Notch信號通路在骨髓間充質干細胞及骨代謝疾病中的功能研究進展
2020-10-26 06:13:24雷樂張玲莉楊杰
中國骨質疏松雜志 2020年9期
關鍵詞:信號
雷樂 張玲莉 楊杰*
1. 上海體育學院,上海 200438 2. 華南師范大學,廣東 廣州 510631
骨髓間充質干細胞(bone mesenchymal stem cells,BMSCs)是一種具有多向分化能力的成體干細胞,位于骨髓腔內。在一定的微環境或培養條件下,BMSCs可誘導分化為成骨細胞、軟骨細胞、脂肪細胞、成肌細胞、神經細胞和血管內皮細胞等[1-2]。而成骨細胞、骨細胞和破骨細胞的數量和協調活動決定了骨骼重塑和骨量的變化,因此BMSCs對骨骼發育和骨轉換起著至關重要的作用。近年來,研究發現Notch信號通路與BMSCs的增殖、分化與凋亡有密切相關。
Notch信號通路在細胞的整個生命活動中扮演著重要的角色,是一條保守而重要的信號轉導通路,在神經、心血管和骨骼等系統中都有重要的意義。Notch信號通路對骨骼細胞活動和骨骼發育尤為關鍵[3],調節細胞分化、增殖和凋亡,其功能異常可導致各種累及顱面與骨骼的先天性疾病[4]。且受多種因素的影響,譬如外源性藥物或因子調節Notch信號通路的關鍵基因和蛋白表達,機械載荷能夠激活Notch信號通路的靶基因,從而調節BMSCs的增殖與分化。
本文通過查閱國內外文獻,旨在闡述外源性藥物和因子和機械載荷介導Notch信號通路對BMSCs的影響,并總結了Notch信號通路在骨代謝疾病中的研究近況。
1 Notch信號通路與BMSCs的關系概述
Notch最早是在果蠅的研究中發現的一種基因,因該基因可以導致果蠅翅膀邊緣缺損,故被稱為Notch[5-6]。Notch信號通路由幾種高度保守的成員組成,主要包括受體、配體、CSL蛋白及Notch信號的效應分子[7]。在哺乳動物中Notch家族主要有4種受體(Notch1、Notch2、Notch3、Notch4)、5種配體(DLL1、DLL3、DLL4、Jag1、Jag2),均屬于單次跨膜蛋白。CSL蛋白是一種DNA結合蛋白,其名稱來源于哺乳動物(CBF1,也稱RBP-Jκ)、果蠅[Su(H)]與線蟲(Lag-1)的三個首字母,因為CSL蛋白主要存在于這三種物種之中。Notch信號效應分子主要為HES,是進化保守的基本螺旋-環-螺旋(bHLH)轉錄因子,HES家族中有6個成員,其中HES1、HES5、HES7在Notch信號通路中發揮重要作用,是Notch信號通路的下游靶基因[8]。Notch信號通路的激活主要經過三次酶切反應和蛋白水解:Notch受體合成后首先在高爾基體內被切割,為第一次酶切(S1);之后在胞外被腫瘤壞死因子(tumor necrosis factor-α,TNF-α)轉化酶酶切為第二次酶切(S2);酶切產生的不穩定過渡多肽被γ-分泌酶復合體識別并進一步將Notch受體的胞內段酶切,為第三次酶切(S3),三次酶切后Notch信號通路被激活,釋放出具有活性的Notch胞內結構域(Notch intracellular domain,NICD)進入細胞質,在胞質內移動進入細胞核,與核內的轉錄抑制因子CSL/CBF-1蛋白結合,將轉錄抑制因子轉換為轉錄活化因子,作用于Notch信號通路的靶基因HES(hairy/enhancer of split)、HEY(hairy/enhancer of split with YRPW motif family members),進而調節細胞的增殖、分化。
Notch信號通路在骨代謝中有重要作用,當其出現異常時會導致骨發育失調、骨質疏松、骨軟化等疾病。其上下游基因遺傳突變也會導致一系列骨疾病[9]。Notch信號通路對成骨細胞、破骨細胞的功能調節均有重要影響。DSL配體誘導Notch受體的切割和NICD的產生。成骨細胞前體中Notch1的活化可通過抑制osterix(Osx)、runt相關轉錄因子2(Runx2)和胞質β-連環蛋白抑制成骨細胞生成,影響骨形成;破骨細胞前體中的Notch2 NICD與NF-κB相關,誘導活化T細胞(Nfatc)1轉錄和破骨細胞增加,骨形成受損和骨吸收增加導致骨量減少(如圖1左側所示)。成骨細胞中Notch1的激活可誘導Rankl抑制劑骨保護素(OPG),從而抑制松質骨吸收。骨細胞中Notch1激活抑制硬化蛋白(Sclerostin)和Dkk1,增強Wnt信號,增加皮質骨形成。松質骨吸收減少和皮質骨形成增強導致骨量增加[10](如圖1右側所示)。Zanotti等[10]認為,Notch通過抑制Wnt信號傳導并與Runx2相互作用來抑制成骨細胞成熟,而在成熟的成骨細胞中Notch抑制其分化功能,Notch1可抑制破骨細胞生成,而Notch2則通過與NfκB相互作用來誘導Nfatc1與破骨細胞分化。同時,Deregowski等[11]研究發現,Notch1活化抑制成骨細胞分化,Notch能夠抑制Wnt/β-catenin信號傳導,但不抑制骨形態發生蛋白(bone morphogenetic protein,BMP)信號轉導。此外,Notch信號通路下游的HES和HEY蛋白能夠抑制Runx2的功能,從而抑制成骨細胞的生成[12]。Ji等[5]研究發現,Notch信號通路在BMSCs分化為成骨細胞時未被激活,在成骨細胞分化為骨細胞時被激活,而BMSCs成骨分化早期會抑制Notch信號傳導促進BMSCs向成骨細胞分化,積累成熟的成骨細胞,而到成骨細胞分化晚期,Notch信號傳導抑制成骨細胞向骨細胞的分化與礦化。Notch抑制MSC向成骨細胞的分化并促進成骨細胞向骨細胞的分化(如圖2所示)。也有研究[13]表明Notch通路對BMSCs的調節并不是單一的促進或抑制,而是在全身或局部相關因子調節下的結果。Notch信號通路對成骨細胞的作用更多是建立在與其他信號通路的相互作用,這些相互作用調節成骨細胞的機制尚未明確,有待進一步研究。
2 不同干預手段介導Notch信號通路對BMSCs的影響
2.1 外源性藥物或因子介導Notch信號通路對BMSCs的影響
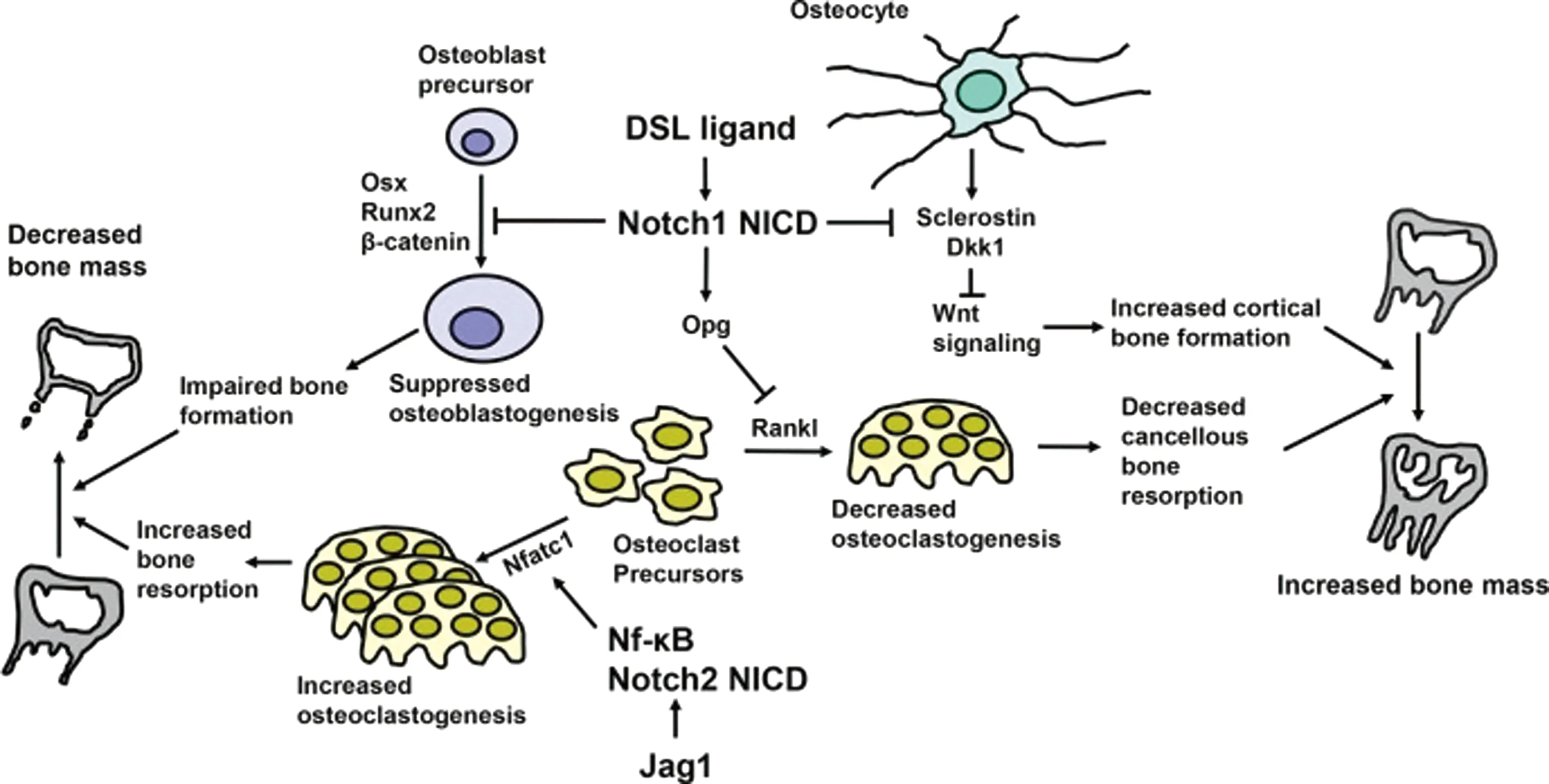
圖1 Notch信號傳導和骨重建調節[13]Fig.1 Notch signaling pathway and bone remodeling regulation
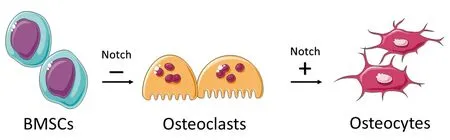
圖2 成骨細胞分化中Notch信號傳導模型Fig.2 Notch signaling model in the osteoblast differentiation
在國內,藥物干預對Notch信號通路的影響以中成藥居多。張穎等[14]研究發現,復方扶芳藤合劑含藥血清能上調大鼠BMSCs中Notch信號通路關鍵基因和蛋白的表達,激活Notch信號通路。韓亞力等[15]使用成分為骨碎補總黃酮(davallia mariesil flavones,DMF)的強骨膠囊進行動物實驗,發現DMF能下調大鼠Notch1與Hes1蛋白表達量,改善大鼠骨質疏松,提示其作用機制可能與Notch信號通路被抑制有關。周靈通[16]實驗發現固本增骨方含藥血清能上調Notch1 mRNA、下調CBF1 mRNA的表達,促進BMSCs增殖與成骨分化。張志明等[17]發現葛根素能使細胞DLL4含量增加,促進BMSCs骨向分化。鄧宇等[18]研究發現,淫羊藿苷能夠通過上調HES1、Runx2 mRNA的表達以及增加Notch1、CBF1、Jag1來促進BMSCs的骨向分化。Liu等[19]研究也發現淫羊藿苷可以抑制MSCs向成脂細胞分化,下調骨組織中Notch1細胞內結構域(N1ICD)Jag1蛋白的表達,并抑制N1ICD對Notch2 mRNA表達,改善去卵巢大鼠骨質疏松。邢貞武[20]發現提取骨質疏松患者的BMSCs進行離體培養,添加補骨脂素后,受體Notch1、配體Jag1和Hes1的表達增高,說明補骨脂素能提高BMSCs的Notch信號通路的關鍵分子表達,提示補骨脂素能通過Notch信號通路促進成骨分化,抑制成脂分化。
目前,對作用于Notch信號通路的西藥研究較少,介導Notch信號通路調節BMSCs分化功能的西藥研究尚未報道,而中藥調節Notch信號通路僅僅是對通路上重要靶基因和關鍵蛋白進行研究,其具體機制尚不明確,還有待于進一步研究。
2.2 外源性因子介導Notch信號通路對BMSCs的影響
外源性因子可以介導Notch信號通路誘導BMSCs分化為成骨細胞、軟骨細胞等,對骨代謝維持平衡具有重要意義。Diaz-Tocados等[21]通過實驗觀察發現,氯化鎂能夠通過Notch信號傳導激活BMSCs擴增,促進成骨細胞成熟。王維東等[22]研究發現甲狀旁腺素(parathyroid hormone,PTH)基因敲除的小鼠其Notch信號通路上的受體Notch1和配體Jag1表達低于野生小鼠,提示內源性PTH缺失可能通過降低Notch信號通路受體和配體的表達來抑制BMSCs的骨向分化。范金柱等[23]發現絕經后骨質疏松女性與健康女性相比,BMSCs中Notch信號通路減弱,給予患者補充雌激素后,Notch信號通路恢復活性,受體Notch1、配體Jag1表達升高,通路下游分子Hes1也表達上升,提示雌激素能促進BMSCs向成骨分化,Notch信號通路可能成為治療絕經后骨質疏松患者的新靶點。徐瑩等[24]對培養的BMSCs給予糖皮質激素(glucocorticoids, GCs)處理后發現Notch信號通路的靶基因Hes1表達降低,提示GCs可以通過抑制Notch信號通路表達抑制BMSCs成骨分化。鄭介柏等[25]發現γ分泌酶特異性抑制劑DAPT(γ-secretase inhibitor)能夠阻斷Notch信號通路活化,抑制BMSCs的增殖,同時Notch信號通路活化抑制后,ALP活性增強,BMSCs骨向分化能力增強。
外源性的因子通過調節Notch信號通路的關鍵受體和配體,影響BMSCs的分化功能,涉及到激素、金屬離子,但是研究較少,其具體機制仍有待進一步闡述。
2.3 機械刺激介導Notch信號通路對BMSCs的影響
機械刺激調節Notch信號傳導并影響細胞的活動。Manokawinchoke等[26]對小鼠前成骨細胞系干預發現,間歇性壓應力上調Notch信號傳導的靶基因表達,誘導的Hes1、Hey1 mRNA可被DAPT抑制,這一實驗提示Notch信號參與了骨細胞在機械應激刺激下的穩態維持。張鵬等[27]使用基因芯片技術檢測到Notch信號通路中的Jag1在應力組明顯上調,可能是Notch信號通路被早期激活,參與BMSC成骨分化過程。Tanabe等[28]研究發現Notch1蛋白在壓力刺激下降低,而骨膜蛋白(Periostin)能夠與Notch1前體結合,維持Notch1在壓力刺激下的信號傳導。劉玉林等[29]研究發現,BMSCs在低聲壓次聲的干預下Jag1、Notch1、Hes1 基因及蛋白表達顯著增加,激活Notch信號通路。Ziouti等[30]研究發現,循環拉伸BMSCs后通過生物反應器系統檢測到Notch靶基因的表達上調,進一步研究證明Notch信號控制BMSCs中機械反應基因的表達,其中Notch2是介導Notch信號對BMSCs作用的關鍵受體。在機械刺激下,BMSCs向成骨分化的生物特性會發生相應改變,Notch信號傳導中的受體和配體也會受到影響。
3 Notch信號通路在骨代謝相關疾病中的功能作用
3.1 Notch信號通路與Alagille綜合征
Alagille綜合征(Alagille syndrome,ALGS)是一種多系統受損的遺傳疾病, 33%~87%的患者出現骨骼異常,表現為椎骨異常(蝴蝶椎骨)或矢狀裂,受影響的椎骨椎體分裂,形成成對的半椎骨,在影像學中表現為“蝴蝶”特征的外觀[31]。同時患者還可能出現身材矮小、骨質疏松等表現[32]。Notch信號通路中的Jag1和Notch2基因突變可能是造成Alagille綜合征的原因之一。在94%~96%的病例中可檢測到Jag1基因出現變異,1%~2%的病例中檢測到Notch2基因突變[33-34]。
3.2 Notch信號通路與Adams Oliver綜合征
Adams Oliver綜合征(Adams Oliver syndrome,AOS)是一種罕見的先天性疾病,其特征是先天性發育不全和末端橫向肢體缺陷,該疾病與6種遺傳基因突變相關(ARH-GAP31、DLL4、Notch1、RBPJ、DOCK6、EOGT)[35],Notch1、DLL4和RBPJ基因編碼的蛋白質是Notch信號轉導途徑的一部分。其中,Notch1和DLL4蛋白對血管發育至關重要,Notch1突變主要與肢體缺陷有關,表現為肢體指體缺失或畸形,患者手指或腳趾可能表現為融合、過短或缺失[36]。
3.3 Notch信號通路與脊椎肋骨發育不良
脊椎肋骨發育不良是一種罕見疾病,其特征是椎體分割缺陷和繼發缺陷性椎體形成的肋骨異常,表現為頸部短而僵硬和軀干較短[37-38]。Notch信號通路的功能之一是在早期發育過程中調節椎骨和肋骨分開,即體節分割。當Notch信號轉導途徑被破壞時,體節分割不能正常發生,導致椎骨、肋骨的畸形和融合[39]。這種疾病產生可能是DLL3突變造成Notch配體截斷或折疊錯誤的蛋白翻譯[40]。
3.4 Notch信號通路與Hajdu-Cheney綜合征
Hajdu-Cheney綜合征(Hajdu-Cheney syndrome,HCS)是一種罕見的遺傳性疾病,也稱為遺傳性骨發育不良并肢端溶骨癥,其特征為手和腳的骨質疏松癥以及骨骼、牙齒和關節的發育缺陷,導致特殊的顱面和顱骨改變,骨質疏松癥和身材矮小。Notch 2的外顯子區域缺失或發生無義突變,導致含脯氨酸-谷氨酸-絲氨酸-蘇氨酸的結構域缺乏蛋白質產物[41]。突變導致截短的穩定Notch2蛋白的形成和Notch2在信號傳導中增強。顱面和心血管發育異常的病理機制可能與Notch對骨骼和心臟發育的影響有關,身材矮小可能繼發于Notch對軟骨形成的抑制作用,但具體的發病機制尚未闡明。
3.5 Notch信號通路與骨折愈合
骨折愈合是一個復雜的再生過程,骨折后會產生結締組織和新骨。當骨折機械穩定時,骨折愈合通過膜內骨形成,當骨不穩定時由軟骨內骨化[42]。在骨折愈合早期的骨膜中,Notch受體Notch1-4、配體Jag2和靶基因HES1、HEY1的表達受到抑制[43],又有研究發現經過5、10、20 d愈合的小鼠脛骨組織中,Notch受體、配體和靶基因上調,主要是由于Notch信號傳導可以調節成骨細胞、軟骨細胞與骨血管生成,在骨生成過程中表達更高以增強骨修復[44]。由于Notch對骨生成和軟骨形成具有抑制作用,Notch信號的下調可能是骨折愈合過程發生的必要條件,使用在MSH同源框1啟動子控制下表達的顯性陰性MAML1來抑制Notch轉錄激活的研究表明,Notch信號傳導是成功骨折愈合所必需的[45]。
綜上所述,Notch信號通路對BMSCs的命運有重要的影響,調節成骨細胞的分化和功能。在未成熟的成骨細胞中,Notch可以抑制成骨細胞成熟及分化;在成熟的成骨細胞中,Notch抑制其分化。這也使Notch信號通路近年來成為骨重建研究中的熱點。
Notch信號通路與先天性骨代謝疾病密切相關,也說明其在骨骼發育中發揮重要功能。通過梳理目前BMSCs中Notch信號通路的相關研究,發現外源性藥物或性因子與機械載荷可以介導Notch信號通路調節BMSCs的多種分化功能,從而調節骨骼發育。Notch信號通路異常時會影響骨骼發育,這成為多種骨代謝疾病可能的病理機制之一。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
