影響,土層分布較為稀薄,土壤結構不夠穩定,制約著植被生長發育,直接影響了作物種植,因此該區域分布大量的草地、喬木等;此外,由于山巒起伏,受風化及巖溶等制約,常常發生自然災害[16-17]。
1.2 試驗設計
在造林地選取榿木林、杜仲林,還包括刺槐林、滇柏林和未造林地(CK,對照組)共5種林分,連續4 a長期定位觀測(2015—2018年4月)。為了保證取樣的一致性,所取樣的土壤坡度均小于5°,各林分處理分別設5個20 m×20 m的樣方,間距>15 m,每個樣方進行標記,長期采樣觀測,共計25個樣方。每個樣方重復采集5個樣品,每個樣品重復取3次,然后混合成一個樣品,采樣深度為0—10,10—20,20—30 cm的擾動土和原狀土樣。4 a共采集1 500個土壤樣品。所取土壤樣品分為兩部分,一部分自然風干后測量土壤微生物,另一部分原狀土為在風干過程中沿自然裂隙掰成直徑為1 cm左右大小的團聚體。團聚體的大小對土壤孔隙度的制約效果不同,對土壤結構的影響效果具有較大差別,對于粒徑分組采取濕篩法進行,然后選取風干土樣100 g,通過5 ml的純水進行浸潤,接下來進行過篩處理,整套篩子包含3種不同的篩目,不僅有53 μm篩,還有0.25,2 mm篩,同時對土樣進行濕篩處理,要求振幅達到3 cm,頻率為50次/min,從而獲取了4種粒徑的土壤團聚體,進行烘干處理后稱重,并計算相應的質量分數。
1.3 土壤微生物測定方法
微生物量碳、氮、磷采用氯仿熏蒸—浸提法測定[8-9];土壤呼吸采用靜態堿液吸收法[11]。
微生物對于凋落物分解效果突出,加之酶的參與進一步提升了分解效果,二者是息息相關的,離開了微生物,凋落物等腐殖質的降解將無從談起,這在土壤養分轉化過程中尤為關鍵,直接制約著土壤肥力及活性,因此,在土壤質量分析過程中微生物常作為關鍵指標加以分析。本研究對于微生物多樣性分析立足于Shannon和Simpson等指數[11]。對于Simpson指數計算而言,首先對相應的數據進行放大1 000倍的處理,這樣將不會出現負數的現象;對于微生物的碳源利用水平進行衡量過程中主要借助于AWCD值衡量法[8-9],即:
AWCD=∑[(Ci-R)/31]
(1)
H=-∑Pi(lnPi)
(2)
Pi=(Ci-Ri)/∑(Ci-Ri)
(3)
Ds=1-∑Pi
(4)
式中:AWCD為顏色平均變化率;Ci為第i個非對照孔吸光度;n為碳源種類數(n=31);R為對照孔吸光度;H為Shannon-wiener多樣性指數;Pi為第i個非對照孔吸光度與所有非對照孔吸光度總和的比值;Ds為Simpson優勢度指數。
1.4 數據分析
本試驗對于團聚體穩定性的評價主要通過以下兩方面進行:一方面借助于平均質量直徑MWD,另一方面借助于幾何平均直徑GMD,其計算公式如下[12-13]:
(5)
GMD=exp[∑Wi·lnXi]
(6)
式中:Xi為第i級的團聚體平均直徑( mm);Wi為第i級團聚體重量百分含量。
2 結果與分析
2.1 不同林地對土壤團聚體組成的影響
通過對圖1的分析不難發現,受到不同林地的制約,不同試驗分組獲取的大團聚體含量還是存在著明顯的差異,其中含量最高的是榿木林地,其次是杜仲林,而滇柏林地含量最低,但是整體而言其超過2 mm的團聚體均明顯高于對照組;從土壤剖面的角度來講,在土層深度不斷上升的情況下,大團聚體的含量呈現明顯的下降態勢。對于中間團聚體而言,不同林地并未帶來較大的差異影響,從土壤剖面的角度來講,在土層深度不斷上升的情況下,其含量并沒有較大差異,對于微團聚體來說亦是如此。對于黏團聚體而言,由于其粒徑一般低于53 μm,不同的林地類型對其產生的影響較為突出,從不同土層的土壤剖面來講,該類型團聚體含量最高的是滇柏林地,其次是刺槐林,而榿木林含量最低,但是整體而言該類型團聚體均明顯高于對照組。
通過對圖2的分析不難發現,對于土壤團聚體而言,其受到林地類型的影響較為明顯,尤其是對于榿木林、杜仲林地來講,其分布的大團聚體居多,占比超過了20%。對于刺槐林、滇柏林而言,其主要的團聚體不僅包括中間團聚體,還包括黏團聚體;而對于對照組來說,其黏團聚體居多,且占比接近于40%。
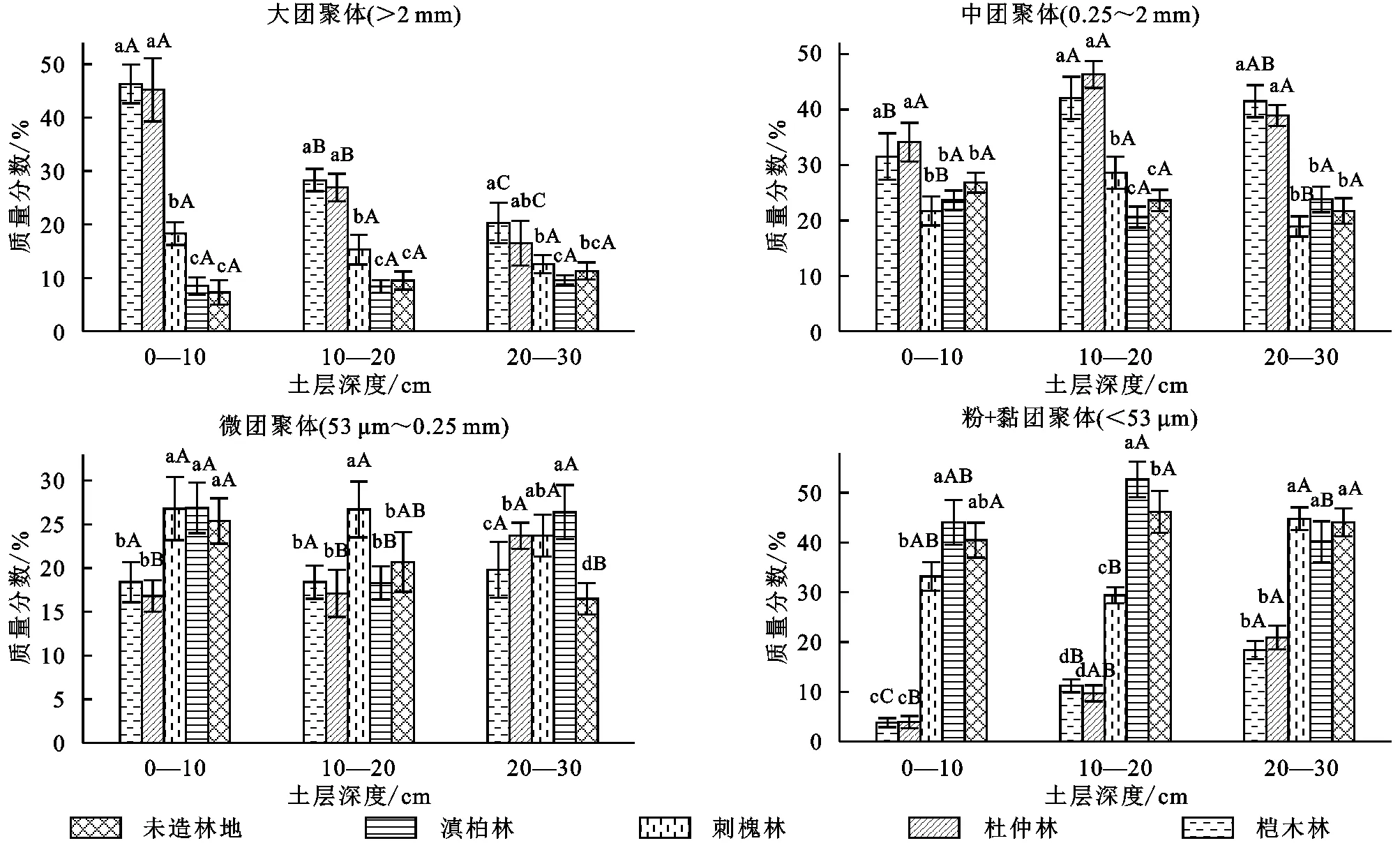
注:不同小寫字母表示同一深度不同林地在0.05水平差異顯著,不同大寫字母表示不同深度同一林地在0.05水平差異顯著,下同。
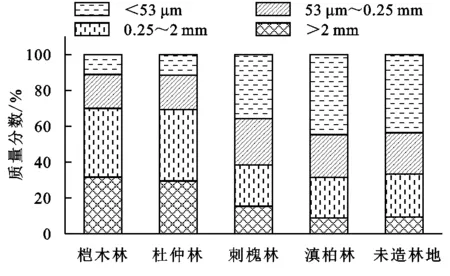
圖2 不同林地0-30 cm土層各粒徑團聚體質量分數平均值
2.2 不同林地對土壤團聚體穩定性的影響
作為土壤的關鍵構成部分,其團聚體不僅直接決定著土壤結構的穩定性[4],在降低水土流失方面效果突出,另一方面能夠有效固定有機碳成分,進而促進土壤肥力的保持及提升,能夠明顯改善土壤質量[5-6],經過大量實證分析可知,該指標能夠作為衡量土壤質量的一個關鍵指標。本試驗對于團聚體穩定性的評價主要通過以下兩方面進行:一方面借助于平均質量直徑MWD,另一方面借助于幾何平均直徑GMD,通過實證得知,其值越大說明其粒徑越大,這也表明其穩定性較高。從圖3分析不難發現,對于不同林地而言,在土層深度不斷增加的情況下,團聚體的穩定性呈現明顯的下降水平,而對于對照組來講恰好相反。從土壤剖面的角度來講,對于榿木林、杜仲林兩種林地而言,其MWD值大幅超過其他林地,對于GMD來說亦是如此;對于20—30 cm土層而言,其值差異明顯降低,從另一個角度來講表層土壤更易受到人類活動影響,這能夠從團聚體穩定性方面可以看出。從地表至30 cm土層,二者的變化態勢基本一致。通過對比分析得知,對于未造林而言,其團聚體的穩定性相對較差,且低于其他4種林地,其更容易在雨水的沖刷下被沖散,相反,開展育林之后其穩定性得到了明顯提升,能夠有效增強土壤穩定性。
2.3 不同林地對土壤微生物對碳源利用強度的影響
在微生物的參與下,加之酶的作用,腐殖質等有機質能夠高效地被分解轉化成養分,促進了能量循環,關乎土壤理化特性,也成為衡量土壤狀況的關鍵指標之一。不同的微生物群落分布,其碳源利用水平存在較大差異,在衡量過程中主要借助于AWCD分析法加以探究,通過對表1的分析不難發現,微生物碳源利用水平受到不同林地的顯著制約,其碳源利用率雖然有所差異,但是均高于對照組未造林地,且這種差異通過了顯著性差異,其中利用率最高的是榿木林地,其次是杜仲林地,且二者的差異并不太大,未能達到顯著水平。人工育林的分組下,其主要的碳源不僅包括碳水化合物,還包括羧酸類碳源,而最低的是胺類碳源。
2.4 不同林地對土壤微生物群落多樣性的影響
微生物群落多樣性指數可用來指示土壤微生物群落利用碳源的程度,根據培養第100小時的AWCD值計算土壤微生物群落的物種豐富度指數(H)、均勻度指數(E)、優勢度指數(Ds)和碳源利用豐富度指數(S)。由圖4可知,不同林地土壤微生物群落物種豐富度指數(H)、均勻度指數(E)和碳源利用豐富度指數(S)呈一致的變化趨勢,均表現為造林地顯著高于未造林地(p<0.05),而榿木林和杜仲林最高,二者之間差異不顯著(p>0.05);均勻度指數(E)呈相反的變化趨勢,造林地顯著低于未造林地(p<0.05)。

圖3 不同林地土壤團聚體平均質量直徑和幾何平均直徑
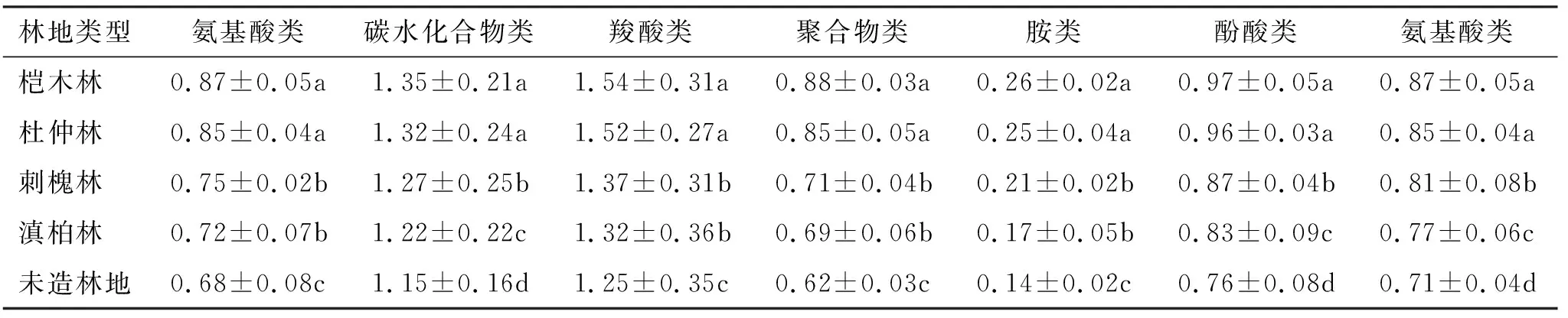
表1 不同林地土壤微生物對碳源利用強度

圖4 不同林地土壤微生物群落多樣性
2.5 不同林地對土壤呼吸及微生物代謝熵的影響
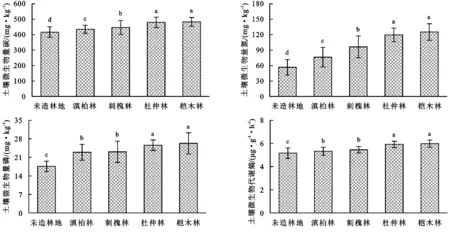
圖5為不同林地土壤微生物量和代謝熵,由圖可知,不同林地土壤微生量碳、微生物量氮、微生物量磷和微生物代謝熵影響顯著,其中不同林地土壤微生量碳、微生物量氮、微生物量磷和微生物代謝熵呈一致的變化趨勢,均表現為造林地顯著高于未造林地(p<0.05),而榿木林和杜仲林最高,二者之間差異不顯著(p>0.05)。
2.6 土壤微生物學特性與土壤團聚體的相關分析
相關性分析表明(表2),氨基酸類、碳水化合物類、羧酸類與大團聚體、中團聚體、MWD和GMD呈顯著正相關(p<0.05);物種豐富度指數(H)、優勢度指數(Ds)、碳源利用豐富度指數(S)與大團聚體、中團聚體呈顯著正相關(p<0.05);微生物量碳、微生物量碳、微生物代謝熵與微團聚體和粉微團聚體呈顯著正相關(p<0.05)。
注:同列不同小寫字母表示差異顯著(p<0.05)。

表2 土壤微生物學特性與土壤團聚體的相關分析
3 討 論
微生物能夠將腐殖質等有機質高效分解,從而為養分轉化奠定基礎,而微生物活性影響到期有機質分解效率,在此過程中常借助于平均顏色變化率AWCD進行衡量,這也是對碳源利用水平的衡量,借助于該指標不僅能夠分析微生物數量,還能夠分析其結構特點[18-19]。對于該指標而言,當其值越大的情況下,不僅說明細菌具有較大的分布密度,更能說明其活性更強,相反則表明其活性相對較差。通過本研究對比得知,對于4種林地而言,其碳源利用效率雖然有所差異,但是均高于對照組未造林地,且這種差異通過了顯著性差異,其中利用率最高的是榿木林地,其次是杜仲林地,且二者的差異并不太大,未能達到顯著水平。人工育林的分組下,其主要的碳源不僅包括碳水化合物,還包括羧酸類碳源,而最低的是胺類碳源。
對于微生物而言,其碳代謝的影響因素具有多樣性,且對于環境的變化具有較強的敏感性,對于其生態環境特征具有明顯的表征作用,其中尤為關鍵的指標不僅包括生物量碳,還包括微生物呼吸[20-21]。就生物量碳而言,其在有機質中具有明顯的高活性,成為土壤活性碳的主要構成,對于該碳物質而言,其不僅具有較快的周轉速度,而且環境敏感性較強,能夠有效反映土壤碳庫變化[22]。微生物群落分布不僅關系著能量的維系,而且制約著其對有機質的利用水平,這能夠通過代謝熵進行有效反映,此外,該指標能夠將微生物量碳及活性進行有機結合,從而對微生物活性加以充分體現[23-24];對于該指標而言,其值越小說明其在呼吸過程中對于碳的消耗量相對較少,而其細胞構造相關的碳會相應增多,其在維系土壤質量方面效果較為突出[25]。通過實地分析得知,無論是微生物量碳、量氮,還是其磷及代謝熵,其雖然受到不同林地的制約,但是其整體而言均超過對照組,且其差異達到了顯著差異,整體而言,其最高的是榿木林地,其次是杜仲林地,且二者的差異并不太大,未能達到顯著水平。對于人工育林而言,林木生長過程中會產生一定量的枯枝落葉等凋落物,在微生物的分解下其有機質含量逐漸增多,這對于微生物量碳的增加起著關鍵作用,同時能夠促進微生物呼吸和代謝活動,促進其活性的提升,而在此過程中需要一定的能量維系其代謝活動,對養分利用率得以明顯提升。
通過對比分析得知,對于不同林地而言,在土層深度不斷增加的情況下,團聚體的穩定性呈現明顯的下降水平。從土壤剖面的角度來講,對于榿木林、杜仲林兩種林地而言,其大團聚體明顯超過其他林地;從土壤剖面的角度來講,在土層深度不斷上升的情況下,大團聚體的含量呈現明顯的下降態勢。對于中間團聚體而言,不同林地并未帶來較大的差異影響,從土壤剖面的角度來講,在土層深度不斷上升的情況下,其含量并沒有較大差異,對于微團聚體來說亦是如此[26]。對于黏團聚體而言,由于其粒徑一般低于53 μm,不同的林地類型對其產生的影響較為突出,從不同土層的土壤剖面來講,該類型團聚體含量最高的是滇柏林地,其次是刺槐林,而榿木林含量最低,但是整體而言該類型團聚體均明顯高于對照組。為了探究團聚體穩定性與微生物方面存在何種聯系,本研究通過開展相關分析發現二者之間的相關性達到了顯著水平;對于土壤的碳水化合物及氨基酸來講,其不僅與大中團聚體息息相關,且與MWD,GMD密切相關,這種相關性均為正向,且通過了顯著性檢驗。而微生物量碳及代謝熵不僅僅與微團聚體息息相關,且與粉微團聚體密切相關,這種相關性均為正向,且通過了顯著性檢驗。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
