可溶性識別分子PTX3 的研究進展
2020-11-09 06:24:36彭婉君趙彬彬劉江寧
中國比較醫學雜志 2020年1期
彭婉君,趙彬彬,武 婧,陳 鑫,牛 棟,劉江寧
(北京協和醫學院比較醫學中心,中國醫學科學院醫學實驗動物研究所,國家衛健委人類疾病比較醫學重點實驗室,北京 100021)
穿透素-3(pentraxin 3,PTX3),與C 反應蛋白(C reactive protein,CRP)、血清淀粉樣蛋白P(serum amyloid P component,SAP)同屬于正五聚蛋白家族。該家族在進化中高度保守,具有循環多聚結構的特點,是天然免疫的重要組成部分。 研究發現,相比CRP 和SAP,PTX3 的基因序列在小鼠與人中同源性極高,因此,可通過研究小鼠的PTX3 蛋白來了解該蛋白在人體中的功能情況。 PTX3 是一種典型急性期蛋白,在炎癥刺激下由炎癥細胞因子進一步誘導產生,參與對抗外源性的微生物感染。 此外,在組織修復和癌癥調節等多種生物過程中發揮著重要的作用。
1 PTX 家族
天然免疫反應是機體抵御外界微生物入侵的第一道防線,由體液反應與細胞反應兩部分組成。與適應性免疫不同,天然免疫受體是一種胚系基因編碼的受體,稱為模式識別受體(pattern recognition receptor,PRR),它識別病原體表面的保守結構,即所謂的病原體相關分子模式(pathogen-associated molecular patterns,PAMPs)。 PRR 可分為細胞相關受體和可溶性分子兩大類。 細胞相關受體包括內吞受體和信號受體,例如Toll 樣受體(Toll-like receptors, TLRs), NOD 樣受體( the NOD-like receptors)等;可溶性分子是體液天然免疫的重要組成部分, 包括補體成分、 甘露糖結合凝集素(mannose-binding lectin,MBL)、表面活性蛋白、纖維膠原素(ficolins)、正五聚蛋白(pentraxins,PTXs),它們在結構和表達方面各不相同,但都具有激活補體、中和病原體和調節凋亡細胞以及炎癥調節等功能[1]。
正五聚蛋白超家族是一種在遺傳進化上保守的多聚體蛋白家族,具有一個獨特的主基序—由200 個氨基酸組成的長結構域。 該家族依據分子量大小及結構,可分為長鏈PTX 和短鏈PTX 兩類。短鏈PTX 包括CRP 和SAP,其分子量均小于25×103;長鏈PTX 包括PTX3、PTX4、NP1(neuronal pentraxin 1)和NP2,其中,PTX3 是最早發現的長鏈PTX。
2 PTX3 的基因及蛋白結構
人類和小鼠的PTX3 基因均定位于3 號染色體,由2 個內含子和3 個外顯子組成,內顯子分別編碼蛋白的前導肽、氨基末端和C 端PTX 結構域。 該基因的近端啟動子具有多個調控元件,包括Pu1、AP-1、NF-κB、SP1 和NF-IL-6,促炎因子例如腫瘤壞死因子(tumor necrosis factor-α,TNF-α)和白細胞介素-1(interleukin-1,IL-1)可以通過結合NF-κB 對其發揮誘導作用[1]。 除此之外,PTX3 轉錄也可被PI3K/Akt 和JNK 通路,或通路下游調節因子FUS/CHOP 易位激活。 近期研究表明,表觀遺傳機制可調控PTX3 基因的表達,特別是PTX3 增強子和啟動子的高度甲基化可導致人癌癥中PTX3 基因沉默[2-3],而較低的甲基化與冠狀動脈疾病患者血漿中更高的PTX3 水平具有很好的相關性[4]。
PTX3 是由8 個亞基組成的同源多聚糖蛋白,共含有381 個氨基酸,其中包括由17 個氨基酸組成的信號肽。 其C 端結構域與CRP 和SAP 同源,8 個高度保守的氨基酸序列(HxCxS/TWxS,x 是任意氨基酸)組成了PTX 家族的標志性序列;而N 端結構域與其他已知蛋白質沒有序列相似性[5]。 CRP 和SAP 的鈣結合位點在PTX3 的五聚體結構域中缺失,這表明了不同正五聚蛋白結合特性的差異,即PTX3 與C1q 的相互作用是非鈣離子依賴的,而CRP 和SAP 會生成鈣離子黏附帶[1]。 PTX3 蛋白具有復雜的四元結構,其特征是兩個四聚體通過共價鍵連接在一起,形成一個340×103的八聚體,其與四聚體相比具有更大的功能活性[6]。 目前已在Asn220 的C 端區域發現了一個單一的N-糖基化位點,該糖苷基調控PTX3 與其他可溶性模式識別分子的相互作用,包括C1q、 factor H 和ficolin-1。 獨特的四元結構和蛋白糖基化狀態決定了PTX3 對配體的識別,并最終決定了其生物學功能。 PTX3 基因及蛋白結構示意圖見圖1。
3 PTX3 的產生
盡管PTX3 與CRP 和SAP 基因序列相似度較高,但其在來源細胞和誘導產生方面表現出重要差異。 CRP 和SAP 主要由肝臟在IL-6 的刺激下產生,而PTX3 可通過Toll 樣受體激動劑、炎性細胞因子(IL-1β 和TNF-α 等)、微生物組成成分如脂多糖(lipopolysaccharide, LPS) 或外膜蛋白 A (outer membrane protein A,OmpA)等刺激,誘導多種細胞類型進行分泌表達,其中包括樹突細胞(dendritic cells,DC)、單核細胞、巨噬細胞、成纖維細胞、滑膜細胞、軟骨細胞、脂肪細胞、上皮細胞、血管內皮細胞、平滑肌細胞、間質細胞、顆粒細胞和神經膠質細胞等。 T 淋巴細胞、B 淋巴細胞及NK 細胞均不表達PTX3,而PTX3 在成熟中性粒細胞中的表達仍存在爭議。 Jaillon 等[7]研究發現,未成熟的中性粒細胞前體在骨髓分化過程中表達PTX3 轉錄本并合成蛋白,而成熟的外周血中性粒細胞不能表達PTX3 mRNA,但PTX3 可儲存于成熟中性粒細胞中,并在微生物及炎癥信號的刺激下釋放。 而Imamura 等[8]研究表明LPS 可刺激剛分離的成熟中性粒細胞表達PTX3 mRNA。
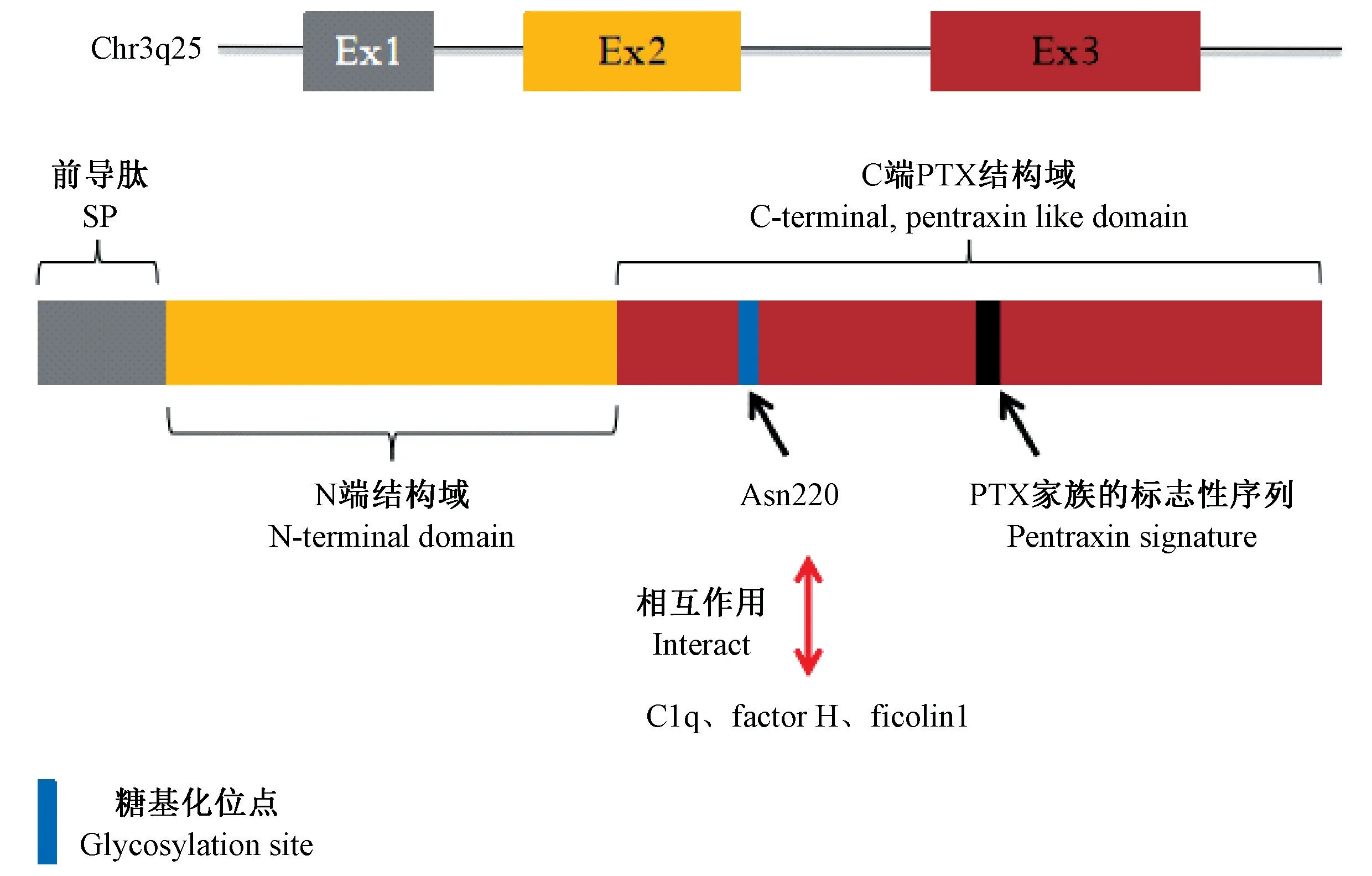
圖1 PTX3 基因及蛋白結構Figure 1 The gene and protein organization of PTX3
4 PTX3 的功能
PTX3 是體液性天然免疫系統的一種可溶性模式識別受體,被稱為“抗體前體”,可識別外源微生物,作為調理素調節炎癥反應。 PTX3 由吞噬細胞產生,由中性粒細胞儲存,經刺激后產生釋放可與部分抗原結合,被巨噬細胞或中性粒細胞上Fcγ 受體IIa(FcγRIIa/CD32)和補體受體3(CD11b/CD18)識別,從而增強其吞噬活性,促進機體天然免疫。近期有研究表明,由脾邊緣區(marginal zone,MZ)周圍的一組中性粒細胞釋放的PTX3 可與脾MZ B 細胞結合。 脾臟中性粒細胞具有粒細胞-巨噬細胞集落刺激因子 ( granulocyte-macrophage colonystimulating factor,GM-CSF)相關的功能特征,可進行免疫活化并增強PTX3 的釋放;MZ B 細胞是一群天然的抗體生成B 淋巴細胞,位于血液循環系統與適應性免疫系統之間,對T 細胞依賴抗原和非依賴抗原都能產生快速的抗體反應。 中性粒細胞分泌的PTX3 以非FcγR 依賴方式結合MZ B 細胞、B-1 細胞和不成熟的B 細胞,促進B 細胞產生的免疫球蛋白從lgM 到lgG 的轉換。 同時,PTX3 也可增強IgM和IgG 對血源性包膜肺炎鏈球菌、包膜多糖或細菌碳水化合物等的中和反應。 因此,PTX3 通過作為MZ B 細胞的內源性佐劑,連接起了天然免疫系統和適應性免疫系統[9]。
PTX3、CRP 和SAP 均可識別多種補體成分,并在多種生理病理條件下激活補體系統。 CRP 和SAP 以聚集或配體結合的形式與經典補體途徑(classical pathway,CCP)的識別單元C1q 相互作用,從而導致CCP 在病原體上的激活[10]。 而PTX3 與C1q 結合既不需要蛋白聚集也不需要鈣離子,C1q通過其球狀結構域與PTX3 相互作用,激活CCP,促進C3、C4 沉積。 此外,PTX3 也可通過與ficolin-1、ficolin-2 和MBL 的特異性相互作用調控凝集素補體通路。 Ma 等[11]發現 PTX3 可促進煙曲霉(A.fumigatus)感染后機體內ficolin -2 介導的補體沉積。這兩種補體通路途徑在下游都匯聚到C4b2a 轉化酶,涉及相同的下游分子,導致C3 轉化,C3 轉化最終可以啟動補體旁路途徑的級聯放大反應[12]。
除了促進補體激活,PTX3 也對補體激活有負調控作用。 一方面,PTX3 可在經典途徑中通過阻斷C1q 與免疫球蛋白的相互作用而抑制補體活化。另一方面,PTX3 是一種獨特的H 因子(Factor H,FH)的配體,它識別FH 的兩個結合位點,SCR7 和SCR19-20[13]。 而FH 是一種抑制因子,PTX3 可募集FH,同時二者的結合不會干擾FH 的抑制作用,因此這種結合有助于防止補體過度活化,在炎癥的控制中發揮重要作用。 研究發現PTX3 的分泌可由氧化應激誘導,而氧化應激是視網膜色素上皮細胞的年齡相關性黃斑變性( age-related macular degeneration,AMD)的主要病因之一。 PTX3 缺乏癥患者體內由于PTX3 的缺失而放大了這種氧化應激所引起的補體活化,造成癥狀加重。 這表明PTX3可通過與視網膜色素上皮細胞中的FH 相互作用,抑制補體和炎性小體的過度激活[14]。
上述的調節補體功能是PTX3 對炎癥反應發揮調控作用的主要機制之一。 除了參與補體活化外,PTX3 還通過調控炎癥細胞的募集來參與炎癥反應。 該效應由PTX3 與粘附分子P-選擇素(Pselectin)的相互作用來介導,二者結合可減少炎癥部位對中性粒細胞募集,抑制白細胞滾動進而抑制炎癥細胞浸潤[15]。 近期有研究表明,PTX3 缺乏癥會導致氣道高反應性增強、黏液生成增多和IL-17a主導的肺部炎癥加劇,說明PTX3 在過敏性炎癥的發展過程中具有調節作用[16]。
5 PTX3 在人類疾病中的作用
5.1 感染性疾病
PTX3 參與機體抵抗微生物入侵的免疫反應,通過不同的機制發揮其抗感染作用,包括調理作用、促進吞噬、調節補體活性、與抗菌蛋白相互作用等。 最早發現PTX3 抗真菌感染功能的是其對煙曲霉(Aspergillus fumigatus) 和銅綠假單胞菌(Pseudomonas aeruginosa)感染的抑制作用,PTX3 與兩種真菌結合增強巨噬細胞與中性粒細胞對其識別能力,促進細胞吞噬功能,發揮調理作用。 近期Ciancarella 等[17]研究發現PTX3 有助于機體細胞外志賀氏菌(Shigella)的清除,志賀氏菌是一種侵染結腸和直腸粘膜的人類腸道致病菌,可引起有害的炎癥反應。 肺炎球菌感染可使肺上皮細胞高表達PTX3,同時,PTX3 高表達使相關細胞因子釋放增加,因此PTX3 對細菌感染也存在調控作用。
在抗病毒入侵方面,Reading 等[18]發現人類和小鼠的PTX3 可與流感病毒結合,介導一系列抗病毒反應,包括抑制血細胞凝集、中和病毒和抑制病毒神經氨酸酶的活性。 PTX3 的抗病毒活性通過其唾液酸配體與病毒血凝素糖蛋白結合來發揮作用。因此,PTX3 在宿主感染早期具有新的抗病毒作用,可作為潛在的治療靶點。
另一方面,PTX3 也可作為多種感染性疾病的生物標志物。 肺曲霉病、肺結核、登革熱病毒感染、腦膜炎球菌病、鉤端螺旋體病、志賀氏菌病、膿毒癥和病毒性呼吸道感染患者血漿中PTX3 水平顯著升高,與疾病的嚴重程度有顯著相關性,可作為預后的重要預測因子[19-20]。 通過研究慢性丙型肝炎(HCV)患者抗病毒治療前PTX3 與炎癥活性、纖維化的生化和病理學參數的關系,發現HCV 患者的PTX3 水平隨著肝纖維化程度的加劇而升高;因此PTX3 作為顯著纖維化的單一生物標志物,提供了新的臨床診斷指標[21]。 PTX3 在感染性疾病中的作用示意圖見圖2。

圖2 PTX3 在感染性疾病中的作用Figure 2 Role of PTX3 in infection
5.2 癌癥
慢性炎癥,尤其是由微生物感染引起的炎癥反應與癌癥密切相關,同時癌癥相關炎癥是導致癌細胞遺傳異質性的重要原因之一。 PTX3 具有調節炎癥反應的功能,提示其在控制腫瘤發展方面也能發揮相應作用。
利用平滑肌肉瘤疾病模型,Eduardo 等[3]發現PTX3 可招募并結合FH,調節肉瘤細胞上的C3 沉積,促進C5a、CCL2 釋放,進而減少M2 表型的巨噬細胞聚集,通過調節補體依賴的腫瘤相關炎癥,在小鼠和人體內發揮外源性抑癌基因作用。 通過對經活檢確診為小細胞肺癌( small-cell lung carcinoma,SCLC)患者的石蠟包埋標本進行PTX3表達量檢測,發現PTX3 的高表達與SCLC 腫瘤細胞的侵襲行為有相關性,表明它可能是SCLC 的一種預后標志物,同時也是SCLC 治療的潛在分子靶點[22]。
最近研究表明,PTX3 不僅具有抑制癌癥的保護作用,也有促進腫瘤細胞遷移、侵襲和增加巨噬細胞趨化的負調節作用。 例如,胃癌來源的PTX3 可促進巨噬細胞的募集,可能促進胃癌相關炎癥的發展[23]。 在頭頸部鱗狀細胞癌(head and neck squamous cell carcinomas,HNSCCs)中,油酸通過AKT/NF-κB 通路誘導PTX3 產生,而PTX3 提高了MMP-3 和波形蛋白的表達,增強了腫瘤細胞與內皮細胞的相互作用,促進了腫瘤的遷移與侵襲[24]。 PTX3 在癌癥中的作用示意圖見圖3。

圖3 PTX3 在癌癥中的作用Figure 3 Role of PTX3 in cancer
5.3 心血管疾病
心血管疾病(Cardiovascular diseases,CVD)是一種嚴重危險人類健康的疾病,位于全球各種死因的首位;在我國心血管疾病的形勢也日趨嚴重。 研究發現,PTX3 對心血管系統具有明顯的調節作用,主要通過影響炎癥反應、血管生成、腫瘤發生、細胞粘附等多種生理過程[25]。
經流行病學和臨床病例分析,認為PTX3 是動脈粥樣硬化的有效生物標志物。 炎癥是導致年輕和老年受試者動脈粥樣硬化的主要有害因素,而PTX3 在調節炎癥反應中發揮著重要作用,其在血漿中的高水平與冠狀動脈粥樣硬化性的嚴重程度成正相關[26]。 在動脈血栓形成的小鼠模型中,源自血栓內和血管壁的血管內皮細胞的PTX3,通過其N端結構域靶向結合纖維蛋白原,抑制血小板粘附和聚集,從而阻止血栓形成[27]。 然而,Shindo 等[28]發現PTX3 在動脈粥樣硬化血栓形成的發病機制中也具有不良作用。 PTX3 可增加組織因子(tissue factor,TF)在單核細胞和內皮細胞中的表達;而作為凝血級聯反應的主要調節因子,TF 水平的升高直接導致血栓的形成。 此外,PTX3 在小鼠腦缺血損傷模型中也發揮保護作用,并參與水腫消退和膠質瘢痕形成[29]。
研究發現在急性心肌梗死(Acute myocardial infarction,AMI)患者發病約7 h 后,血漿PTX3 水平顯著升高,3 天后降低至基線水平;因此PTX3 可作為缺血性心肌病的心肌細胞不可逆損傷的早期指標。 此外,考慮到其在血漿中的濃度在心肌梗死發生時發生快速變化,PTX3 被認為是一種新的潛在的心肌梗死標志物[25]。 對于進行藥物支架置入術的冠狀動脈疾病患者和心絞痛患者,PTX3 也可作為一種預后標志物[30]。
5.4 組織重塑與修復
PTX3 在組織修復與重塑中起著不可替代的作用,分析PTX3 缺陷小鼠不同的組織損傷模型,包括皮膚傷口愈合、無菌肝肺損傷等,發現PTX3 缺陷導致纖維蛋白沉積和持久性增加、凝塊變厚,膠原沉積增加。 PTX3 通過與臨時基質蛋白即纖維蛋白相互作用,參與組織修復的調控,TLRs 和IL-1 是其有效誘導因子,在組織損傷和修復過程中出現的酸性pH 值可能是PTX3 功能的一個開關信號[31]。Vezzoli 等[32]研究表明,PTX3 在急性無菌損傷的骨骼肌再生過程中發生表達上調,PTX3 可減少炎癥反應時間,加快肌肉細胞碎片清除,加快骨骼肌損傷愈合時間。 另有研究者比較PTX3 在骨質疏松癥、骨關節炎患者和正常對照年輕受試者成骨細胞中的表達和功能,發現PTX3 在骨內平衡中參與了成骨細胞的增殖、分化[33]。 在急性-慢性肝損傷中,PTX3 受脂多糖誘導表達上調,介導肝星狀細胞損傷愈合反應[34]。
此外,PTX3 也參與了血管生成的調控。 血管生成受促血管生成因子和抗血管生成因子之間的平衡以及細胞外基質蛋白相互作用的調控。 成纖維細胞生長因子2(Fibroblast growth factor 2,FGF2)是一種主要的促血管生成因子,PTX3 可抑制FGF2對血管生成的促進作用。 而腫瘤壞死因子刺激因子-6(tumor necrosis factor-stimulated gene-6,TSG-6)通過與FGF2 競爭結合PTX3,可逆轉PTX3 對FGF2介導的血管生成的抑制作用[35]。 近期有研究者發現微粒物質中的內毒素誘導誘導調控PTX3 和血管內皮生長因子釋放[36]。
6 展望
PTX3 是一種可溶性模式識別分子,通過調節補體活化等生物功能,促進機體對病原體的識別,是抵抗真菌、細菌和病毒感染的重要介質,也是炎癥反應的調節因子,并可作為多種疾病的生物標志物。 目前已發現PTX3 在病原體感染、癌癥、心血管疾病等多種人類疾病中發揮著重要作用,在生理病理學中具有深遠意義,深入研究PTX3 的功能可為多種疾病臨床診斷和治療鋪平道路。
