脊尾白蝦低鹽轉錄組信息挖掘
2020-11-02 02:47:07沈曄王興強曹梅
安徽農業科學 2020年19期
沈曄 王興強 曹梅

摘要 通過前期對自然海水組(鹽度31.0)和淡水組(鹽度0.2)脊尾白蝦(Exopalaemon carinicauda)轉錄組的測序及分析,篩選出低鹽脅迫下差異表達顯著的基因并進行實時定量驗證,同時進行KEGG通路富集分析,挖掘低鹽脅迫相關信號通路,以期闡明脊尾白蝦在低鹽環境下的生理調控機制。轉錄組數據分析發現,脊尾白蝦機體可能通過磺基轉移酶活性的變化來調控類固醇、激素水平。表皮蛋白AMP5樣的表達量的變化表明低鹽脅迫可能影響脊尾白蝦節間膜的形成。低鹽脅迫試驗實時定量結果顯示,基因在鰓、肝胰腺和肌肉中的表達情況各有不同,并隨時間的變化,基因的表達量也會有所波動。Hedgehog信號通路分析發現,低鹽度脅迫條件下脊尾白蝦C型凝集素和低密度脂蛋白受體相關蛋白表達上調,推測機體通過調控C型凝集素和低密度脂蛋白受體相關蛋白,促進低密度脂蛋白的代謝,水解得到的膽固醇與Smo結合,激活Hedgehog信號通路,以響應低鹽脅迫。該研究挖掘了差異表達基因相關信號通路,推測低鹽脅迫可能通過調控代謝相關酶的活性來促進脊尾白蝦的生長代謝,并且低鹽脅迫可能影響神經遞質的調控過程。
關鍵詞 脊尾白蝦;低鹽脅迫;轉錄組
中圖分類號 Q786 ?文獻標識碼 A ?文章編號 0517-6611(2020)19-0089-08
doi:10.3969/j.issn.0517-6611.2020.19.024
Abstract By sequencing and analyzing the transcriptome of Exopalaemon carinicauda in natural seawater (salinity 31.0) and fresh water (salinity 0.2) in the early stage, we screened out the genes with significant differential expression under low salt stress and carried out realtime quantitative verification. At the same time, the enrichment analysis of KEGG pathway was carried out to explore the signal pathway related to low salt stress, in order to clarify the physiological regulation mechanism of E. carinicauda in low salt environment. The results of transcriptome analysis showed that E. carinicauda might regulate steroid and hormone levels by increasing the activity of sulfotransferase. The downregulation of amp5like expression of epidermal protein suggests that low salt stress might affect the formation of internode membrane. ?The realtime quantitative results of low salt stress test showed that the expression of genes in gill, hepatopancreas and muscle were different, and with the change of time, the expression of genes would fluctuate. The analysis of hedgehog signaling pathway showed that the Ctype lectin and LDL receptor related proteins were upregulated under low salinity stress. It was speculated that the body could promote the metabolism of LDL by regulating the Ctype lectin and LDL receptor related proteins, and the hydrolyzed cholesterol binds to Smo, thus activating hedgehog signaling pathway in response to low salt stress. In this study, we explored the differentially expressed gene related signal pathways, and speculated that low salt stress might promote the growth and metabolism by regulating the activity of metabolism related enzymes, and low salt stress might affect the regulation process of neurotransmitters.
Key words Exopalaemon carinicauda;Low salinity stress;Transcriptome
基金項目 江蘇省研究生科研與實踐創新計劃項目(SJCX18_0927);江蘇省海洋生物技術重點實驗室開放基金項目(HS16005);連云港市“海燕計劃”科研項目。
作者簡介 沈曄(1994—),女,江蘇泰州人,碩士研究生,研究方向:水產動物增養殖。*通信作者,教授,博士,從事養殖生態學研究。
收稿日期 2020-02-22;修回日期 2020-03-20
鹽度是影響甲殼類生長與存活的重要環境因素,對其耗氧率和抗氧化酶活力等產生顯著影響,通過調節滲透壓使其生理活動發生變化[1]。一些甲殼動物鹽度適應范圍廣,如中國對蝦(Fenneropenaeus chinensis)和脊尾白蝦(Exopalaemon carinicauda)等[2-3]。鹽度脅迫下,脊尾白蝦孵化率和幼體成活率降低,三磷酸腺苷酶和超氧化物歧化酶活性均受到不同程度的影響[4-5]。絲氨酸蛋白酶抑制劑和蛻皮抑制激素等參與鹽度脅迫的應激反應[6-7]。轉錄組測序可以對物種在各種環境條件下進行高通量測序,有助于了解該條件下某些生命過程中相關基因的表達情況,揭示其代謝網絡及其調控機理[8-9]。張曉釵等[10]進行杜氏鹽藻轉錄組測序,探討不同濃度鹽脅迫對其生長發育以及相關信號通路的影響。郝慶玲等[11]運用Illumina 2000測序篩選牛卵泡發育調控基因。水產動物的生長與其生存環境的狀況有著密不可分的關系。轉錄組測序技術在大菱鲆(Scophthalmus maximus)、牙鲆(Paralichthys olivaceus)、青海湖裸鯉(Gymnocypris przewalskii)、大黃魚(Larimichthys crocea)、尼羅羅非魚(Tilapia nilotica)等水產動物在外界脅迫下的基因表達分析已有應用[12-16]。筆者基于以前通過第二代Illumina測序技術獲得的脊尾白蝦高通量轉錄組數據,對低鹽脅迫下的差異表達基因進行篩選,并設計引物對其進行驗證,同時挖掘代謝通路中差異基因表達情況,以期闡明脊尾白蝦在低鹽環境下的生理調控機制。
1 材料與方法
1.1 試驗材料
試驗在江蘇海洋大學海水養殖動物病害實驗室進行。試驗用蝦來自連云港南極路水產市場,均為規格整齊、體質健壯的個體,暫養7 d,以適應實驗室環境,暫養期間海水溫度24~26 ℃、鹽度31.0、pH 8.25,溶解氧含量7.6~8.1 mg/L。
1.2 低鹽脅迫試驗
隨機挑選120只脊尾白蝦,濕重(4.12±0.56)g,自然海水組(鹽度31.0)和淡水組(鹽度0.2)每組設3個重復,每個水族箱中放養20只白蝦,加入水體約30 L。待到淡水組脊尾白蝦低鹽脅迫致側臥后,再過5 min左右整體取樣,與海水組同時取樣后將樣品速凍于液氮中備用。
1.3 轉錄組測序
用液氮將脊尾白蝦研磨均勻進行總RNA提取后,檢測樣品純度、濃度和完整性合格后,合成得到片段大小合適的雙鏈cDNA,最終通過PCR富集構建文庫。檢測文庫的濃度和插入片段大小,并通過Q-PCR準確定量有效濃度,然后用Illumina HiSeq2500進行高通量測序。測序共得到13.92 Gb高質量數據,海水組(鹽度31.0)與淡水組(鹽度0.2)樣品Q30堿基百分比不小于87.93%,質量較高。
1.4 差異表達基因篩選與驗證
使用EBSeq進行差異表達分析,將錯誤發現率FDR(false discovery rate)<0.01且差異倍數FC(fold change)≥2作為篩選標準,最終得到自然海水組和淡水組樣品間差異表達基因。篩選出顯著上調和下調的前50個unigenes列表,再進一步篩選出感興趣的差異表達基因,通過實時定量PCR的方法進行檢驗。實時定量驗證試驗將鹽度驟降至0,鹽度脅迫0、0.5、1.0、2.0、4.0和6.0 h后,每個時間段分別設3個重復,分別取脊尾白蝦的鰓、肌肉、肝胰腺,液氮速凍保存。設計引物后,進行實時定量驗證。
1.5 GO富集分析
利用topGO軟件對自然海水組和淡水組樣品間差異表達基因進行GO富集分析,挖掘顯著富集節點中的差異表達基因,分析差異表達基因可能信使的分子功能。
1.6 KEGG通路富集分析
KEGG數據庫是關于Pathway的主要公共數據庫。通過GDP-L-巖藻糖合成酶樣蛋白、甘氨酸受體亞α-2樣、犬尿氨酸/α-氨基己二酸轉氨酶、C型凝集素、α-微管蛋白等差異表達基因比對到相應的通路,分析KEGG通路注釋示意圖。
2 結果與分析
2.1 差異表達基因的篩選與驗證
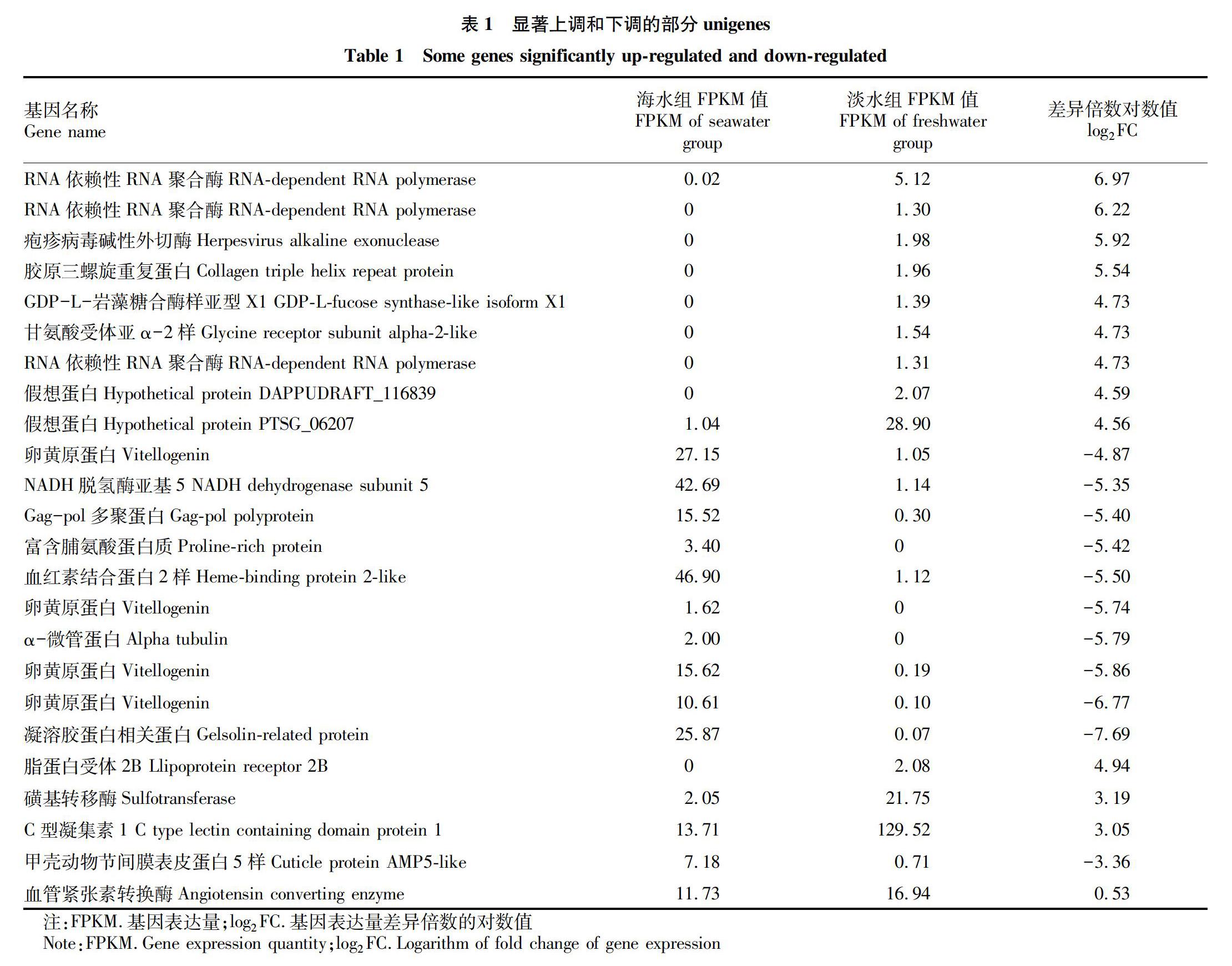
表1列出了顯著上調和下調的部分unigenes的信息,差異表達最顯著的基因分別是RNA依賴性RNA聚合酶和凝溶膠蛋白相關蛋白,log2FC分別為6.97和-7.69。從顯著上調和下調的前50個unigenes中隨機挑選脂蛋白受體2B、磺基轉移酶、C型凝集素1、甲殼動物節間膜表皮蛋白5樣和血管緊張素轉換酶5個基因進行引物設計,表2為驗證引物序列。
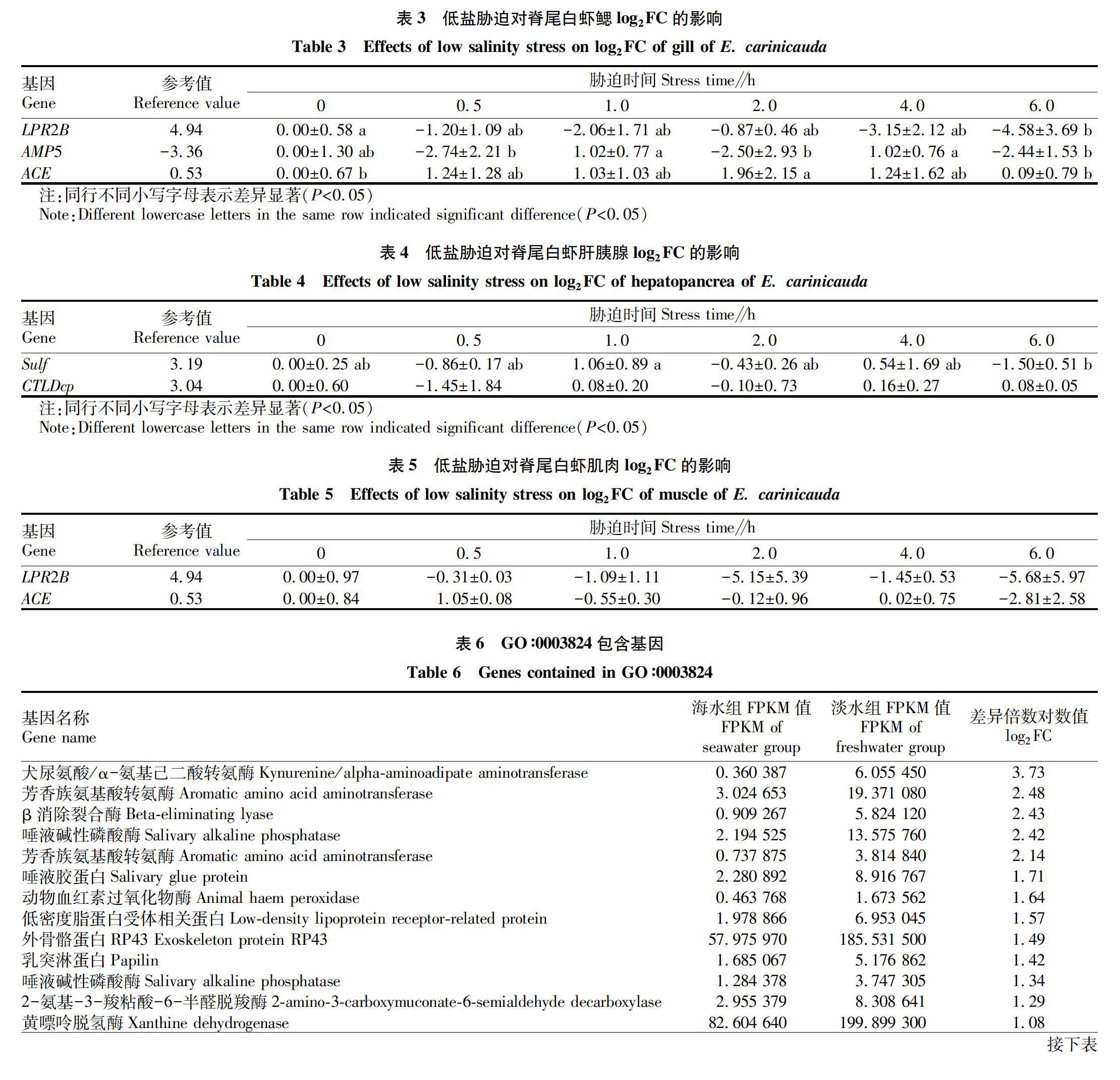
運用實時定量PCR測得差異表達基因在脊尾白蝦不同部位(鰓、肝胰腺、肌肉)中的表達量,并得到差異倍數對數值(log2FC)隨時間的變化。通過轉錄組測序得到該基因表達量差異倍數的對數值,作為參考值。如表3~5所示,LPR2B基因在鰓和肌肉中表達下調,在肌肉中差異不顯著,在肝胰腺中不表達。Sulf和CTLDcp基因表達量在肝胰腺中呈波動變化,在鰓和肌肉中不表達。AMP5基因表達量在鰓中呈波動變化,在肝胰腺和肌肉中不表達。ACE基因在鰓中表達上調,呈先上升再下降的趨勢,2.0 h時log2FC為1.96;在肌肉中呈波動變化,在肝胰腺中不表達。
2.2 GO富集分析
對海水組和淡水組的組間差異表達基因進行功能富集,主要富集到催化活性(catalytic activity)、表皮結構成分(structural constituent of cuticle)、水解酶活性(hydrolase activity)、跨膜轉運蛋白活性(transmembrane transporter activity)、脂質轉運蛋白活性(lipid transporter activity)等,其中催化活性富集顯著性最高。表6為該節點中差異表達較顯著的27個unigenes,其中表達量上調幅度最大的是犬尿氨酸/α-氨基己二酸轉氨酶,log2FC為3.73;下調幅度最大的是脂肪酶,log2FC為-2.73。
2.3 KEGG通路富集分析
GDP-L-巖藻糖合成酶樣基因比對到果糖及甘露糖代謝、氨基酸和核苷酸糖代謝等信號通路上。對果糖和甘露糖代謝通路進行展示(圖1),結果顯示低鹽度脅迫條件下,脊尾白蝦GDP-L-巖藻糖合成酶樣蛋白和木糖異構酶樣蛋白表達上調,而磷酸丙糖異構酶表達下調,以上基因的詳細信息見表7。
甘氨酸受體亞α-2樣基因比對到神經活性配體-受體相互作用信號通路上。對該通路進行展示(圖2),結果顯示低鹽度脅迫條件下脊尾白蝦甘氨酸受體亞α-2樣基因表達量顯著上調,而γ-氨基丁酸受體表達量顯著下調,以上基因的詳細信息見表8。
犬尿氨酸/α-氨基己二酸轉氨酶基因比對到賴氨酸生物合成、色氨酸代謝、賴氨酸降解信號通路上,對賴氨酸降解信號通路進行展示(圖3),結果顯示低鹽度脅迫條件下脊尾白蝦犬尿氨酸/α-氨基己二酸轉氨酶表達顯著上調,乙醛脫氫酶1表達下調,γ-丁基甜菜堿雙加氧酶表達有上調也有下調,以上基因的詳細信息見表9。
C型凝集素比對到Hedgehog信號通路上,對該信號通路進行展示(圖4),結果顯示低鹽度脅迫條件下脊尾白蝦含C α-微管蛋白比對到吞噬體信號通路上。對該通路進行展示(圖5),結果顯示低鹽度脅迫條件下脊尾白蝦α-微管蛋白、C型凝集素2表達下調,心肌肌動蛋白(cardiac muscle actin)表達下調,以上基因的詳細信息見表11。
3 討論
表皮蛋白屬于CPR(cuticular proteins with the Rebers and Riddiford Consensus)家族,參與蛻皮前后新表皮的形成,分為RR-1和RR-2兩個亞家族,其中RR-1主要存在于甲殼動物節間膜(arthrodial membranes)部分[17-18]。磺基轉移酶催化多種內源性和外源性復合物的硫化代謝過程,如類固醇、激素和生物胺類等,調節體內脂質穩態和激素水平[19]。該研究轉錄組數據分析發現,低鹽脅迫顯著提高磺基轉移酶的表達,而表皮蛋白AMP5樣表達顯著降低;據此推測,脊尾白蝦機體可能通過磺基轉移酶活性的變化來調控類固醇、激素水平,表皮蛋白AMP5樣的表達量變化表明低鹽脅迫可能影響脊尾白蝦節間膜的形成。低鹽脅迫試驗實時定量結果顯示,基因表達量的上調和下調隨時間的延長有所變化,且基因在不同組織中表達情況也有不同。這可能是由于實時定量驗證所用模板為白蝦的不同組織,且測量了0、0.5、1.0、2.0、4.0和6.0 h幾個時間點,而轉錄組測序是低鹽脅迫后單一時間點的全蝦整體混合測序。
通過差異表達基因比對到低鹽脅迫相關的信號通路有果糖及甘露糖代謝通路、神經活性配體-受體相互作用信號通路和賴氨酸降解信號通路等。GDP-L-巖藻糖是L-巖藻糖的活性核糖形式,參與巖藻糖基化反應,是糖類代謝中間體。賴氨酸為機體必需氨基酸,其中動物可吸收利用的賴氨酸為L-型賴氨酸,是水產飼料中重要的添加劑之一,在通路中被犬尿氨酸轉氨酶Ⅲ/L-谷氨酰胺轉氨酶轉變為α-酮-ε-氨基己酸,通路的中間物哌啶酸與γ-氨基丁酸受體結合并參與神經遞質的調控過程[20]。甘氨酸和γ-氨基丁酸都是主要的抑制性神經遞質,通過其受體介導發揮作用,研究顯示γ-氨基丁酸A受體能抑制甘氨酸受體[21]。該研究挖掘了上述通路中差異表達基因的表達情況,推測低鹽脅迫可能通過提高GDP-L-巖藻糖合成酶、犬尿氨酸/α-氨基己酸轉氨酶的表達,調節脊尾白蝦機體的生長代謝,并且低鹽脅迫可能影響神經遞質的調控過程。
Hedgehog信號通路在多種生理調控中起著關鍵作用,參與細胞生長、分化以及組織器官的損傷修復和免疫調節[22]。Hedgehog通路中的關鍵因素包括信號配體(Hedgehog,Hh)、受體Patched(Ptch)和Smoothened(Smo)。膽固醇是Smo的內源性配體,通過與Smo結合和共價連接起作用[23]。巨蛋白(Megalin)屬低密度脂蛋白受體家族,其配體主要有維生素結合蛋白、脂蛋白、低分子量蛋白及激素和受體相關蛋白等[24-25]。低密度脂蛋白受體是一種細胞表面糖蛋白,與低密度脂蛋白結合后經胞吞作用使膽固醇酯水解,對調節體內膽固醇平衡起重要作用[26-27]。甲殼動物不具有抗體介導的免疫反應,凝集素作為先天性免疫中一種的模式識別受體在其機體內發揮重要作用,其中C型凝集素是首次被發現的動物凝集素[28]。C型凝集素通常有至少1個C型凝集素結構域,具有識別糖類的作用。其中,含有非Ca2+依賴的C型凝集素結構域的常稱之為非經典C型凝集素,可識別蛋白質和脂類等非碳水化合物配體,如氧化低密度脂蛋白等[29]。該研究通過對Hedgehog信號通路的分析發現,低鹽度脅迫條件下脊尾白蝦C型凝集素和低密度脂蛋白受體相關蛋白均上調,推測機體通過調控低密度脂蛋白受體相關蛋白和C型凝集素,促進低密度脂蛋白的代謝,水解得到的膽固醇與Smo結合,激活Hedgehog信號通路以響應低鹽脅迫。
該研究挖掘了脊尾白蝦低鹽脅迫相關信號通路以及差異表達基因數據,獲取了脊尾白蝦低鹽脅迫相關的基因資源,豐富了脊尾白蝦轉錄組數據庫,可為今后進行脊尾白蝦低鹽脅迫生理機制的探討和關鍵基因的克隆等提供技術支撐。
參考文獻
[1] CHOI C Y,AN K W,AN M I.Molecular characterization and mRNA expression of glutathione peroxidase and glutathione Stransferase during osmotic stress in olive flounder(Paralichthys olivaceus)[J].Comparative biochemistry and physiology:Part A,2008,149(3):330-337.
[2] 雷衍之.養殖水環境化學[M].北京:中國農業出版社,2004:1-371.
[3] LI J T,MA P,LIU P,et al.The roles of Na+/K+ -ATPase αsubunit gene from the ridgetail white prawn Exopalaemon carinicauda in response to salinity stresses[J].Fish and shellfish immunology,2015,42(2):264-271.
[4] 李國峰,張振華,嚴玉洲.脊尾白蝦在低鹽度水體中的人工繁育試驗[J].水產養殖,2000(1):6-7.
[5] 李玉全,李永生,趙法箴.鹽度漸變與驟變對脊尾白蝦滲透、代謝及免疫相關酶活力的影響[J].生態學報,2015,35(21):7229-7235.
[6] 李洋,劉萍,李健,等.脊尾白蝦絲氨酸蛋白酶抑制劑基因克隆及表達分析[J].中國水產科學,2013,20(6):1166-1174.
[7] 張美.脊尾白蝦高血糖激素家族基因的克隆、表達及功能研究[D].大連:大連海洋大學,2015.
[8] VAN DIJK E L,AUGER H,JASZCZYSZYN Y,et al.Ten years of nextgeneration sequencing technology[J].Trends in genetics,2014,30(9):418-426.
[9] KLEPPE L,EDVARDSEN R B,FURMANEK T,et al.Global transcriptome analysis identifies regulated transcripts and pathways activated during oogenesis and early embryogenesis in atlantic cod[J].Molecular reproduction and development,2014,81(7):619-635.
[10] 張曉釵,李亮,何寧芳,等.不同鹽度脅迫下杜氏鹽藻全轉錄組測序及注釋[J].微生物學報,2019,59(7):1342-1353.
[11] 郝慶玲,景炅婕,朱芷葳,等.基于Illumina平臺轉錄組測序篩選牛卵泡發育調控基因[J].湖南農業大學學報(自然科學版),2019,45(1):60-67.
[12] 楊凱,黃智慧,馬愛軍,等.高溫脅迫條件下大菱鲆腎臟轉錄組研究[J].漁業科學進展,2020,41(1):86-95.
[13] 聶苗苗.大菱鲆與牙鲆耐低溫機制初探及連鎖SNP標記篩選[D].青島:中國科學院大學(中國科學院海洋研究所),2019.
[14] 周秉正,趙凱.鹽度脅迫下青海湖裸鯉組織學和轉錄組分析[C]//第八屆中國西部動物學學術研討會會議摘要匯編.成都:四川省動物學會,2019:99.
[15] 韓兆方.大黃魚轉錄組數據分析及密集脅迫前后皮膚轉錄組變化[D].廈門:集美大學,2016.
[16] 張旭,周毅,羅永巨,等.光周期影響羅非魚腦組織轉錄組基因表達分析[J].西南農業學報,2019,32(11):2704-2711.
[17] 苗澤龍,呂艷杰,張俊芳,等.日本沼蝦表皮蛋白基因的克隆及表皮組織差異性表達分析[J].河南師范大學學報(自然科學版),2018,46(2):79-83.
[18] 張宇.日本沼蝦表皮蛋白基因的克隆及分析[D].新鄉:河南師范大學,2017.
[19] 梁健,王歡,付常振,等.磺基轉移酶SULT在疾病中的作用[J].生理科學進展,2019,50(5):381-386.
[20] HALLEN A,JAMIE J F,COOPER A J L.Lysine metabolism in mammalian brain:An update on the importance of recent discoveries[J].Amino acids,2013,45(6):1249-1272.
[21] FODOR L,BOROS A,DEZSO P,et al.Expression of heteromeric glycine receptorchannels in rat spinal cultures and inhibition by neuroactive steroids[J].Neurochemistry international,2006,49(6):577-583.
[22] 閔明,王婷婷,張繼東.Hedgehog信號傳導通路及其生物學功能[J].中國免疫學雜志,2019,35(15):1903-1906,1912.
[23] HU A,SONG B L.The interplay of Patched,Smoothened and cholesterol in Hedgehog signaling[J].Current opinion in cell biology,2019,61:31-38.
[24] KUKIDA M,SAWADA H,DAUGHERTY A,et al.Megalin:A bridge connecting kidney,the reninangiotensin system,and atherosclerosis[J].Pharmacological research,2020,151:1-7.
[25] 黃愛霞.巨蛋白的研究進展[J].國外醫學(泌尿系統分冊),2002,22(4):251-254.
[26] 楊洪雁,蘇雙良,付晶,等.低密度脂蛋白受體基因研究進展[J].東北農業大學學報,2013,44(3):141-146.
[27] YANG H X,ZHANG M,LONG S M,et al.Cholesterol in LDL receptor recycling and degradation[J].Clinica chimica acta,2020,500:81-86.
[28] 王英慧.日本沼蝦C型凝集素及其配體螺原體糖基化蛋白的功能研究[D].南京:南京師范大學,2017:14-17.
[29] 謝建輝,顧建新.C型凝集素[J].生命科學,2011,23(6):555-562.
