珠江三角洲濱海濕地土壤微生物群落多樣性與養分的耦合關系
2020-10-21 07:32:38孟妍君
水土保持研究 2020年6期
孟妍君, 秦 鵬
(1.桂林旅游學院, 廣西 桂林 541006; 2.桂林理工大學 博文管理學院, 廣西 桂林 541006)
從生態環境的角度來講,由于其構成要素眾多且存在顯著的交互作用,因此其復雜性和多樣性尤為突出,對于陸地生態而言更是如此,由于其直接作為人類生存發展的載體,動植物的生長發育受其直接制約,因此其具有復雜性的同時更突顯了其重要性[1-2]。立足于陸地生態環境,其中關鍵的構成部分離不開土壤和微生物,絕大多數植被的生長發育離不開土壤,在物質和能量交換過程中離不開微生物,從而形成了土壤—植物局地生態系統[3]。作為土壤生態的重要構成部分,微生物對于凋落物的分解發揮著無可替代的效果,同時在酶的參與下進一步提升了分解效果,二者是息息相關的,此外,環境因子也發揮著明顯的制約效應[4-5]。離開了微生物,凋落物等腐殖質的降解將無從談起,這在土壤養分轉化過程中尤為關鍵,直接制約著土壤肥力及活性,因此,在土壤質量分析過程中微生物常常作為關鍵指標加以分析。近些年來,微生物數量及分布特點等方面受到了大量學者的關注,并對此開展了長期的實地研究,加之研究方法及研究技術的不斷改進優化,對其研究更為深入,尤其是其結構及區系等方面,研究更為深入、多樣,其多樣性產生的影響進一步探究,尤其是隨著分子研究水平的不斷提升,微生物多樣性等方面的分析更為深入[6-8]。目前來講,對于微生物多樣性研究,首先要探究其物種分布,此外還需要對其功能及遺傳方面加以探究,在此過程中常用的研究方法不僅包括碳素利用法[9-10],還包括核酸分析法等多元化研究方法;有機質分解在土壤養分積累的過程中尤為關鍵,尤其是凋落物的降解,不僅與植被生長息息相關,同時與微生物活動關系密切,凋落物降解依賴于微生物分解[11-12],同時又為微生物新陳代謝提供能量支撐,因此來說,探究土壤與微生物及植被之間的關系具有重要的現實意義。
對于珠江三角洲濕地而言,其形成主要受制于珠江和海洋的交互效應,是入海口地帶的產物,由于其河口地質構造突出,受制于河口地帶水動力作用等,形成了突出的河口三角洲地帶,呈現出明顯的濕地生態環境[13-15],這也是當地較為獨特的地貌之一。隨著海平面的不斷上升,大量泥沙沉積于河口區域,從而不僅形成了濱海濕地,同時出現了大量的淺灘,出現了大量的顆粒沉積物,對于珠江三角洲生態產生了重要影響和變化,甚至出現了水下三角洲的獨特生態,其泥質沉積體現象較為突出[16-17]。然而,從20世紀80年代開始,在工業化進程的不斷發展過程中,珠江三角洲濕地生態發生了巨大變化,不僅濕地面積大幅下降,同時局地生態變化突出,呈現出明顯的退化現象,導致原有的濕地景觀衰退,還直接導致生物多樣性下降,破壞了其原有的生態平衡[18]。尤為突出的環境問題影響了當地的生態環境,對于濕地保護尤為不利,基于此,本研究從2015年開始進行連續3 a的實地觀測研究分析,長期觀測珠三角濱海濕地[互花米草(Spartinaalterniflora)濕地、堿蓬灘(Suaedaglauca)濕地、蘆葦(Phragmitescommunis)濕地、紅樹林(Rhizophoraapiculata)濕地],探究該區域的土壤狀況及微生物特點,揭示養分和微生物之間的內在關系,探究微生物群落分布特性,探尋其交互作用機理,從而為濕地保護提供有益參考和借鑒,為修復退化的濕地環境提供借鑒。
1 材料與方法
1.1 研究區概況
本研究選取珠江河口的淇澳島(113°36′40″—113°39′15″E,22°23′40″—22°27′38″N),該區域面積約為24 km2,受地理位置制約而具有較為突出的亞熱帶海洋氣候特點,通過近些年來該地區的氣象資料分析得知,年均氣溫達到22℃,即使是氣溫最低1月,其月均溫度依然能達到15℃,其最低溫為2.4℃。受地理位置及水文特點的影響,其潮汐現象呈現出半日潮的特點,且具有不規則的變化特點,潮漲潮落的規律難以找尋,潮差變化較大,容易受枯水季的影響,潮差一般不超過2 m。通過對其氣象數據進行分析得知,該區域受熱帶氣旋的影響較為常見,每年出現的次數甚至達到了12次,對于珠江口的影響較大。
1.2 樣品采集
2015年8月,在淇澳島選取蘆葦、紅樹林、互花米及堿蓬濕地,每種濕地選取3個重復樣地(每個樣地面積為50 m×50 m),每個樣地長期布設15個2 m×2 m的重復樣方,共計180個樣方,連續觀測3 a。連續3 a在每個樣方采集土壤混合樣品,采集深度為20 cm,采樣四點混合法采集約為2 kg新鮮土壤樣品,每年采集180個土壤樣品,3 a共計540個土壤樣品。取樣完成之后把樣品分為兩部分處理,一部分帶回實驗室進行風干操作,并對其雜質進行去除之后來科學地測定其養分和有效養分含量,另一部分則在取樣現場過100目篩后保存在4℃保溫冰箱之中,并對其土壤中各種微生物量進行測定[7-8]。
本研究過程中對土壤養分開展了一系列的測定,對有機碳、全氮及全鉀的測定分別采取氧化法、定氮法及光度計法,同時借助于烘干法來測定其含水量,要求溫度達到105℃,對于其pH值的測定則借助于電位法。
通過平板梯度稀釋法來測定土壤中各種微生物的數量。值得注意的是,高氏一號瓊脂培養基、馬丁氏培養基、牛肉膏蛋白胨瓊脂培養基這3種培養基分別為放線菌、真菌以及細菌的培養基,這樣能夠獲得多次重復的菌落平均數,用其乘以相應的稀釋倍數就得到了相應的微生物菌落數。對于微生物量碳、氮及磷的測定借助于浸提法進行。所有指標測定均重復3次后取平均值。
1.3 土壤微生物群落功能多樣性
微生物對于凋落物的分解發揮著無可替代的效果,同時在酶的參與下進一步提升了分解效果,二者是息息相關的,離開了微生物,凋落物等腐殖質的降解將無從談起,這在土壤養分轉化過程中尤為關鍵,直接制約著土壤肥力及活性,因此,在土壤質量分析過程中微生物常常作為關鍵指標加以分析。本研究過程中對于微生物多樣性的測定主要立足于McIntosh,Shannon和Simpson等指數的角度[19]。
在微生物呼吸的測定過程中,主要借助于堿液吸收法,其與微生物量碳之間的比值即為代謝熵:qCO2=MR/MBC[19]。本研究對于水解酶進行相應的測定,研究過程中借助于對硝基苯酚(pNP)法,對于微生物群落分布主要通過McIntosh,Simpson等多樣性指數進行衡量,對于Simpson指數計算而言,首先對相應的數據進行放大1 000倍的處理,這樣將不會出現負數的現象;對于微生物的碳源利用水平進行衡量過程中主要借助于AWCD值衡量法[20],即:
AWCD=∑[(Ci-R)/31]
(1)
H=-∑Pi(lnPi)
(2)
Pi=(Ci-Ri)/∑(Ci-Ri)
(3)
在開展指標統計分析之前對數據進行相應的均值處理,并充分考慮標準誤差,然后借助于Excel 2010整理相關數據,并開展SPSS 21.0統計檢驗,尤其是相關分析及顯著性檢驗等,并對不同的置信水平開展相應檢驗。在探究微生物和養分之間關系的過程中主要運用了CANOCO 4.5,此外還對其與環境因子之間的關系進行探討。
2 結果與分析
2.1 濱海濕地不同植物群落土壤養分
由圖1可知,濱海濕地不同植物群落土壤pH值變化范圍為7.13~7.56,基本表現為:互花米草濕地>堿蓬灘濕地>蘆葦濕地>紅樹林濕地,其中不同濕地植物群落土壤pH值差異均不顯著(p>0.05);土壤有機碳變化范圍為9.63~13.98 g/kg,基本表現為:互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中蘆葦濕地和紅樹林濕地差異不顯著(p>0.05),二者顯著高于互花米草濕地和堿蓬灘濕地(p<0.05);土壤全氮變化范圍為0.91~1.18 g/kg,基本表現為:互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地差異均顯著(p<0.05);土壤全氮變化范圍為26.52~43.09 g/kg,基本表現為:互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,紅樹林濕地顯著高于其他濕地(p<0.05)。
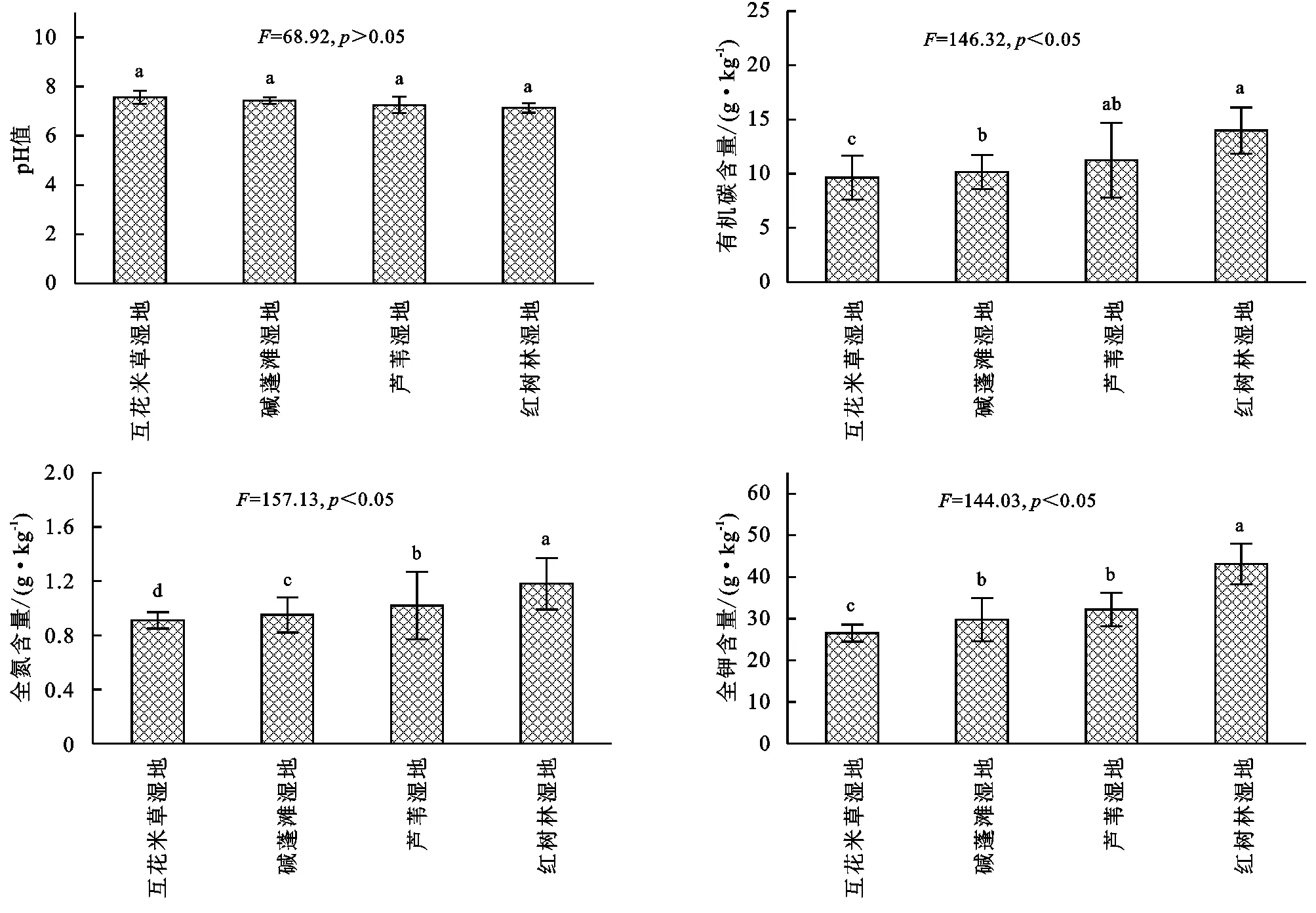
注:不同小寫字母表示差異顯著(p<0.05),下同。
2.2 濱海濕地不同植物群落土壤微生物數量
由圖2可知,濱海濕地不同植物群落土壤細菌數量占絕對優勢,占微生物總數的70%左右,其次是放線菌,真菌最少。土壤微生物總數變化范圍為(2.03~2.81)×107cfu/g,基本表現為:互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地土壤微生物總數差異均顯著(p<0.05);土壤細菌數目變化范圍為(1.23~1.57)×107cfu/g,基本表現為:互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中蘆葦濕地和紅樹林濕地差異不顯著(p>0.05),互花米草濕地顯著低于其他濕地(p<0.05);放線菌數目變化范圍為(3.1~6.1)×106cfu/g,基本表現為:互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地放線菌數目差異均顯著(p<0.05);真菌數目變化范圍為(4.5~8.4)×105cfu/g,基本表現為:互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地真菌數目差異均顯著(p<0.05)。綜合分析可知,微生物總數、細菌、真菌和放線菌數量最大值出現在紅樹林濕地,最小值出現在互花米草濕地。
2.3 濱海濕地不同植物群落土壤微生物量
由圖3可知,濱海濕地不同植物群落土壤微生物量存在一定差異,土壤微生物量碳變化范圍為124~169 mg/kg,基本表現為互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地差異均顯著(p<0.05);土壤微生物量氮變化范圍為68~92 mg/kg,基本表現為互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地差異均顯著(p<0.05);微生物量磷變化范圍為13~16 mg/kg,基本表現為互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地差異均不顯著(p>0.05)。
2.4 濱海濕地不同植物群落土壤微生物群落多樣性
由圖4可知,濱海濕地不同植物群落土壤微生物群落功能多樣性指數存在一定差異,土壤微生物物種豐富度指數(H)變化范圍為1.63~3.35,基本表現為互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地差異均顯著(p<0.05);土壤微生物均勻度指數(E)變化范圍為0.24~0.75,基本表現為互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中互花米草濕地和堿蓬灘濕地差異均不顯著(p>0.05);優勢度指數(Ds)變化范圍變化范圍為0.686~0.701,基本表現為互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地差異均不顯著(p>0.05);碳源利用豐富度指數(S)變化范圍為7.15~16.79,基本表現為互花米草濕地<堿蓬灘濕地<蘆葦濕地<紅樹林濕地,其中不同濕地差異均顯著(p<0.05)。
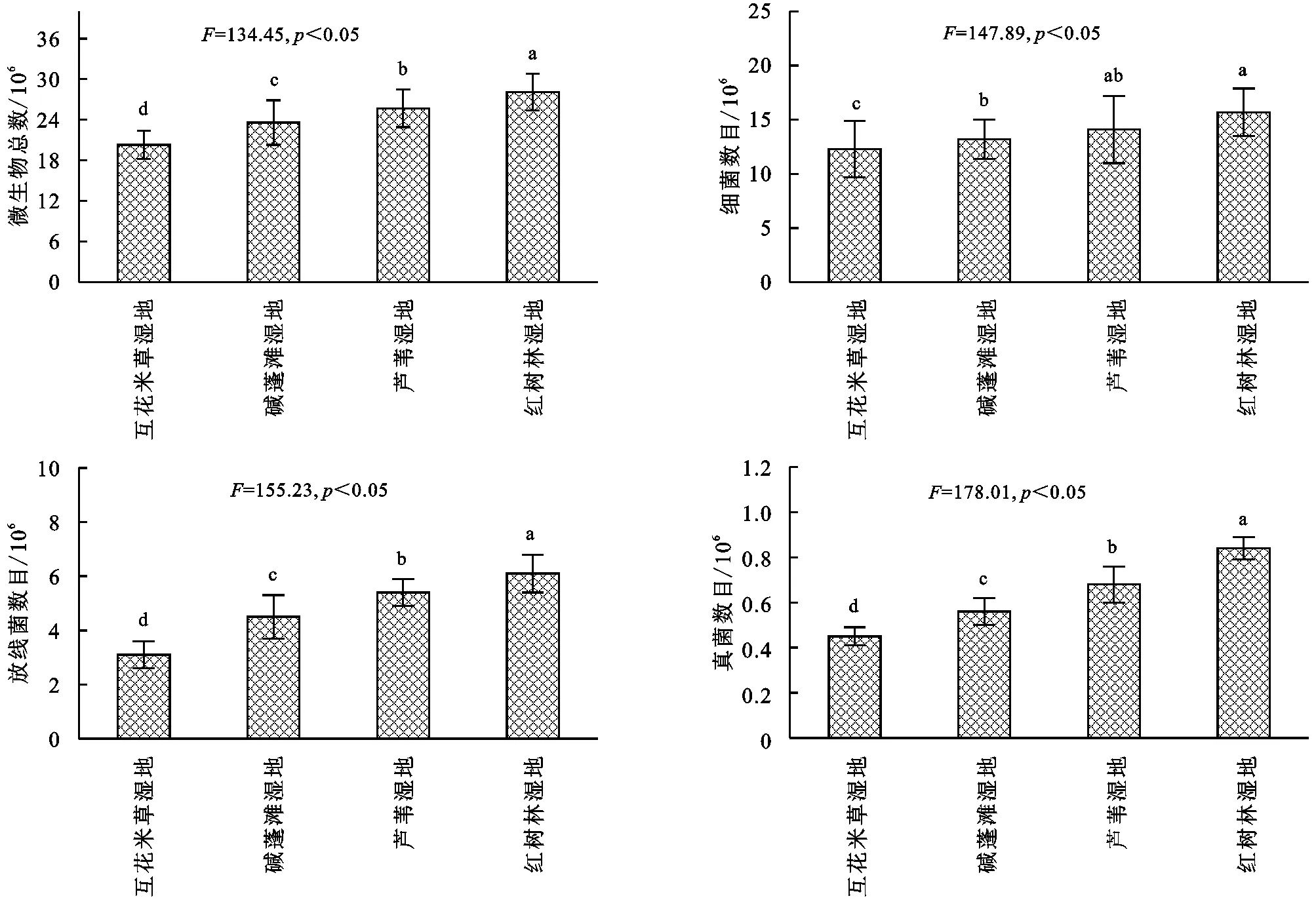
圖2 濱海濕地不同植物群落土壤微生物數量的變化
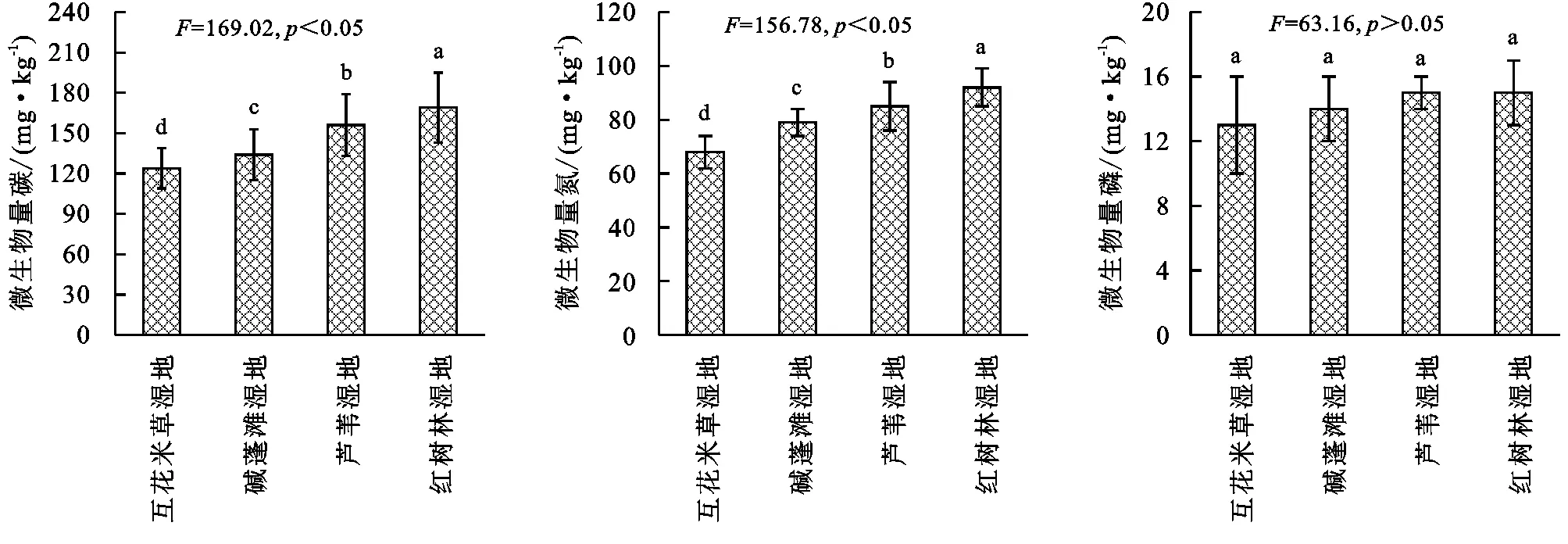
圖3 濱海濕地不同植物群落土壤微生物量的變化
2.5 土壤微生物及其與土壤養分之間的耦合關系
由表1可知,土壤微生物群落功能的變化與土壤養分的變化呈正比例關系。與土壤pH值之間的關系為負相關。土壤微生物碳源利用豐富度指數(S)、均勻度指數(E)和物種豐富度指數(H)土壤養分各指標的相關系數,均高于優勢度指數(Ds)。概而言之,土壤養分、土壤pH值和微生物群落功能多樣性之間,是相互依賴和影響的,土壤養分的貢獻程度較高,土壤pH值的貢獻程度較低。由以上推斷可知,濱海濕地不同植物群落土壤微生物群落多樣性會存在較大差異。
研究表明(表2),不同濕地4個特征值的累積方差貢獻率分別為70.23%,75.15%,72.27%,73.09%,基本能反映出大部分的變量信息,由此建立兩兩之間的4對典型變量的構成(表3),由于第1,2對典型變量相關系數大,第3,4對典型變量的影響較小,因此,只取第一、二對典型變量進行分析。
互花米草濕地,土壤微生物特征和土壤養分的第一、二對典型相關系數分別為0.992和0.856,表明土壤養分的第一、二對變量對土壤微生物第一、二對變量的影響較大(表3);堿蓬灘濕地,0.987**和0.814**(p<0.01),表明土壤養分的第一、二對變量對土壤微生物第一、二對變量的影響較大;蘆葦濕地,0.956**和0.809**(p<0.01),表明土壤養分的第一、二對變量對土壤微生物的第一、二對變量的影響較大;紅樹林濕地,0.986**和0.814**(p<0.01),表明土壤養分的第一、二對變量對土壤微生物第一、二對變量的貢獻較大。
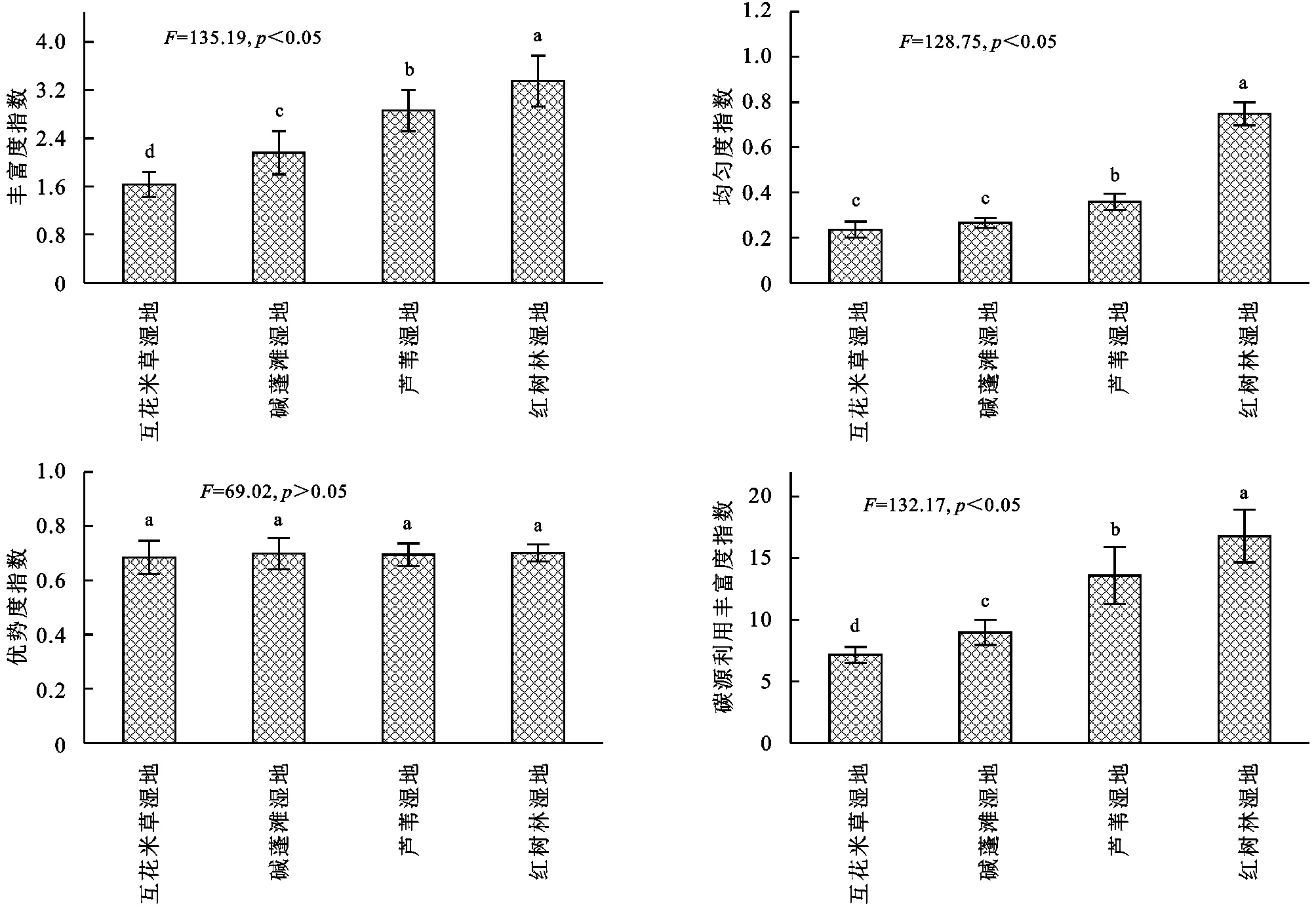
圖4 濱海濕地不同植物群落土壤微生物群落多樣性

表1 土壤養分與微生物多樣性之間相關性(n=540)
3 討 論
對于陸地生態系統,土壤微生物在物質和能量交換過程中起著重要的調節作用;土壤微生物也常常用于衡量土壤質量,一方面微生物活動改變著土壤理化特性,另一方面土壤環境為微生物活動創造更為適宜的新陳代謝環境,二者具有明顯的相互促進性[21]。
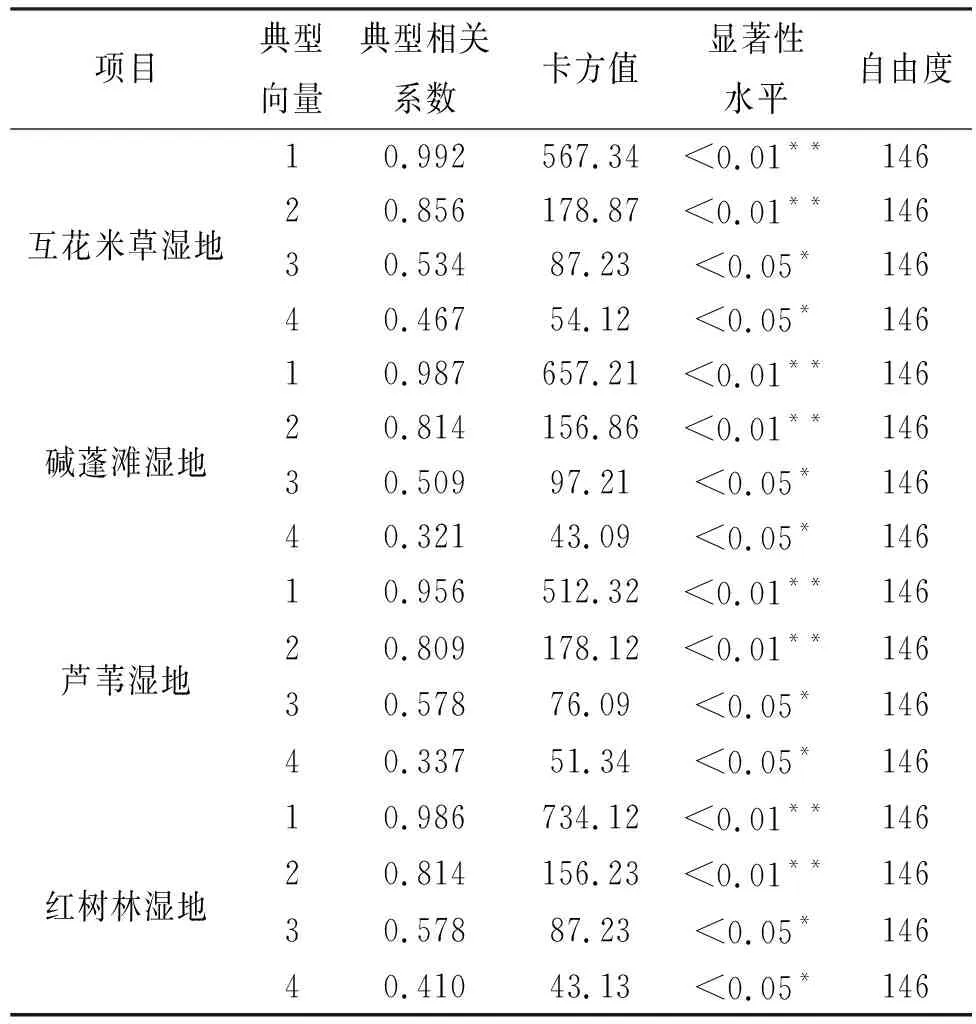
表2 土壤微生物與土壤養分典范相關分析的卡方檢驗

表3 土壤微生物與土壤養分的前兩對典型變量的構成
通過本試驗連續3年的觀測研究得知,雖然土壤的有機碳、全氮及全鉀存在較大的差異,但是其整體變化規律較為接近,其中分布含量最高的是紅樹林濕地,其次是堿蓬灘、蘆葦濕地,而最低的是互花米草濕地,對于微生物菌群而言,數量方面難以匹敵的依然是細菌,其占比高達70%,而真菌數量最不占據優勢;從其數量分布方面來講,其規律性較為明顯,數量最高的濕地依然是紅樹林,而數量最少的濕地依然是互花米草區域,對于不同濕地而言,其微生物數量呈現較大差異,且這種差異具有顯著性,達到了0.05的顯著性檢驗水平。對于紅樹林濕地區域而言,大量的枯枝落葉等凋落物形成了大量的腐殖質來源,在微生物作用下,其被分解為有機質,為土壤養分的積累奠定了基礎,同時,凋落物能夠有效促進土壤水分的保持,利于涵養降雨等水分,從而在促進林木生長的同時提升微生物新陳代謝的條件。此外,對于紅樹林區域而言,其具有更適宜微生物活動的表土結構,增強微生物對有機質的分解效率,加速了能量轉換。通過相關性分析得知,微生物數量及其生物量受到土壤養分的顯著制約,二者正向變化關系尤為突出,且其協調發展效應較強,具有較強的互補性[15]。微生物活動離不開必要的養分支撐,相反,凋落物等有機質的分解離不開微生物,從而促進養分積累,二者關系尤為密切,對于不同濕地而言,雖然其碳、氮含量存在較大差異,但是其含量的提升將會促進微生物量碳及氮的聚集。通過研究對比分析得知,對于微生物量而言,其與養分的關系密切,作為養分的核心來源之一,有機質含量起著關鍵作用;綜合來看,對于濱海濕地而言,雖然群落分布有所差異,但是微生物和養分之間的關系具有不可分割性,微生物的新陳代謝等活動利于養分的積累[22-23],對于有機質的降解效應尤為關鍵,二者具有較強的協同發展效應。
在土壤有機碳中,微生物量碳的含量一般不超過5%,對于全氮而言,微生物量氮的占比一般在6%左右,有些學者認為這并不是普遍現象,碳、氮含量的變化具有不確定性,也不是土壤的固化特點[22-23]。通過連續3年的觀測分析得知,對于微生物量碳、氮、磷而言,雖然在不同濕地區域來說,其含量存在較大差異,但是其依然具有一定的規律性,其中含量最高的是紅樹林濕地,其次蘆葦濕地區域,而互米花草濕地的含量最低,除了微生物量磷之外,碳、氮含量均呈現了顯著性差異。對于濱海濕地而言,微生物不同菌群在數量方面的差異較大,且其變化特點與總微生物數量基本接近,相比而言,細菌更具有數量優勢。不同的濕地區域,其不僅具有差異較大的土壤結構,其土壤肥力及活性方面也會出現較大差異,加之土壤通透性的差異,其將會對微生物活動產生顯著的制約,對于紅樹林區域而言,其不僅具有多樣性的植被分布,同時生物量突出,受凋落物影響,其水源涵養能力得以有效提升,土壤養分狀況相對較好,加之河水沖擊形成了養分集聚,對于提升該區域的養分水平效果顯著,受此影響,微生物活動進一步加強,對于有機質降解效果更好。雖然菌群的數量存在較大差異,但是不同的菌群對于環境的酸堿性喜好并不相同,對于真菌而言,其更適宜于在酸性條件下生長[24],而對于其他兩種菌群而言,則更喜好中性環境。
綜合來看,微生物活動改變著土壤理化特性,另一方面土壤環境為微生物活動創造更為適宜的新陳代謝環境,二者具有明顯的相互促進性,在濕地生態保護過程中要充分注重微生物和養分的協調,合理控制氮、磷等含量,發揮微生物效應。
猜你喜歡
歌海(2024年6期)2024-03-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
幼兒園(2021年4期)2021-07-28 07:38:04
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
歌海(2018年4期)2018-05-14 12:46:15
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
