冰封期內(nèi)蒙古南海湖細(xì)菌菌群結(jié)構(gòu)特征分析
2020-10-28 03:12:40楊文煥毋凡琛李衛(wèi)平
灌溉排水學(xué)報(bào) 2020年10期
楊文煥,毋凡琛,申 涵,李衛(wèi)平,張 生
(1.內(nèi)蒙古農(nóng)業(yè)大學(xué) 水利與土木建筑工程學(xué)院,呼和浩特 010018; 2.內(nèi)蒙古科技大學(xué) 能源與環(huán)境學(xué)院,內(nèi)蒙古 包頭 014010)
0 引 言
細(xì)菌群落作為湖泊生態(tài)系統(tǒng)中重要組成部分,在推動(dòng)湖區(qū)生物地球化學(xué)循環(huán)方面發(fā)揮著重要作用[1]。細(xì)菌不僅可通過(guò)自身新陳代謝與環(huán)境發(fā)生作用,進(jìn)而改變水環(huán)境因子,亦可作為生物學(xué)指標(biāo),來(lái)指示湖泊生態(tài)系統(tǒng)的健康狀況[2]。【研究意義】寒旱區(qū)湖泊由于其特殊的地理位置,和氣候特征,導(dǎo)致其有冰封期長(zhǎng)、冰層厚、流量小、污染重等特有的水體特征以及其重要的生態(tài)系統(tǒng)功能,逐漸成為國(guó)內(nèi)外研究的焦點(diǎn)[3-4]。在冰封期間,由于湖泊水溫急劇下降,大量生物在極端環(huán)境下存活困難,但依然有些細(xì)菌還具有強(qiáng)大的生物活性,特殊的遺傳背景和代謝途徑使其能夠產(chǎn)生適合低溫極端環(huán)境的特殊酶類及活性物,從而使其能夠在寒冷條件下表現(xiàn)出較高的活性[5-6]。【研究進(jìn)展】目前對(duì)于寒旱區(qū)湖泊的研究主要集中在其功能、水質(zhì)、魚(yú)類群落、鳥(niǎo)類、浮游生物和水生大型植物方面,而針對(duì)南海湖的研究多集中在營(yíng)養(yǎng)鹽分布、生態(tài)風(fēng)險(xiǎn)評(píng)價(jià)和浮游植物分布等,而對(duì)微生物的研究還較少。【切入點(diǎn)】由于寒旱區(qū)湖泊具有較長(zhǎng)的冰封期這一特點(diǎn),而在冰封過(guò)程中,湖泊的生態(tài)系統(tǒng)特征受到物理和化學(xué)波動(dòng)的影響,由此產(chǎn)生的一系列生態(tài)過(guò)程及表象將對(duì)生存在該生態(tài)系統(tǒng)中的細(xì)菌多樣性和代謝活動(dòng)產(chǎn)生更大的影響,進(jìn)而導(dǎo)致整個(gè)食物鏈的結(jié)構(gòu)發(fā)生變化[8]。因此,通過(guò)探究寒旱區(qū)湖泊冰封期細(xì)菌種群特征,了解細(xì)菌群落的組成和多樣性,對(duì)分析湖泊生態(tài)系統(tǒng)物質(zhì)循環(huán),以期能錨定該湖泊中主要優(yōu)勢(shì)菌屬,進(jìn)而解決寒旱區(qū)水環(huán)境問(wèn)題具有指導(dǎo)意義。【擬解決的關(guān)鍵問(wèn)題】本研究采用高通量測(cè)序技術(shù),通過(guò)對(duì)寒旱區(qū)湖泊在冰封期不同區(qū)域水體及沉積物細(xì)菌群落特征進(jìn)行分析,與環(huán)境因子進(jìn)行相關(guān)性分析,來(lái)探究寒旱區(qū)冰封期湖泊理化因子對(duì)細(xì)菌群落特征的影響,為進(jìn)一步探究高原寒旱區(qū)湖泊細(xì)菌群落與環(huán)境的相應(yīng)關(guān)系和富營(yíng)養(yǎng)化水體的生物修復(fù)提供參考。
1 材料與方法
1.1 采樣點(diǎn)布設(shè)
南海湖受人為因素影響顯著,不同水域有不同特征。根據(jù)湖泊補(bǔ)水特征、水生植物生長(zhǎng)特點(diǎn)及人為因素干擾等,將其劃分為4 個(gè)區(qū)域,分別為進(jìn)水口區(qū)S1(NH1、NH2、NH3),旅游開(kāi)發(fā)區(qū)S2(NH4、NH5、NH6),湖心區(qū)S3(NH7、NH8、NH9)及水草區(qū)S4(NH10、NH11、NH12),在每個(gè)區(qū)域均布設(shè)3 個(gè)采樣點(diǎn)。湖泊西南部定期接受黃河補(bǔ)水,水質(zhì)影響明顯;補(bǔ)水口東北部坐落一座小島,供游人觀光,污染較多;島嶼周?chē)昂礀|北部水草密集生長(zhǎng),以蘆葦為主;湖中心污染較少,水力條件穩(wěn)定。采樣點(diǎn)具體分布如圖1 所示。
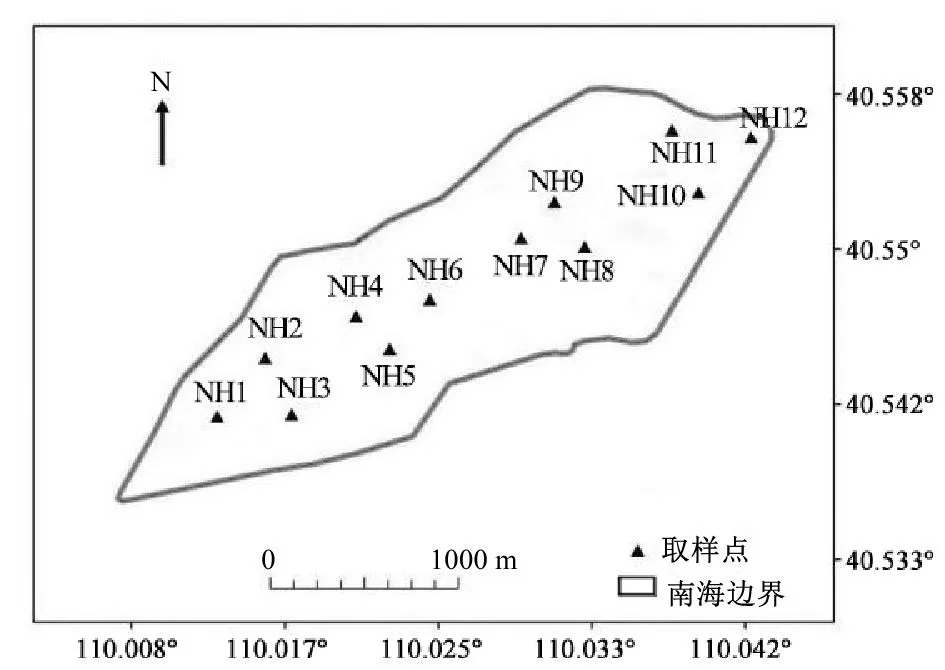
圖1 包頭南海湖冰封期采樣點(diǎn)布設(shè)圖 Fig.1 Distribution of sampling sites in the ice period of Nanhai Lake in Baotou
1.2 樣品采集與處理
于2017 年12 月—2018 年2 月在研究區(qū)域各采樣點(diǎn)采集樣品。采集水樣時(shí),使用不銹鋼柱狀采樣器,取冰體下表面之下0.5 m 與沉積物之上0.5 m 處水樣進(jìn)行混合,水樣存放于經(jīng)滅菌消毒的聚乙烯取樣瓶中。于實(shí)驗(yàn)室內(nèi)將樣品分為兩部分,一部分用于理化指標(biāo)的測(cè)定,另一部分經(jīng)0.22 μm 無(wú)菌微濾孔濾膜過(guò)濾,將濾膜置于50 mL 無(wú)菌離心管中,放置于-80 ℃液氮中保存,用于微生物群落結(jié)構(gòu)和多樣性分析。
使用不銹鋼采樣器取表層沉積物樣品,每個(gè)樣點(diǎn)均取3 個(gè)平行樣,充分混合后存放于經(jīng)滅菌消毒聚乙烯自封袋中(保證無(wú)菌)。將樣品分為2 部分,一部分待自然風(fēng)干后研磨用于各理化指標(biāo)測(cè)定,另一部分置于-80 ℃液氮冷凍保存,用于微生物群落結(jié)構(gòu)和多樣性分析。
1.3 理化因子檢測(cè)
本研究水質(zhì)檢測(cè)指標(biāo)為水溫(T)、溶解氧(DO)、pH 值、總氮(TN)、總磷(TP)、氨態(tài)氮(NH4+-N)、硝態(tài)氮(NO3--N)、亞硝態(tài)氮(NO2--N)、葉綠素a(Chl.a)和化學(xué)需氧量(CODCr)。其中水溫,溶解氧和pH 值采用便攜式水質(zhì)檢測(cè)儀在采樣現(xiàn)場(chǎng)檢測(cè),其他指標(biāo)根據(jù)國(guó)家環(huán)境保護(hù)總局《水和廢水監(jiān)測(cè)分析方法》中所述方法檢測(cè)。沉積物理化指標(biāo)包括pH 值、總有機(jī)碳(TOC)、總磷(TP)、總氮(TN)、氨態(tài)氮(NH4+-N)、硝態(tài)氮(NO3-N)和碳氮比(C/N)。具體測(cè)定方法如表1、表2 所示。

表1 水體理化指標(biāo)測(cè)定方法 Table 1 Test method of physical and chemical indexes of water
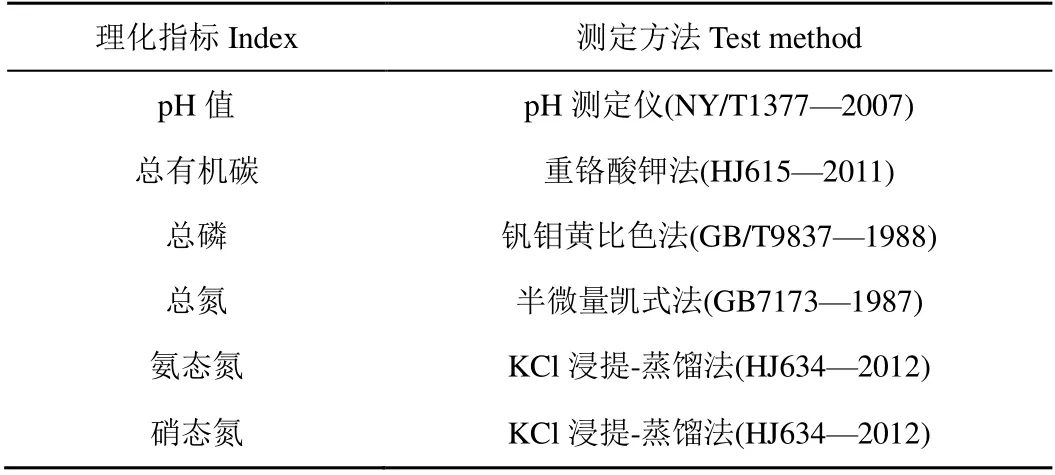
表2 沉積物理化指標(biāo)測(cè)定方法 Table 2 Test method of physical and chemical indexes of sediments
1.4 高通量測(cè)序
采用 OMEGA 的 Soil DNA Isolation Kit(BioTek,Inc.,GA,USA)提取沉積物及水體樣品總DNA,完成基因組DNA 抽取后,使用1%瓊脂糖凝膠電泳檢測(cè)抽取基因組DNA。采用通用引物338F(5'-ACTCCTACGGGAGGCAGCA-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3')作為 16S rRNA 基因序列PCR 擴(kuò)增的引物序列,進(jìn)行V3~V4區(qū)目的基因PCR 擴(kuò)增。將PCR 產(chǎn)物用2%瓊脂糖凝膠電泳檢測(cè),使用 AxyPrepDNA 凝膠回收試劑盒(AXYGEN)公司切膠回收PCR 產(chǎn)物,Tris-HCl 洗脫,用25%瓊脂糖電泳檢測(cè),參照電泳初步定量結(jié)果,將PCR 產(chǎn)物用QuantiFluorTM-ST 藍(lán)色熒光定量系統(tǒng)進(jìn)行檢測(cè)定量,隨后按照每個(gè)樣本測(cè)序量要求,按照相應(yīng)比例混合,做Miseq 測(cè)序[10],高通量測(cè)試在上海美吉生物公司進(jìn)行。
1.5 數(shù)據(jù)分析
將相似性大于97%的優(yōu)化序列劃分為一個(gè)操作分類單元(Operational Taxonomic Units,OTU),對(duì)OTU 樣本進(jìn)行聚類分析,計(jì)算Chao1 指數(shù)、香農(nóng)指數(shù)(Shannon)、文庫(kù)覆蓋率(Coverage),采用RDP-classifier 貝葉斯算法從各個(gè)分類水平對(duì)各個(gè)樣本的群落組成進(jìn)行分析[11]。利用相關(guān)性Spearman圖分析理化因子與細(xì)菌群落結(jié)構(gòu)關(guān)系;Origin(8.5版本)繪制物種豐度柱狀圖。采用SPSS 進(jìn)行方差分析。
2 結(jié)果與分析
2.1 理化指標(biāo)
南海湖水環(huán)境及沉積物理化因子分布情況見(jiàn)表3、表4。從表3 可以看出,不同湖區(qū)營(yíng)養(yǎng)水平有一定差異,從水體看(表3),TN、NH4+-N、NO3--N、T 差異顯著(0.01
島內(nèi)各種生活生產(chǎn)廢水排放入旅游開(kāi)發(fā)區(qū),由于受人為污染較多,這一區(qū)域營(yíng)養(yǎng)水平較其他區(qū)域偏高;由于水草區(qū)水生植物在冰封期的腐敗,殘?bào)w分解等過(guò)程,體內(nèi)營(yíng)養(yǎng)元素釋放,各污染物的量處于較高的水平,且TP 和COD 較其他區(qū)域高;進(jìn)水口受到黃河補(bǔ)水的影響,稀釋了水體中營(yíng)養(yǎng)物質(zhì)造成,營(yíng)養(yǎng)水平較低;湖心區(qū)受人為干擾因素較小,污染較少。總體來(lái)看沉積物營(yíng)養(yǎng)水平與冰封期冰下水體分布大致相同。
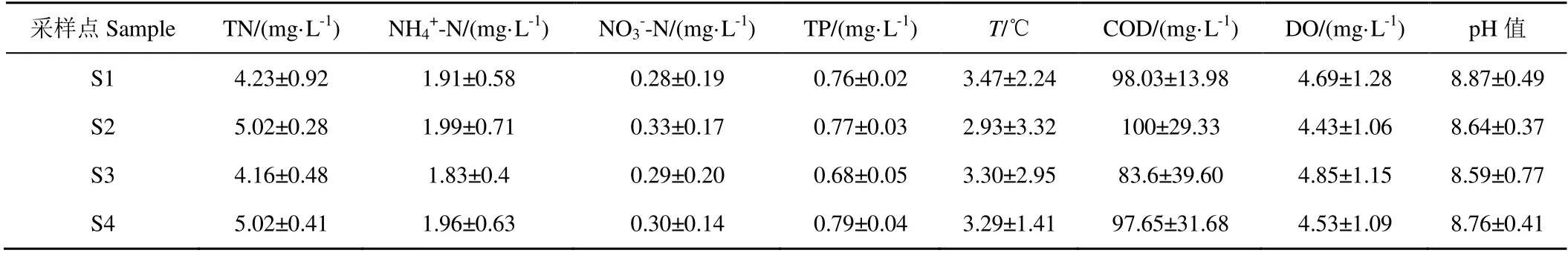
表3 南海湖水環(huán)境因子分布特征 Table 3 Distribution characteristics of water environmental factors in Nanhai Lake

表4 南海湖沉積物理化因子分布特征 Table 4 Distribution characteristics of sediments factors in Nanhai Lake
不同湖區(qū)沉積物中污染物質(zhì)的量有一定差異,污染嚴(yán)重程度由高到低依次為S2>S4> S1> S3。沉積物整體呈弱堿性,TN、NH4+-N差異極顯著(p<0.01),NO3--N 在S1、S3 湖區(qū)差異極顯著(p<0.01),在S2、S4 湖區(qū)(0.01
2.2 OTU 聚類及多樣性分析
各采樣點(diǎn)α多樣性指數(shù)及OTU聚類結(jié)果見(jiàn)表5。測(cè)序結(jié)果顯示覆蓋率均在99%以上,表明測(cè)序結(jié)果基本代表了樣本中微生物的真實(shí)情況[12]。
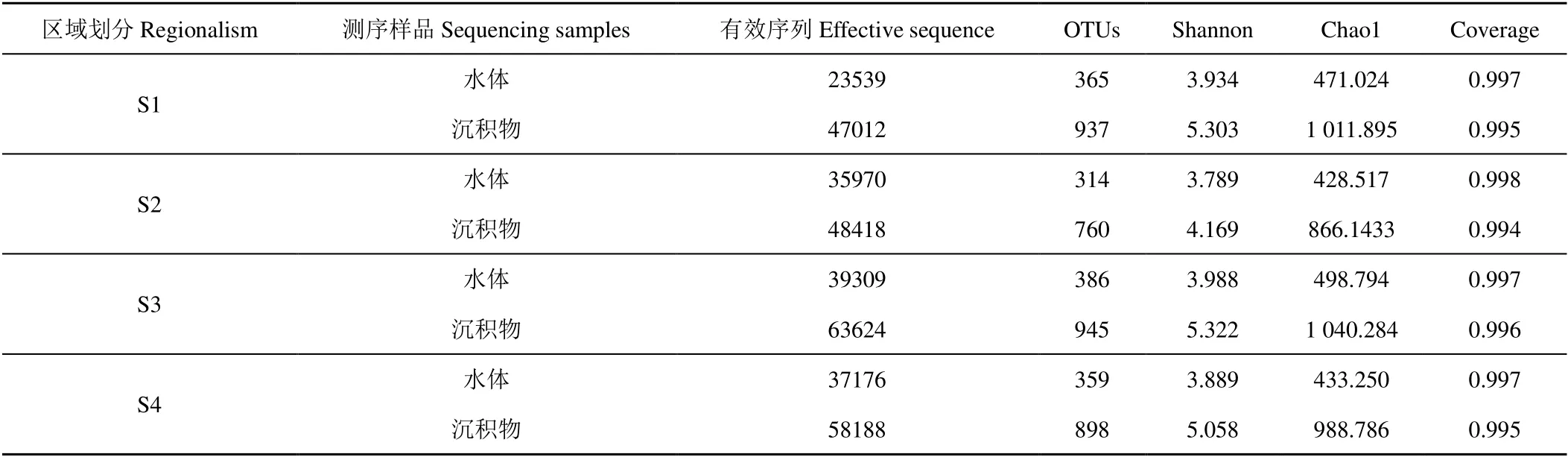
表5 各區(qū)域水體及沉積物OTU 聚類與多樣性分析 Table 5 OTU clustering and diversity analysis of water and sediment in different regions
以多樣性指數(shù)Sobs 做稀釋曲線(圖2),各樣點(diǎn)的稀釋曲線逐漸趨緩。通過(guò)Chao1、Shannon 等多樣性指數(shù)綜合來(lái)看,在本次試驗(yàn)中不同湖區(qū)樣品物種豐度和均勻度大小依次為S3>S1>S4>S2。在冬季冰封期由于冰蓋的影響導(dǎo)致水體內(nèi)溶解氧較低,因此溶解氧成為制約微生物量的關(guān)鍵因素。
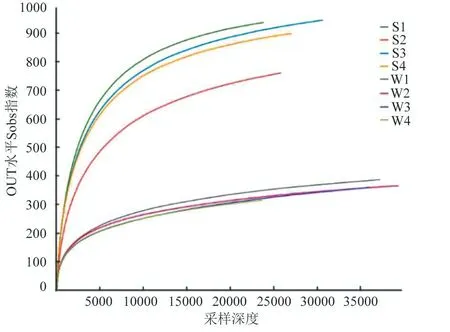
圖2 稀釋曲線 Fig.2 Rarefaction curve
為了探究不同區(qū)域水體、沉積物細(xì)菌的群落組成差異,對(duì)各個(gè)區(qū)域樣品進(jìn)行β 多樣性分析(圖3),在沉積物中湖心區(qū)與水草區(qū)沉積物細(xì)菌組成最為接近,進(jìn)水口區(qū)次之;進(jìn)水口區(qū)與旅游開(kāi)發(fā)區(qū)細(xì)菌組成差異最大。在水體中,進(jìn)水口區(qū)與湖心區(qū)細(xì)菌群落最為相似,水草區(qū)次之,類似的營(yíng)養(yǎng)條件使得細(xì)菌群落結(jié)構(gòu)最為接近。旅游開(kāi)發(fā)區(qū)在其他季節(jié)受人為影響較大,故在經(jīng)歷冰封期的沉降作用后與其他區(qū)域細(xì)菌群落組成差異較大;雖然在冰封期進(jìn)水口區(qū)接受黃河補(bǔ)水有限,但也對(duì)沉積物細(xì)菌組成造成了一定影響;湖心區(qū)與水草區(qū)均位于內(nèi)湖,受外界干擾較小,細(xì)菌組成也最為接近。
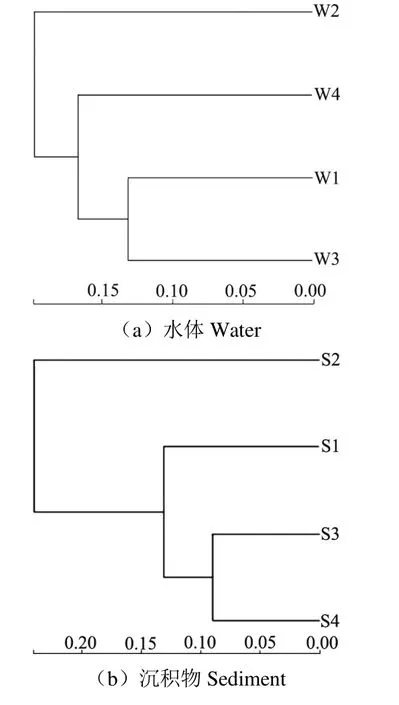
圖3 樣本層級(jí)聚類樹(shù) Fig.3 Hierarchical clustering tree
2.3 細(xì)菌群落結(jié)構(gòu)
對(duì)測(cè)序結(jié)果進(jìn)行分析得出,南海湖在冰封期水體及沉積物中具有較豐富的微生物群落結(jié)構(gòu)。共計(jì)46 個(gè)門(mén),1 073 個(gè)屬。按門(mén)水平分類,水體及沉積物中優(yōu)勢(shì)種均集中在變形菌門(mén)(Proteobacteria)、綠彎菌門(mén)(Chloroflexi)、藍(lán)藻門(mén)(Cyanobacteria)、擬桿菌門(mén)(Bacteroidetes)和放線菌門(mén)(Actinobacteria)等(圖4),其中變形菌門(mén)和綠彎菌門(mén)是沉積物中的優(yōu)勢(shì)菌屬,綠彎菌門(mén)和擬桿菌門(mén)是水體中的優(yōu)勢(shì)菌。進(jìn)水口區(qū)沉積物中變形菌門(mén)和綠彎菌門(mén)為優(yōu)勢(shì)菌種,此外,藍(lán)藻細(xì)菌也較為豐富,水體中放線菌門(mén)、藍(lán)藻門(mén)為優(yōu)勢(shì)菌種。旅游開(kāi)發(fā)區(qū)沉積物中主要菌種為變形菌門(mén)、綠彎菌門(mén)和擬桿菌門(mén),水體中主要菌種為藍(lán)藻門(mén)、變形菌門(mén)和放線菌門(mén)。湖心區(qū)與水草區(qū)沉積物細(xì)菌組成較為相似,主要菌種為變形菌門(mén)、綠彎菌門(mén),但二者的水體細(xì)菌差異較大,湖心區(qū)的優(yōu)勢(shì)菌種為放線菌門(mén),水草區(qū)的優(yōu)勢(shì)菌種為藍(lán)藻門(mén)。

圖4 樣品門(mén)水平優(yōu)勢(shì)細(xì)菌分布 Fig.4 Distribution of dominant bacteria at the phylum level

圖5 樣品屬水平優(yōu)勢(shì)細(xì)菌分布 Fig.5 Distribution of dominant bacteria at the genus level
對(duì)樣品在屬水平上進(jìn)行相對(duì)豐度計(jì)算(圖5), 在沉積物中硫桿菌屬(Thiobacillus)、未命名厭氧繩菌科(norank-f-Anaerolineaceae)、未分類綠彎菌科(unclassified-p-Chloroflexi)是優(yōu)勢(shì)菌屬,在水體中,未分類藍(lán)藻細(xì)菌(unclassified-Cyanobacteria)、分支桿菌屬(Mycobacterium)和hgcl-clade 是優(yōu)勢(shì)菌屬。硫桿菌屬和norank-f-Anaerolineaceae 在進(jìn)水口區(qū)豐度接近;硫桿菌屬在旅游開(kāi)發(fā)區(qū)內(nèi)豐度較其他區(qū)域更高,此外unclassified-p-Chloroflexi 豐度也處于4 個(gè)采樣點(diǎn)的最高水平;在湖心區(qū)與水草區(qū),豐度最高的為norank-f-Anaerolineaceae,硫桿菌屬的稍低,除此之外螺旋體屬(Spirochaeta-2)、暖發(fā)菌屬(Caldithrix)、硝化螺菌屬(Nitrospira)等也均有檢出。
2.4 細(xì)菌群落與環(huán)境因子相關(guān)性
環(huán)境因子對(duì)細(xì)菌群落結(jié)構(gòu)有著顯著影響,通過(guò)對(duì)環(huán)境因子與主要屬水平細(xì)菌的相關(guān)性熱圖得出(圖6):unclassified-c-Cyanobacteria 與pH 值、COD、DO、T 正相關(guān),與TP、TN 負(fù)相關(guān);Mycobacterium與TP、COD 正相關(guān),與NH4+-N、NO3--N、NO2--N呈負(fù)相關(guān);norank-o-PeM15 則與NH4+-N、NO3--N、NO2--N、Chaol 呈極顯著負(fù)相關(guān)(p<0.001)。對(duì)hgcl-clade 影響較大的環(huán)境因子是TN。總體來(lái)看,TN、DO、T 對(duì)水體細(xì)菌群落結(jié)構(gòu)影響較大。

圖6 Spearman 相關(guān)性熱圖 Fig.6 Spearman correlation heat map
沉積物中硫桿菌屬、unclassified-p-Chloroflexi 與TN、NH4+-N 正相關(guān),與C/N 負(fù)相關(guān),與TP 相關(guān)性不大;norank-f-Anaerolineaceae、norank-c-KD4-96與 TOC 負(fù) 相 關(guān), 與 pH 值 正 相 關(guān); 對(duì)norank-c-Cyanobacteria 影響較大的環(huán)境因子是TOC。總體來(lái)看,TN、NH4+-N、C/N 對(duì)沉積物細(xì)菌群落結(jié)構(gòu)影響較大。沉積物理化因子的量受湖泊水體質(zhì)量長(zhǎng)期影響,短期內(nèi)不會(huì)產(chǎn)生劇烈變化。由于南海湖氮污染及有機(jī)物污染嚴(yán)重,因而沉積物中氮元素的量及總有機(jī)碳對(duì)細(xì)菌群落結(jié)構(gòu)影響顯著。
3 討 論
變形菌門(mén)能夠適應(yīng)復(fù)雜的生存環(huán)境,另外在變形菌門(mén)中已發(fā)現(xiàn)多種可參與氮循環(huán)的細(xì)菌,因此,其在南海湖中占據(jù)優(yōu)勢(shì)地位,綠彎菌門(mén)為兼性厭氧微生物,在冰封期水低體溶解氧的條件下依然可以保持較好的活性[14]。藍(lán)藻門(mén)并不是沉積物中的優(yōu)勢(shì)菌種,但在水體中的量較高。各采樣點(diǎn)間營(yíng)養(yǎng)水平的差異,使微生物擁有不同的生存環(huán)境,進(jìn)而影響細(xì)菌群落結(jié)構(gòu),變形菌門(mén)、綠彎菌門(mén)、放線菌門(mén)、擬桿菌門(mén)和藍(lán)藻門(mén)在4 個(gè)采樣點(diǎn)之間具有顯著差異,Thiobacillus、unclassified-p-Chloroflexi、unrank-c-Bacteroidetes-vadinHA17 等在營(yíng)養(yǎng)化水平較高區(qū)域豐度更高,放線菌門(mén)在營(yíng)養(yǎng)化水平較低區(qū)域的量較高,Liu 等[15]在對(duì)太湖進(jìn)行相關(guān)研究時(shí)也得出了類似結(jié)論。
在冰封期間,雖然低溫在一定程度上限制了細(xì)菌的活性,但其仍然在湖泊物質(zhì)循環(huán)中發(fā)揮著重要作用,且在本研究中細(xì)菌種類繁多。本次研究中,變形菌門(mén)、綠彎菌門(mén)在沉積物中有較高的量,這與薛銀剛等[16]在對(duì)太湖進(jìn)行的相關(guān)研究相似,但是與太湖相比,南海湖在冰封期獨(dú)有的冰蓋導(dǎo)致溶解氧量較低,檢測(cè)到了種類繁多的厭氧菌,如硝化螺菌屬、螺旋體屬等。硫桿菌屬是沉積物中的優(yōu)勢(shì)菌種,屬于硫氧化細(xì)菌,可將還原態(tài)硫化物和元素硫氧化為硫酸。由于冰蓋的存在使湖中污染物發(fā)生濃縮效應(yīng),污染狀況較其他時(shí)間更為嚴(yán)重,占據(jù)優(yōu)勢(shì)菌種的硫桿菌屬可以極大程度地減少水體中還原態(tài)硫化物的量,降低了水體中的污染,對(duì)減輕湖泊富營(yíng)養(yǎng)化現(xiàn)象具有積極作用[17]。
unclassified-c-Cyanobacteria 是水體中占比最高的屬。unclassified-c-Cyanobacteria 分布極廣,普遍生長(zhǎng)在淡水、海水和土壤中,并且在極端環(huán)境中也能生長(zhǎng),故有“先鋒生物”的美稱[18],因此在冰封期水體中雖然外界條件惡劣,但也能發(fā)現(xiàn)較多unclassified-c-Cyanobacteria 存在。在相關(guān)研究中unclassified-c-Cyanobacteria 類群具有固氮能力,通過(guò)氮?dú)獾墓潭ㄌ岣叩咎锱c其他土壤的肥力[19],但其在富營(yíng)養(yǎng)化嚴(yán)重的水體中具有較高的量,是水體富營(yíng)養(yǎng)化的指示生物,其中的一些屬種普遍存在于發(fā)生赤潮與水華的水體中,導(dǎo)致水體環(huán)境的惡化及水生動(dòng)物的死亡,由此可見(jiàn)南海湖中氮磷污染較為嚴(yán)重,若產(chǎn)生合適的條件極易爆發(fā)水華[20]。
造成湖泊富營(yíng)養(yǎng)化的因素有很多,情況復(fù)雜,特別是在北方冰封期,由于冰蓋的影響造成水體中溶解氧較低,并且營(yíng)養(yǎng)物質(zhì)發(fā)生沉降作用,使水體污染狀況加劇。以往研究表明,南海湖在冰封期水體已呈現(xiàn)出富營(yíng)養(yǎng)狀態(tài),沉積物中相關(guān)污染物也隨之加劇,從而對(duì)沉積物細(xì)菌群落造成影響[9]。Winter等[21]對(duì)勞倫森大湖研究后發(fā)現(xiàn)硝態(tài)氮、氨氮以及有機(jī)物量是影響其中細(xì)菌群落結(jié)構(gòu)的主要影響因子,而同樣地處我國(guó)北方寒旱區(qū)的烏梁素海中,總磷、水溶鹽總量和氨氮對(duì)其中細(xì)菌群落結(jié)構(gòu)的影響最為明顯。
unclassified-c-Cyanobacteria 為水體中各區(qū)域豐度較高的菌屬,較多unclassified-c-Cyanobacteria 的存在,說(shuō)明南海湖冰封期水體已呈現(xiàn)富營(yíng)養(yǎng)化狀態(tài),污染已十分嚴(yán)重。pH 值、COD 是冰封期unclassified-c-Cyanobacteria 的主要影響因子,控制pH 值、COD 量是避免南海湖水華爆發(fā)的重要舉措。
硫桿菌屬為沉積物中各個(gè)區(qū)域豐度較高的菌屬,其在南海湖冰封期生物物質(zhì)循環(huán)中扮演著重要的角色[22]。從相關(guān)性熱圖可以看出,TN、NH4+-N以及COD 與硫桿菌屬有較強(qiáng)的正相關(guān)。冰封期沉積物為硫桿菌屬提供了良好的生存環(huán)境,鑒于其在預(yù)防水體惡化中的重要作用,應(yīng)給予足夠重視,通過(guò)研究分析出與硫桿菌屬呈負(fù)相關(guān)的理化因子,從而控制其在水體及沉積物中的量,有效預(yù)防南海湖在冰封期水質(zhì)發(fā)生惡化。
4 結(jié) 論
1)在冰封期,溶解氧成為影響細(xì)菌物種豐富度的關(guān)鍵因素,物種豐富度依次為旅游開(kāi)發(fā)區(qū)>水草區(qū)>進(jìn)水口區(qū)>湖心區(qū)。
2)冰封期南海湖細(xì)菌主要類群分別為Proteobacteria 、 Chloroflexi 、 Cyanobacteria 、Bacteroidetes 和Acidobacteria;從屬水平分析,優(yōu)勢(shì)類群集中在Thiobacillus、unclassified-c-Cyanobacteria、unclassified-p-Chloroflexi。
3)南海湖在冰封期已經(jīng)呈富營(yíng)養(yǎng)化狀態(tài),控制TN、NH4+-N 以及COD 量是避免南海湖水華爆發(fā)的重要舉措。
