基于mtDNACOI和mtDNACOⅡ分子標(biāo)記的貴州省茶棍薊馬種群遺傳多樣性分析
2020-11-09 03:09:31羅林麗孟澤洪李帥趙興麗周羅娜賀圣凌周玉鋒
南方農(nóng)業(yè)學(xué)報(bào) 2020年7期
關(guān)鍵詞:分析
羅林麗 孟澤洪 李帥 趙興麗 周羅娜 賀圣凌 周玉鋒
摘要:【目的】研究貴州省茶棍薊馬不同地理種群間的遺傳多樣性,為掌握貴州省茶棍薊馬的遺傳動(dòng)態(tài)和擴(kuò)散規(guī)律及制定防控措施提供理論依據(jù)。【方法】以貴州省11個(gè)不同地理區(qū)域的茶棍薊馬種群mtDNA CO I和mtDNA CO II基因序列為靶標(biāo),利用MEGA 6.0、DnaSP 5.10、Arlequin 3.5.2.2和Network 2.0等軟件對(duì)種群遺傳分化、基因流水平、分子變異及種群遺傳多樣性等進(jìn)行分析。【結(jié)果】基于mtDNA CO I和mtDNA CO II基因分析時(shí),分別檢測(cè)到11種和9種單倍型,各地理種群的單倍型多樣度(Hd)較高,分別為0.609和0.633;總體遺傳固定指數(shù)(FST)分別為0.11902和0.09052,基因流(Nm)分別為2.00和2.51,表明貴州省茶棍薊馬各地理種群間的基因交流水平較高,種群間遺傳分化較小。mtDNA CO I和mtDNA CO II基因序列的單倍型網(wǎng)狀樹分析均無明顯分支;種群間的分子變異(AMOVA)分析結(jié)果表明,茶棍薊馬的遺傳變異主要來自種群內(nèi)部。總?cè)后w的中性檢驗(yàn)Fus Fs不顯著(P>0.05),且錯(cuò)配分布曲線呈現(xiàn)多峰,說明貴州省茶棍薊馬種群在較近的歷史時(shí)期內(nèi)保持相對(duì)穩(wěn)定,未經(jīng)歷明顯的種群擴(kuò)張,或處于臨界狀態(tài)。【結(jié)論】貴州省茶棍薊馬種群遺傳多樣性較豐富,雖然尚未形成明顯的種群擴(kuò)張,但可能已處于臨界狀態(tài),應(yīng)積極采取綜合防控措施,力爭(zhēng)將茶棍薊馬種群數(shù)量控制在危害水平以下,以保證貴州省茶產(chǎn)業(yè)生產(chǎn)健康可持續(xù)發(fā)展。
關(guān)鍵詞: 茶棍薊馬;mtDNA CO I基因;mtDNA CO II基因;地理種群;遺傳多樣性
中圖分類號(hào): S433.89? ? ? ? ? ? ? ? ? ?文獻(xiàn)標(biāo)志碼: A 文章編號(hào):2095-1191(2020)07-1684-09
Abstract:【Objective】The study was conducted to investigate genetic diversity among different geographical populations of Dendrothrips minowai Priesner in Guizhou, so as to provide theoretical foundation for the analysis of population genetic dynamics, population dispersion mechanism and formulating control measures. 【Method】The mtDNA CO I and mtDNA CO II sequences of D. minowai populations, which collected from 11 different geographical areas in Guizhou, were selected as molecular markers, population genetic differentiation, gene flow, molecular variance and genetic diversity of populations were analyzed by using software MEGA 6.0, DnaSP 5.10, Arlequin 3.5.2.2, Network 2.0 etc. 【Result】A total of 11 and 9 types of haplotype were detected when the mtDNA CO I and mtDNA CO II gene were analyzed, respectively, haplotype diversity(Hd) of different geographic population were high, which were 0.609 and 0.633, respectively. The total genetic fixations index(FST) were 0.11902 and 0.09052,the total gene flow(Nm) were 2.00 and 2.51, respectively, which indicated that the level of gene communication was high and the genetic differentiation among populations were small. The haplotype network of different geographical populations based on mtDNA CO I and mtDNA CO II showed no obvious branches, and the analysis of molecular variance(AMOVA) of different geographical populations indicated that the genetic variation mainly occurred within the same population. The total neutral test showed that Fus Fs were not significant(P>0.05), and the mismatch distribution curve had multiple peaks, which indicated the population of D. minowai in Guizhou were relatively stable in recent historical period, and has not experienced evident expansion or at critical state. 【Conclusion】The genetic diversity of D. minowai populations in Guizhou is relatively rich, which has not yet formed an obvious sudden expansion, but might approach to critical phase. Therefore, comprehensive prevention and control measures should be taken actively to control the D. minowai populations quantity under damage level and ensure the sustainable and healthy development of tea production in Guizhou.
Key words: Dendrothrips minowai Priesner; mtDNA CO I gene;? mtDNA CO II gene; geographical population; genetic diversity
Foundation item: National Natural Science Foundation of China(31560515); National Modern Agricultural Industry Technology System Construction Project(CARS-19); Guizhou Science and Technology Plan Project(NY〔2015〕3011-2)
0 引言
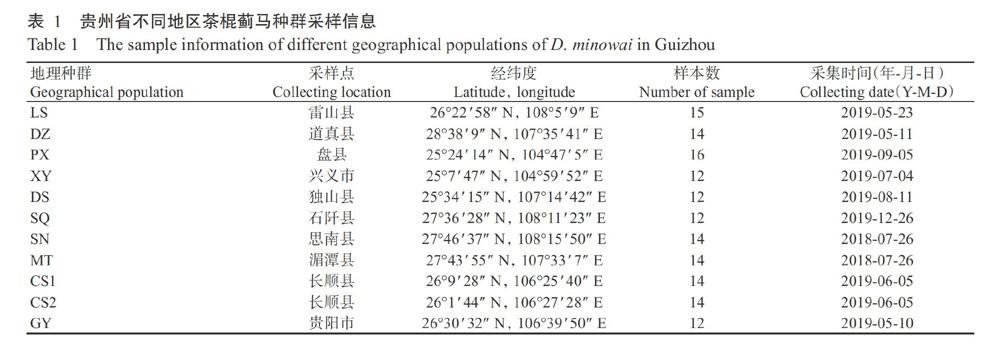
【研究意義】茶棍薊馬(Dendrothrips minowai Priesner)又稱茶棘皮薊馬,屬纓翅目(Thysanoptera)薊馬科(Thripidae)(唐美君和肖強(qiáng),2018),在我國(guó)主要分布于浙江、福建、廣東、廣西、海南和貴州等南方茶區(qū),在山東等也有少量報(bào)道(王洪濤等,2018;李貌等,2020)。近年來,茶棍薊馬在貴州省多地為害且逐年加重,個(gè)別區(qū)域暴發(fā)成災(zāi),嚴(yán)重影響茶葉的產(chǎn)量和品質(zhì),成為影響貴州省茶葉生產(chǎn)的主要害蟲之一(彭萍等,2013;呂召云等,2015)。貴州省地貌以山地居多,受多種因素的影響,氣候呈現(xiàn)多樣性,生物多樣性及群體遺傳多樣性也極其豐富,在此基礎(chǔ)上探索茶棍薊馬種群內(nèi)在的遺傳變異與進(jìn)化關(guān)系等,能從遺傳學(xué)角度為有效防控茶棍薊馬提供理論指導(dǎo)。【前人研究進(jìn)展】目前,針對(duì)薊馬遺傳多樣性的研究多集中在入侵性害蟲西花薊馬上。武曉云等(2009)利用PCR克隆并分析了西花薊馬rDNA ITS2和mtDNA CO I基因序列的進(jìn)化速率,發(fā)現(xiàn)rDNA ITS2序列的變異程度比mtDNA CO I高,提出mt-DNA CO I較rDNA ITS2更適合應(yīng)用于西花薊馬的種內(nèi)遺傳分析;Brunner和Frey(2010)利用mtDNA CO I基因和微衛(wèi)星分子標(biāo)記對(duì)美國(guó)西部地區(qū)西花薊馬種群的遺傳結(jié)構(gòu)進(jìn)行分析,發(fā)現(xiàn)在起源地西花薊馬種群發(fā)生了分化,且這種遺傳分化與形態(tài)上的分化不一致,推測(cè)這種分化是由棲息地的環(huán)境不同(地理隔離)造成;喬瑋娜(2012)運(yùn)用DNA條形碼技術(shù)對(duì)我國(guó)13個(gè)省(市)不同地理種群西花薊馬的175條mt-DNA CO I基因序列進(jìn)行遺傳多樣性和遺傳分化程度分析,推測(cè)我國(guó)西花薊馬各地理種群的可能來源;Yang等(2012a,2012b)用微衛(wèi)星和mtDNA CO I分子標(biāo)記研究西花薊馬在我國(guó)的入侵遺傳學(xué),發(fā)現(xiàn)傳入我國(guó)的西花薊馬在遺傳多樣性上表現(xiàn)出一個(gè)明顯的瓶頸效應(yīng),揭示西花薊馬可能首先傳入我國(guó)云南,隨后云南成為我國(guó)其他西花薊馬種群的源頭。沈登榮等(2014)對(duì)薊馬科9個(gè)屬27種的mtDNA-CO I基因序列變異進(jìn)行分析,首次報(bào)道了茶棍薊馬的mtDNA-CO I基因序列。目前茶棍薊馬雖然已在貴州等多地為害,但其研究?jī)H限于形態(tài)(王洪濤等,2018)、空間分布(張莉等,2019)和防治措施(趙志清和陳流光,1998;李慧玲等,2014;涂娟等,2016;董照鋒和張小平,2018;曾明森,2019)等方面,鮮少有利用分子標(biāo)記技術(shù)探討茶棍薊馬遺傳變異方面的相關(guān)研究,僅Lü等(2016)利用mtDNA CO I作為分子標(biāo)記研究了貴州茶棍薊馬的遺傳結(jié)構(gòu)與多樣性,認(rèn)為貴州省茶棍薊馬種群在近期發(fā)生了種群擴(kuò)張。【本研究切入點(diǎn)】線粒體基因已廣泛運(yùn)用于動(dòng)物(尤其是昆蟲)分子進(jìn)化與系統(tǒng)發(fā)育研究(Dickey et al.,2015;Lü et al.,2016)。線粒體基因中的3個(gè)細(xì)胞色素氧化酶亞基編碼基因(CO I、CO II和CO III)進(jìn)化速率不同,CO II基因核苷酸序列及其推導(dǎo)氨基酸序列的進(jìn)化速率均比其他2個(gè)亞基(CO I和CO III)高(Peterson et al.,2001)。本研究在Lü等(2016)的研究成果基礎(chǔ)上,選用mtDNA CO I和mtDNA CO II基因作為分子標(biāo)記,同時(shí)增加每個(gè)地理種群的測(cè)序樣本數(shù),分析貴州省茶棍薊馬不同地理種群的遺傳變異與進(jìn)化關(guān)系。【擬解決的關(guān)鍵問題】對(duì)貴州省11個(gè)地理種群茶棍薊馬群體mtDNA CO I和mtDNA CO II基因的部分序列進(jìn)行測(cè)序,研究貴州省茶棍薊馬群體的遺傳多樣性及遺傳分化情況,探討群體內(nèi)在的遺傳變異與進(jìn)化關(guān)系,為掌握貴州省茶棍薊馬的遺傳動(dòng)態(tài)和擴(kuò)散規(guī)律及制定防控措施提供理論依據(jù)。
1 材料與方法
1. 1 試驗(yàn)材料
以貴州省不同地理種群的茶棍薊馬為供試樣本。在野外用小毛筆直接蘸取茶樹上的茶棍薊馬,將其放入盛有無水乙醇的樣本管中,記錄樣本采集時(shí)間、采集地點(diǎn)及其經(jīng)緯度等信息,樣本帶回實(shí)驗(yàn)室在顯微鏡下進(jìn)行鑒定后置于-20 ℃保存。具體采樣信息見表1。
1. 2 基因組DNA提取
茶棍薊馬總DNA的提取參照張利娟等(2011)的鹽析法,提取單頭薊馬DNA,提取的DNA以20 μL無菌超純水充分溶解后-20 ℃保存?zhèn)溆谩?/p>
1. 3 PCR擴(kuò)增與序列測(cè)定
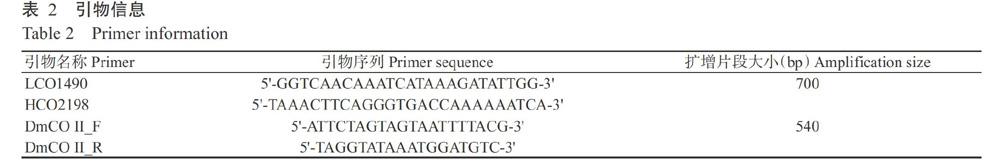
mtDNA CO I基因序列擴(kuò)增使用DNA條形碼標(biāo)準(zhǔn)引物L(fēng)CO1490/HCO2198(Folmer et al.,1994);參照NCBI數(shù)據(jù)庫(kù)中茶棍薊馬線粒體基因組DNA全長(zhǎng)序列中的mtDNA CO II區(qū)域,設(shè)計(jì)mtDNA CO II基因引物DmCO II_F/DmCOI I_R(表2)。反應(yīng)體系50.0 μL:TaKaRa PrimeSTAR HS DNA Polymerase 0.5 μL,5×PrimerSTAR Buffer 10.0 μL,dNTP Mixture 4.0 μL,上、下游引物各2.0 μL,DNA模板3.0 μL,無菌雙蒸水28.5 μL。mtDNA CO I擴(kuò)增程序:94 ℃預(yù)變性5 min;94 ℃ 1 min,53 ℃ 1 min,72 ℃ 80 s,進(jìn)行32個(gè)循環(huán);72 ℃延伸10 min,4 ℃保存。mtDNA CO II擴(kuò)增程序:94 ℃預(yù)變性5 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,進(jìn)行32個(gè)循環(huán);72 ℃延伸10 min,4 ℃保存。取2.0 μL PCR產(chǎn)物進(jìn)行電泳檢測(cè)后,對(duì)確定含有目的片段的產(chǎn)物進(jìn)行雙向測(cè)序(引物合成和測(cè)序由北京擎科新業(yè)生物技術(shù)有限公司完成)。
1. 4 序列分析
利用Chromas 2.23對(duì)測(cè)序所得序列進(jìn)行峰圖質(zhì)量驗(yàn)證,對(duì)序列進(jìn)行人工讀取和反復(fù)校驗(yàn)后,將正、反向序列進(jìn)行拼接得到的完整序列片段在NCBI數(shù)據(jù)庫(kù)中進(jìn)行同源比對(duì),以確定序列是否為所需的目的片段。
1. 5 遺傳學(xué)參數(shù)計(jì)算與分析
利用MEGA 6.0分析序列的堿基組成與多態(tài)性位點(diǎn)、轉(zhuǎn)換/顛換偏倚率等(Tamura et al.,2013);通過DnaSP 5.10計(jì)算核苷酸多樣度(π)、單倍型多樣度(Hd)、核苷酸平均差異數(shù)(K)及種群間遺傳固定指數(shù)(FST)、基因流(Nm)等遺傳學(xué)參數(shù)(Librado and Rozas,2009)。利用SAMOVA 2.0(Dupanloup et al.,2002)對(duì)不同地理種群間的分子空間變異進(jìn)行分析,然后根據(jù)SAMOVA分組結(jié)果,應(yīng)用Arlequin 3.5.2.2進(jìn)行中性檢驗(yàn)、分子遺傳變異方差(AMOVA)和錯(cuò)配分布分析(Excoffier and Lischer,2010)。使用Network 5.0繪制基于Median-joining的單倍型中介網(wǎng)絡(luò)圖(Bandelt et al.,1999)。
2 結(jié)果與分析
2. 1 茶棍薊馬mtDNA CO I和mtDNA CO II基因序列特征分析結(jié)果
11個(gè)茶棍薊馬種群的mtDNA CO I基因經(jīng)雙向拼接和ClustalW序列比對(duì)分析后得到137條序列,片段長(zhǎng)度665 bp,共檢測(cè)到17個(gè)多態(tài)性位點(diǎn),包括11個(gè)自裔位點(diǎn)和6個(gè)簡(jiǎn)約信息位點(diǎn);所有序列的總突變數(shù)為17,其中發(fā)生轉(zhuǎn)換的位點(diǎn)數(shù)為15,發(fā)生顛換的位點(diǎn)數(shù)為5,轉(zhuǎn)換/顛換率為18.741;所有序列核苷酸A、T、C和G的平均含量分別為31.73%、37.95%、16.64%和13.68%,A+T含量為69.68%,序列堿基組成有明顯的A/T偏好性。mtDNA CO II基因經(jīng)雙向拼接和比對(duì)分析后得到132條序列,片段長(zhǎng)度525 bp,共檢測(cè)到16個(gè)多態(tài)性位點(diǎn),包括9個(gè)自裔位點(diǎn)和7個(gè)簡(jiǎn)約信息位點(diǎn);所有序列的總突變數(shù)為16,其中發(fā)生轉(zhuǎn)換的位點(diǎn)數(shù)為13,發(fā)生顛換的位點(diǎn)數(shù)為3,轉(zhuǎn)換/顛換率為5.797;所有序列核苷酸A、T、C和G的平均含量分別為35.24%、37.27%、14.55%和12.95%,A+T含量為72.51%,序列堿基組成有明顯的A/T偏好性。
2. 2 茶棍薊馬單倍型及遺傳多樣性分析結(jié)果
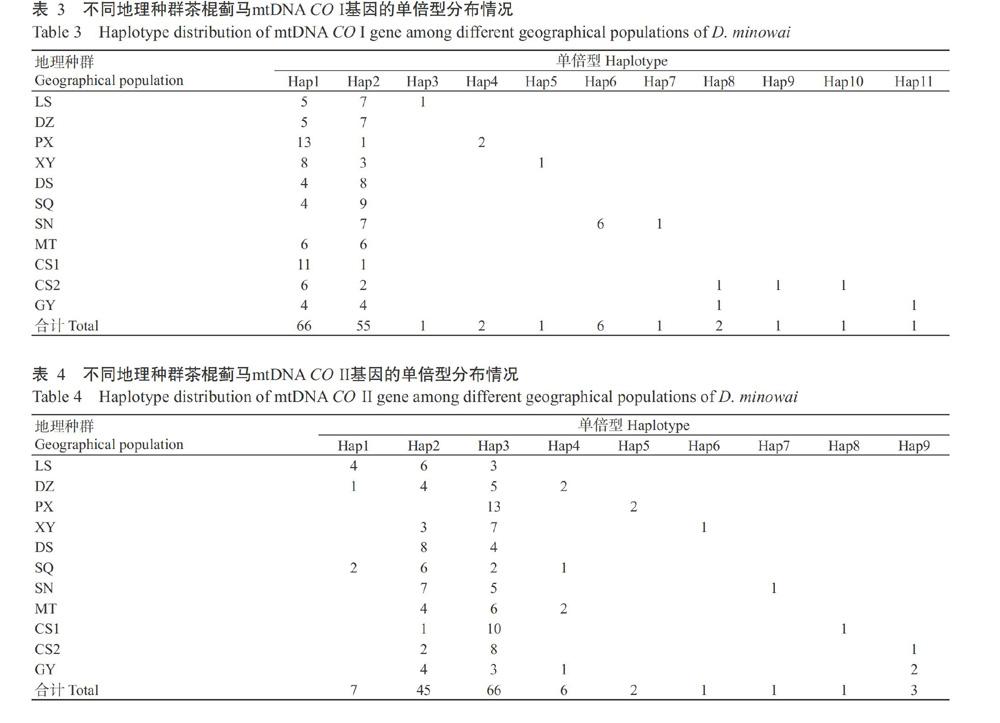
以mtDNA CO I為靶標(biāo)基因時(shí),在測(cè)序成功的137條序列中共觀測(cè)到11種單倍型(Hap1~Hap11),其中,Hap2為共享單倍型,在11個(gè)茶棍薊馬種群中均有分布,Hap1為優(yōu)勢(shì)單倍型,在除SN外的10個(gè)種群中均有分布;Hap1和Hap2序列共計(jì)121條,占序列總數(shù)的88.32%;種群?jiǎn)伪缎蛿?shù)的范圍為2~5,CS2種群的單倍型數(shù)最多(表3)。以mtDNA CO II為靶標(biāo)基因時(shí),在測(cè)序成功的132條序列中共觀測(cè)到9種單倍型(Hap1~Hap9),其中,Hap3為共享單倍型,在11個(gè)茶棍薊馬種群中均有分布,Hap2在除PX外的10個(gè)種群中均有分布;Hap2和Hap3序列共計(jì)111條,占序列總數(shù)的84.09%;種群?jiǎn)伪缎蛿?shù)的范圍為2~4(表4)。
基于mtDNA CO I基因進(jìn)行分析,結(jié)果(表5)顯示,貴州省11個(gè)茶棍薊馬地理種群總體Hd為0.609,Hd范圍0.167~0.733,其中GY種群的Hd最大,CS1種群的Hd最小;總體π為0.00129,π范圍為0.00025~0.00213;總體K為0.859,K范圍為0.167~1.418;π和K均以CS2種群最大、CS1種群最小。基于mtDNA CO II基因進(jìn)行分析,結(jié)果(表6)顯示,貴州省11個(gè)茶棍薊馬地理種群總體Hd為0.633,Hd范圍為0.248~0.778;總體π為0.00188,π范圍為0.00047~0.00347;總體K為0.985,K范圍為0.248~1.820;GY種群的Hd、π和K均最大,而PX種群的最小。
2. 3 遺傳分化與分子變異分析結(jié)果
將11個(gè)茶棍薊馬種群作為一個(gè)整體進(jìn)行分析,以mtDNA CO I基因?yàn)榘袠?biāo)時(shí),總?cè)后w的FST為0.11902,變異范圍為-0.08556~0.52427;總?cè)后w的Nm為2.00,變異范圍為-36.02~58.57(表7)。以mtDNA CO II基因?yàn)榘袠?biāo)時(shí),總?cè)后w的FST為0.09052,變異范圍為-0.08211~0.54221;總?cè)后w的Nm為2.51,變異范圍為-215.77~182.23(表8)。綜合分析表明,貴州省不同地理區(qū)域的茶棍薊馬種群間基因交流水平較高,種群間遺傳分化較小。但mtDNA CO I基因和mtDNA CO II基因反映各地理種群間的遺傳分化水平并不完全一致。如以mtDNA CO I為靶標(biāo)基因的分析結(jié)果顯示,SN種群與DS和SQ種群間可能存在中度分化;而以mtDNA CO II為靶標(biāo)基因顯示,彼此間無明顯遺傳分化。
在SAMOVA 2.0中對(duì)不同地理種群茶棍薊馬間的分子空間變異進(jìn)行分析,將各種群的地理信息與遺傳信息結(jié)合,得到可能的種群組合方式,結(jié)果顯示mtDNA CO I和mtDNA CO II基因的分組結(jié)果一致,均以K=2時(shí)的分組結(jié)果最佳,即將11個(gè)地理種群分成兩組,PX、XY、CS1和CS2為第1組,LS、DZ、DS、SQ、SN、MT和GY為第2組。mtDNA CO I和mtDNA CO II基因的AMOVA分析結(jié)果顯示(表9),同一種群內(nèi)的遺傳變異分別占總體變異的79.72%和81.63%,各種群內(nèi)的FST均呈顯著性差異(P<0.01),2個(gè)基因的分析均表明種群遺傳變異主要來自種群內(nèi)部。此外,2個(gè)基因分析均表明,不同組間的遺傳變異(FCT)均表現(xiàn)出顯著性差異(P<0.01),組內(nèi)種群間的遺傳分化顯著,而組內(nèi)種群間的遺傳變異(FSC)均表現(xiàn)為差異不顯著(P>0.05,下同),組內(nèi)種群間的遺傳變異不明顯。
2. 4 單倍型系統(tǒng)發(fā)育分析結(jié)果
基于Median-joining構(gòu)建的單倍型中介網(wǎng)絡(luò)樹分析結(jié)果(圖1)顯示,mtDNA CO I基因與mtDNA CO II基因的單倍型中介網(wǎng)絡(luò)圖結(jié)果基本一致,貴州省11個(gè)茶棍薊馬地理種群均未表現(xiàn)出明顯分支。mtDNA CO I基因序列定義了11種單倍型(3個(gè)共享單倍型和8個(gè)獨(dú)享單倍型),其中,Hap1為其他單倍型的核心,出現(xiàn)頻率較高的是Hap1和Hap2,為優(yōu)勢(shì)單倍型,基本分布在每個(gè)地理種群中,其他獨(dú)享單倍型通過一步或幾步突變與優(yōu)勢(shì)單倍型相連。mtDNA CO II基因序列定義了9種單倍型(4個(gè)獨(dú)享單倍型和5個(gè)共享單倍型),其中,Hap3為其他單倍型的核心,Hap2和Hap3出現(xiàn)頻率較高,為優(yōu)勢(shì)單倍型,基本分布在每個(gè)地理種群中,其他獨(dú)享單倍型通過一步或幾步突變與Hap2和Hap3相連。
2. 5 種群動(dòng)態(tài)分析結(jié)果
從mtDNA CO I和mtDNA CO II基因的Taijimas D和Fus Fs中性進(jìn)化模型檢驗(yàn)結(jié)果(表5和表6)可知,11個(gè)種群作為一個(gè)整體進(jìn)行分析時(shí),mtDNA CO I和mtDNA CO II基因的Taijimas D檢驗(yàn)結(jié)果達(dá)顯著性負(fù)值(P<0.05),而Fus Fs檢驗(yàn)結(jié)果為非顯著性負(fù)值,二者結(jié)果相反,同時(shí),各種群的Taijimas D和Fus Fs檢驗(yàn)結(jié)果均不顯著地偏離零。11個(gè)茶棍薊馬種群總體的錯(cuò)配分布曲線呈現(xiàn)多峰(圖2),表明種群較穩(wěn)定,種群近期歷史上未經(jīng)歷明顯的擴(kuò)張,與Fus Fs檢驗(yàn)一致。
3 討論
本研究獲得貴州省11個(gè)茶棍薊馬地理種群的137條mtDNA CO I基因序列和132條mtDNA CO II基因序列,2個(gè)基因的堿基組成均有明顯的A/T偏好性,具有典型的昆蟲mtDNA基因堿基組成特點(diǎn)(Jermiin and Crozier,1994;Simon et al.,1994)。另外,Simon等(1994)認(rèn)為,親緣關(guān)系較近的分類單元核苷酸堿基轉(zhuǎn)換大于顛換,而親緣關(guān)系遠(yuǎn)的分類單元?jiǎng)t相反。本研究中,mtDNA CO I和mtDNA CO II基因序列的轉(zhuǎn)換/顛換偏倚率R分別為18.741和5.797,與Simon等(1994)的觀點(diǎn)一致。Knight和Mindell(1993)認(rèn)為,轉(zhuǎn)換/顛換偏倚率R的臨界值為2,當(dāng)R<2時(shí),可認(rèn)為基因突變達(dá)飽和狀態(tài);反之,當(dāng)R>2時(shí)表示基因突變尚未達(dá)飽和狀態(tài)。本研究中,貴州省11個(gè)茶棍薊馬地理種群所有mtDNA CO I和mtDNA CO II基因序列的轉(zhuǎn)換/顛換偏倚率R均大于2,表明貴州省茶棍薊馬mtDNA CO I和mtDNA CO II基因序列的堿基突變尚未達(dá)飽和狀態(tài)。
Hd和π主要用于衡量一個(gè)種群線粒體基因的遺傳多樣性(張桂芬等,2014)。本研究對(duì)貴州省11個(gè)茶棍薊馬地理種群基于mtDNA CO I和mtDNA CO II基因的研究結(jié)果顯示,mtDNA CO I基因序列定義11種單倍型(3個(gè)共享單倍型和8個(gè)獨(dú)享單倍型);mtDNA CO II基因序列定義9種單倍型(4個(gè)獨(dú)享單倍型和5個(gè)共享單倍型),說明貴州省各地理種群茶棍薊馬的mtDNA既存在一定的交流,又有一定程度的遺傳分化。基于mtDNA CO I基因和mtDNA CO II基因進(jìn)一步對(duì)總?cè)后w的FST和Nm進(jìn)行綜合分析顯示,總?cè)后w的FST均在0.05~0.15范圍內(nèi),Nm均大于1,說明貴州省不同地理區(qū)域的茶棍薊馬種群間基因交流水平較高,種群間遺傳分化較小(Balloux and Lugon-Moulin,2002)。另一方面,mtDNA CO I基因和mtDNACO II基因反映各地理種群間的遺傳分化水平并不完全一致,Hua等(2008)研究認(rèn)為不同線粒體基因和蛋白編碼基因不同位點(diǎn)的進(jìn)化速率不同,因此,可推測(cè)mtDNA CO I基因和mtDNACO II基因反映各地理種群間的遺傳分化水平不一致是由基因突變的速率存在差異所造成。
Taijimas D(Tajima,1989)和Fus Fs(Fu,1997)檢驗(yàn)是目前應(yīng)用最廣泛的兩種中性檢驗(yàn)?zāi)P停捎眠@2種模型來推算種群的歷史動(dòng)態(tài)變化(Nielsen,2005)。當(dāng)中性檢驗(yàn)結(jié)果為顯著負(fù)值時(shí),表明種群在歷史上曾發(fā)生過規(guī)模擴(kuò)張和定向選擇;當(dāng)中性檢驗(yàn)結(jié)果為顯著正值時(shí),表明種群在歷史上曾發(fā)生過萎縮和平衡選擇;當(dāng)中性檢驗(yàn)結(jié)果不顯著地偏離零時(shí),中性零假說也不能被排除,表明種群在歷史上保持相對(duì)穩(wěn)定(Tajima,1989;Fu,1997)。從mtDNA CO I和mtDNACO II基因的中性進(jìn)化模型檢驗(yàn)結(jié)果可看出,各種群的Taijimas D和Fus Fs檢驗(yàn)結(jié)果均未達(dá)顯著差異水平;但將11個(gè)茶棍薊馬種群作為一個(gè)整體進(jìn)行分析時(shí),2個(gè)基因的Taijimas D檢驗(yàn)結(jié)果達(dá)顯著水平(P<0.05),而Fus Fs檢驗(yàn)結(jié)果未達(dá)顯著水平,結(jié)果正好相反。有研究表明,F(xiàn)us Fs比Taijimas D能更好地檢驗(yàn)種群的擴(kuò)張現(xiàn)象(Ramosonsins and Rozas,2002)。進(jìn)一步進(jìn)行錯(cuò)配分布分析發(fā)現(xiàn),貴州省11個(gè)茶棍薊馬種群總的錯(cuò)配分布曲線為多峰;Harpending等(1998)認(rèn)為最近形成的或發(fā)生了擴(kuò)張現(xiàn)象的種群,其錯(cuò)配分布曲線呈單峰,相反,存在時(shí)間較長(zhǎng)或相對(duì)穩(wěn)定的種群,其錯(cuò)配分布曲線呈多峰。本研究中,2個(gè)分子標(biāo)記的錯(cuò)配分布曲線為多峰,與Fus Fs檢驗(yàn)結(jié)果一致,表明種群較穩(wěn)定,近期歷史尚未形成明顯擴(kuò)張。Lü等(2016)研究表明貴州省茶棍薊馬經(jīng)歷了明顯的擴(kuò)張,與本研究結(jié)果存在一定的分歧,究其原因:一方面可能是分析樣本數(shù)引起的偏差;另一方面,從2個(gè)基因的錯(cuò)配分布曲線上看,除第一個(gè)峰外,后面的峰形很低,可能存在一種擴(kuò)張與未擴(kuò)張之間的臨界狀態(tài)。
本研究樣本采集地僅限于貴州省部分地區(qū),在遺傳分化與多樣性的分析上可能存在一定偏差。另外,關(guān)于茶棍薊馬目前在多省(區(qū))茶區(qū)為害的情況,應(yīng)對(duì)全國(guó)各省(區(qū))為進(jìn)行深入調(diào)查,如有必要可對(duì)全國(guó)范圍內(nèi)的茶棍薊馬遺傳多樣性進(jìn)行系統(tǒng)研究,及時(shí)掌握種群動(dòng)態(tài),以防止該蟲在全國(guó)各大茶區(qū)擴(kuò)張和暴發(fā)。
4 結(jié)論
貴州省茶棍薊馬總體遺傳多樣性較高,種群間發(fā)生了一定分化,雖然尚未形成明顯的種群擴(kuò)張,但可能已處于擴(kuò)張與未擴(kuò)張之間的臨界狀態(tài)。因此,在農(nóng)業(yè)生產(chǎn)中應(yīng)重視對(duì)茶棍薊馬的防控工作,積極采取綜合防控措施,力爭(zhēng)將茶棍薊馬種群數(shù)量控制在危害水平以下,以保證貴州省茶產(chǎn)業(yè)生產(chǎn)健康可持續(xù)發(fā)展。
參考文獻(xiàn):
董照鋒,張小平. 2018. 5種殺蟲劑對(duì)茶棍薊馬、茶黃薊馬田間防效[J]. 東北農(nóng)業(yè)科學(xué),43(5):38-40. [Dong Z F,Zhang X P. 2018. Field control effect of 5 pesticides on Dendrothrips minowai Priesner and Scirtothrips dorsalis Hood[J]. Journal of Northeast Agricultural Sciences,43(5):38-40.]
李慧玲,張輝,王定鋒,李良德,鄧福新,王慶森. 2014. 不同懸掛高度數(shù)字化色板對(duì)茶棍薊馬的誘集效果試驗(yàn)[J]. 茶葉科學(xué)技術(shù),(4):45-46. [Li H L,Zhang H,Wang D F,Li L D,Deng F X,Wang Q S. 2014. Effects of different suspending heights of digital color boards on trapping Dendrothrips minowai Priesner[J]. Tea Science and Technology,(4):45-46.]
李貌,胡昌雄,吳道慧,陳國(guó)華,陳志星,殷紅慧,徐天養(yǎng),張曉明. 2020. 非施藥植物上薊馬種群動(dòng)態(tài)及優(yōu)勢(shì)種的生態(tài)位分析[J]. 河南農(nóng)業(yè)大學(xué)學(xué)報(bào),54(3):430-438. [Li M,Hu C X,Wu D H,Chen G H,Chen Z X,Yin H H,Xu T Y,Zhang X M. 2020. Population dynamics and niche analysis of dominant species of thrips in non-medicated plants[J]. Journal of Henan Agricultural University,54(3):430-438. ]
呂召云,郅軍銳,周玉鋒,孟澤洪,張駿,楊廣明. 2015. 茶棍薊馬(Dendrothrips minowai Priesner)觸角感器的掃描電鏡觀察[J]. 茶葉科學(xué),35(2):185-195. [Lü Z Y,Zhi J R,Zhou Y F,Meng Z H,Zhang J,Yang G M. 2015. Scanning electron microscopic observations of antennal sensilla of tea stick thrips(Dendrothrips minowai Priesner)[J]. Journal of Tea Science,35(2): 185-195.]
彭萍,王小琴,李品武. 2013. 茶樹病蟲害測(cè)報(bào)與防治技術(shù)[M]. 北京: 中國(guó)農(nóng)業(yè)出版社: 167. [Peng P,Wang X Q,Li P W. 2013. Pests forecast and control technology of tea plant[J]. Beijing:China Agriculture Press: 167.]
喬瑋娜. 2012. DNA條形碼技術(shù)在我國(guó)常見薊馬種類識(shí)別及西花薊馬傳播擴(kuò)散趨勢(shì)分析中的應(yīng)用[D]. 北京: 中國(guó)農(nóng)業(yè)科學(xué)院. [Qiao W N. 2012. Application of DNA barcoding technology for species identification of common trips(Insecta:Thysanoptera) and spread trending analysis of Franklinella occodentialis in China[D]. Beijing: Chinese Academy of Agriculture Sciences.]
沈登榮,宋文菲,袁盛勇,田學(xué)軍,和紹禹,張宏瑞. 2014. 薊馬科部分種類mtDNA-CO I序列變異及系統(tǒng)發(fā)育分析[J]. 應(yīng)用昆蟲學(xué)報(bào),51(1):139-145. [Shen D R,Song W F,Yuan S Y,Tian X J,He S Y,Zhang H R. 2014. Phylogenetic analysis of some Thripidae species based on sequence variation in the mtDNA-CO I segment[J]. Chinese Journal of Applied Entomology,51(1): 139-145. ]
唐美君,肖強(qiáng). 2018. 茶樹病蟲及天敵圖譜[M]. 北京: 中國(guó)農(nóng)業(yè)出版社:136.[Tang M J,Xiao Q. 2018. Colored pitiorial handbook of tea plant pests and natural enemies[J]. Beijing:China Agriculture Industry Press: 136.]
涂娟,金玲莉,謝楓,胡擁軍,樂美旺,劉艷南. 2016. 不同黃、藍(lán)板對(duì)茶園昆蟲的誘捕效果[J]. 蠶桑茶葉通訊,(1):32-34. [Tu J,Jin L L,Xie F,Hu Y J,Yue M W,Liu Y N. 2016. Effects of yellow and blue trap boards on pests of tea garden[J]. Newsletter of Sericulture and Tea,(1):32-34.]
王洪濤,姜法祥,陳敏,王麗麗,王英姿. 2018. 不同顏色粘蟲板對(duì)茶園主要害蟲的誘集效果[J]. 中國(guó)植保導(dǎo)刊,38(6): 55-58. [Wang H T,Jiang F X,Chen M,Wang L L,Wang Y Z. 2018. Effects of different colors trap boards on main pests of tea garden[J]. China Plant Protection,38(6): 55-58.]
武曉云,程曉非,張仲凱. 2009. 西花薊馬(Frankliniella occidentalis) rDNA ITS2和CO I基因5'末端序列的克隆與比較分析[J]. 浙江大學(xué)學(xué)報(bào)(農(nóng)業(yè)與生命科學(xué)版),35(4):355-364. [Wu X Y,Cheng X F,Zhang Z K. 2009. Clo-ning and comparative analysis of rDNA ITS2 and 5' terminal sequence of CO I gene of Frankliniella occidentalis[J]. Journal of Zhejiang University(Agriculture and Life Sciences),35(4):355-364.]
曾明森. 2019. 茶棍薊馬泡沫法施藥防治試驗(yàn)[J].茶葉學(xué)報(bào),60(2):75-79.[Zeng M S. 2019. Control of Dendrothrips Minowai Priesne with aqueou foam covering[J]. Acta Tea Sinica,60(2):75-79.]
張桂芬,喬瑋娜,古君伶,閔亮,萬方浩. 2014. 我國(guó)西花薊馬線粒體DNA-CO I基因變異及群體遺傳結(jié)構(gòu)分析[J]. 生物安全學(xué)報(bào),23(3):196-209. [Zhang G F,Qiao W N,Gu J L,Min L,Wan F H. 2014. Genetic variability of mtDNA-CO I and population structure of Frankliniella occidentalis(Pergande) in China[J]. Journal of Biosafety,23(3):196-209.]
張莉,楊春,李帥,孟澤洪,趙興麗,羅林麗,周玉鋒. 2019. 貴州中部地區(qū)茶園茶棍薊馬的空間分布[J]. 貴州農(nóng)業(yè)科學(xué),47(12):52-54. [Zhang L,Yang C,Li S,Meng Z H,Zhao X L,Luo L L,Zhou Y F. 2019. Spatial distribution pattern of Dendrothrips minowai in tea garden in central Guizhou[J]. Guizhou Agricultural Sciences,47(12):52-54.]
張利娟,沈登榮,張宏瑞,張宏偉,李正躍. 2011. 薊馬基因組DNA提取方法的改進(jìn)[J]. 應(yīng)用昆蟲學(xué)報(bào),48(3):775-781. [Zhang L J,Shen D R,Zhang H R,Zhang H W,Li Z Y. 2011. Method improvement for extraction genomic DNA from thrips[J]. Chinese Journal of Applied Entomology,48(3):775-781.]
趙志清,陳流光. 1998. 茶棍薊馬的發(fā)生規(guī)律與防治技術(shù)[J]. 中國(guó)茶葉,(5):6-7. [Zhao Z Q,Chen L G. 1998. Deve-lopment regularity and control technique of Dendrothrips minowai[J]. China Tea,(5):6-7.]
Balloux F,Lugon-Moulin N. 2002. The estimation of population differentiation with microsatellite markers[J]. Mole-cular Ecology,11(2):155-165.
Bandelt H J,F(xiàn)orster P,R?hl A. 1999. Median-joining networks for inferring intraspecific phylogenies[J]. Molecular Biology and Evolution,16(1):37-48.
Brunner P C,F(xiàn)rey J E. 2010. Habitat-specific population structure in native western flower thrips Frankliniella occidentalis(Insecta,Thysanoptera)[J]. Journal of Evolutionary Biology,23(4):797-804.
Dickey A M,Vivek K,Hoddle M S,F(xiàn)underburk J E,Kent M J,Antonella J C,Shatters R G,Osborne L S,Mckenzie C L,Mans B J. 2015. The Scirtothrips dorsalis species complex:Endemism and invasion in a global pest[J]. PLoS One 10(4):e0123747.
Dupanloup I,Schneider S,Excoffier L. 2002. A simulated annealing approach to define the genetic structure of populations [J]. Molecular Ecology,11(12):2571-2581.
Excoffier L,Lischer H E L. 2010. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows[J]. Molecular Ecology Resources,10(3):564-567.
Folmer O,Black M,Hoeh W,Lutz R,Vrijenhoek R. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates[J]. Molecular Marine Biology and Biotechnology,3(5):294-299.
Fu X Y. 1997. Statistical tests of neutrality of mutations against population growth,hitchhiking and background selection[J]. Genetics,147(2):915-925.
Harpending H C,Batzer M A,Gurven M,Jorde L B,Rogers A R,Sherry S T. 1998. Genetic traces of ancient demography[J]. Proceedings of the National Academy of Scien-ces of the United States of America,95(4):1961-1967.
Hua J M,Li M,Dong P Z,Cui Y,Xie Q,Bu W J. 2008. Comparative and phylogenomic studies on the mitochondrial genomes of Pentatomomorpha(Insecta: Hemiptera: Heteroptera)[J]. BMC Genomics,9(1):610.
Jermiin L S,Crozier R H. 1994. The cytochrome b region in the mitochondrial DNA of the ant Tetraponera rufoniger:Sequence divergence in Hymenoptera may be associated with nucleotide content[J]. Journal of Molecular Evolution,38(3):282-294.
Knight A,Mindell D P. 1993. Substitution bias,weighting of DNA sequence evolution,and the phylogenetic position of Fea s viper[J]. Systematic Biology,42(1):18-31.
Librado P,Rozas J. 2009. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data[J]. Bioinformatics(Oxford,England),25(11):1451-1452.
Lü Z Y,Zhi J R,Zhou Y F,Meng Z H,Yue W B. 2016. Genetic diversity and origin of Dendrothrips minowai (Thysanoptera: Thripidae) in Guizhou,China[J]. Journal of Asia-Pacific Entomology,19(4):1035-1043.
Nielsen R. 2005. Molecular signatures of natural selection [J]. Annual Review of Genetics,39(1):197-218.
Peterson M A,Dobler S,Holland J,Tantalo L,Locke S. 2001. Behavioral,molecular,and morphological evidence for a hybrid zone between Chrysochus auratus and C. cobaltinus(Coleoptera: Chrysomelidae)[J]. Annals of the Entomological Society of America,94(1):1-9.
Ramosonsins S E,Rozas J. 2002. Statistical properties of new neutrality tests against population growth[J]. Molecular Biology and Evolution,19(12):2092-2100.
Simon C,F(xiàn)rati F,Beckenbach A,Crespi B,Liu H,F(xiàn)look P. 1994. Evolution,weighting,and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers[J]. Annals of the Entomological Society of America,87(6):651-701.
Tajima F. 1989. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism[J]. Genetics,123(3): 585-595.
Tamura K,Stecher G,Peterson D,F(xiàn)ilipski A,Kumar S. 2013. MEGA 6:Molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution,30(12):2725-2729.
Yang X M,Sun J T,Xue X F,Li J B,Hong X Y. 2012a. Invasion genetics of the western flower thrips in China: Evidence for genetic bottleneck,hybridization and bridgehead effect[J]. PLoS One,4(7):e34567.
Yang X M,Sun J T,Xue X F,Zhu W C,Hong X Y. 2012b. Development and characterization of 18 novel EST-SSRs from the western flower thrips,F(xiàn)rankliniella occidentalis (Pergande)[J]. International Journal of Molecular Scien-ces,13(3):2863-2876.
(責(zé)任編輯 麻小燕)
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
