干旱脅迫下腐植酸肥料對(duì)燕麥光響應(yīng)曲線的影響
2020-11-16 07:19:12李英浩劉景輝趙寶平米俊珍戴云仙
關(guān)鍵詞:模型
李英浩 劉景輝 趙寶平 田 露 米俊珍 王 琦 戴云仙
(內(nèi)蒙古農(nóng)業(yè)大學(xué) 雜糧產(chǎn)業(yè)協(xié)同創(chuàng)新中心,呼和浩特 010019)
全球干旱、半干旱地區(qū)面積約占總耕地面積的40%以上,是影響植物生存、生長(zhǎng)和分布最重要的逆境脅迫因素之一,由于近年來全球性氣候惡化使得干旱發(fā)生周期越來越短,干旱程度越來越重,已經(jīng)嚴(yán)重影響大部分作物正常的生長(zhǎng)發(fā)育,對(duì)糧食生產(chǎn)構(gòu)成的威脅也越來越大,產(chǎn)量大大降低[1]。提高作物抗旱性以及高效利用水資源,已成為我國(guó)旱區(qū)促進(jìn)農(nóng)業(yè)可持續(xù)發(fā)展的必然選擇[2]。水分是植物生長(zhǎng)過程中不可或缺的環(huán)境因子,參與細(xì)胞內(nèi)的多種代謝活動(dòng),水分脅迫能夠造成植物生長(zhǎng)與發(fā)育發(fā)生顯著變化[3-4]。因此,研究植物的耐旱性是植物生理生態(tài)學(xué)的重點(diǎn)問題[5]。光合作用是植物最基本的生命活動(dòng),較高的光合碳同化能力是獲得高產(chǎn)的前提[6],提高葉片的光合性能是實(shí)現(xiàn)作物高產(chǎn)的重要途徑[7]。植物的光響應(yīng)曲線反映的是光合速率隨著光合有效輻射變化特性,是判定植物光合能力的重要指標(biāo)[8],光響應(yīng)曲線的模擬可得出多個(gè)基礎(chǔ)生理參數(shù)(包括表觀量子效率、最大凈光合速率、光補(bǔ)償點(diǎn)、光飽和點(diǎn)以及暗呼吸速率)[9-12],這些參數(shù)能直接或間接反映出植物生理生態(tài)學(xué)過程。目前,國(guó)內(nèi)外多采用直角雙曲線、非直角雙曲線模型和指數(shù)模型對(duì)植物的光響應(yīng)過程進(jìn)行擬合[9],但在使用這 3 種光響應(yīng)模型時(shí),求得的光飽和點(diǎn)低于實(shí)測(cè)值,甚至無光飽和點(diǎn)出現(xiàn),最大凈光合速率大于實(shí)測(cè)值[10-11]。葉子飄等[11]提出直角雙曲線修正模型,國(guó)內(nèi)也有學(xué)者利用該模型擬合光響應(yīng)曲線[12-15],但此模型在研究禾本科作物光合性能方面應(yīng)用較少。
燕麥?zhǔn)羌Z飼兼用作物,喜冷涼、耐低溫且抗逆性強(qiáng),在中國(guó)西北干旱地區(qū)廣泛種植,這些地區(qū)常常遭受到干旱的侵襲,是導(dǎo)致燕麥減產(chǎn)的主要原因之一[16]。腐植酸是動(dòng)植物遺骸經(jīng)過微生物分解和轉(zhuǎn)化等一系列過程形成的一類有機(jī)物質(zhì)[17]。李英浩等[16]研究發(fā)現(xiàn),在干旱脅迫條件下噴施腐植酸可以有效調(diào)節(jié)燕麥的各項(xiàng)光合生理參數(shù),改善其光合性能,進(jìn)而提高燕麥產(chǎn)量。但上述研究只集中在單一光強(qiáng)條件下,僅僅針對(duì)腐植酸在干旱脅迫下對(duì)燕麥葉片光合參數(shù)的瞬時(shí)值影響做研制。關(guān)于作物光合參數(shù)隨光合有效輻射梯度的動(dòng)態(tài)變化研究尚未見報(bào)道。因此,本試驗(yàn)利用直角雙曲線模型、非直角雙曲線模型、指數(shù)模型以及直角雙曲線修正模型對(duì)不同干旱脅迫處理的燕麥噴施腐植酸后的光響應(yīng)曲線進(jìn)行擬合,比較4種模型的差異,從中選出最優(yōu)的光合響應(yīng)模型,旨在分析干旱脅迫下腐植酸對(duì)燕麥光合作用的影響,以期為燕麥生產(chǎn)中應(yīng)用腐植酸肥料提高抗旱性提供科學(xué)依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料及試驗(yàn)設(shè)計(jì)
試驗(yàn)于2018年5—9月在內(nèi)蒙古自治區(qū)呼和浩特市內(nèi)蒙古農(nóng)業(yè)大學(xué)燕麥產(chǎn)業(yè)研究中心溫室進(jìn)行。供試燕麥品種為‘燕科二號(hào)’,由內(nèi)蒙古自治區(qū)呼和浩特市農(nóng)牧業(yè)科學(xué)研究院提供。腐植酸水溶肥料由內(nèi)蒙古自治區(qū)永業(yè)農(nóng)豐生物技術(shù)有限責(zé)任公司提供,其水溶腐植酸含量≥50 g/L,(N+P+K)≥200 g/L,微量元素(錳、硼、鉬、鋅等)≥10 g/L。采用盆栽方法,土壤為蛭石與泥炭土按質(zhì)量比為1∶1的比例混合而成。泥炭土中(N+P+K)>10 g/kg,有機(jī)質(zhì)含量>50 g/kg,pH 5.5~6.5,土壤含水量為10%。塑料盆高25 cm,直徑20 cm,混合土2.5 kg/盆,播種前底施磷酸二銨2 g/盆(每千克土施氮0.14 g,施磷0.37 g),在拔節(jié)初期(6月1日)追施固態(tài)尿素2 g/盆(每千克土施氮0.37 g),隨灌水逐漸溶解。
試驗(yàn)采用隨機(jī)區(qū)組排列,設(shè)置重度干旱脅迫(45%田間持水量),中度干旱脅迫處理(60%田間持水量)和正常供水處理(75%田間持水量) 3個(gè)水分條件,不同水分條件下設(shè)腐植酸肥料(HA)和等量清水(CK)2個(gè)處理,共6個(gè)處理,每個(gè)處理重復(fù)3次,每個(gè)重復(fù)種植3盆,共54盆。5月1日播種,5月7日出苗,水分脅迫在拔節(jié)期(6月1日)開始,于抽穗期(7月1日)噴施腐植酸,噴施量為0.2 mL/盆,噴施前稀釋500倍,于9:00進(jìn)行噴施,7月3日進(jìn)行光響應(yīng)曲線測(cè)定。每天17:00利用稱重法進(jìn)行補(bǔ)水以保持各處理土壤含水量穩(wěn)定,土壤田間持水量、永久萎蔫點(diǎn)及土壤質(zhì)量含水量按Ryan等[18]的方法分析測(cè)定。
1.2 光響應(yīng)曲線的測(cè)定
采用(CIRAS-3)便攜式光合測(cè)定系統(tǒng)對(duì)抽穗期燕麥旗葉進(jìn)行測(cè)定,在晴天的8:00—11:00連續(xù)測(cè)定,每處理測(cè)定3株植株,每株選取長(zhǎng)勢(shì)相同的旗葉葉片重復(fù)測(cè)3次,取平均值進(jìn)行分析。測(cè)定時(shí)葉溫控制在 20~25 ℃、相對(duì)濕度(65±5)%左右、CO2濃度(380±2) μmol/mol。測(cè)定時(shí)采用紅藍(lán)光源,設(shè)置(2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、80、50、0) μmol/(m2·s)共 14個(gè)遞減光合有效輻射梯度,采用自動(dòng)程序測(cè)量,每個(gè)設(shè)置值下停留200~150 s。光合測(cè)定儀自動(dòng)記錄凈光合速率(Pn)、氣孔導(dǎo)度(Gs)、胞間CO2濃度(Ci)、蒸騰速率(Tr)等光合參數(shù),最終選取平均數(shù)進(jìn)行分析,PAR為光合有效輻射。
1.3 數(shù)據(jù)分析
根據(jù)得到的不同光合有效輻射的凈光合速率,繪制光合速率的光響應(yīng)(Pn-PAR) 曲線,根據(jù)實(shí)測(cè)數(shù)據(jù)點(diǎn)的走勢(shì)估計(jì)最大凈光合速率(Pnmax)、光飽和點(diǎn)(LSP)、光補(bǔ)償點(diǎn)(LCP)和暗呼吸速率(Rd)等參數(shù),同時(shí)采用傳統(tǒng)弱光下(PAR≤200 μmol/mol)PAR與Pn的線性回歸法,估算出光響應(yīng)參數(shù)作為實(shí)測(cè)值與4種模型擬合值比較分析,分別采用直角雙曲線模型[19]、非直角雙曲線模型[20]、指數(shù)模型[21]和直角雙曲線修正模型[22]對(duì)光響應(yīng)曲線進(jìn)行擬合。
試驗(yàn)數(shù)據(jù)用Excel 2016進(jìn)行整理,用SPSS 19.0進(jìn)行非線性回歸擬合數(shù)據(jù)及相關(guān)性分析,用Origin 9.1系統(tǒng)繪制圖形,用SAS 9.0進(jìn)行方差分析和多重比較分析。
2 結(jié)果與分析
2.1 燕麥凈光合速率的光響應(yīng)變化趨勢(shì)
由圖1可知,PAR≤1 400 μmol/(m2·s)時(shí),在不同的干旱脅迫處理下,燕麥Pn的變化趨勢(shì)一致,均隨著PAR的提高逐漸上升。當(dāng)PAR>1 400 μmol/(m2·s)時(shí),正常供水和中度干旱脅迫處理下,Pn開始略微減少并逐漸趨于平穩(wěn),即達(dá)到光飽和點(diǎn),而重度干旱脅迫處理下Pn達(dá)到飽和點(diǎn)后有顯著下降的趨勢(shì),且隨著PAR增大,光抑制現(xiàn)象顯著。在相同PAR水平下,Pn隨土壤含水量的減少而降低,噴施HA相比于CK,正常供水時(shí)Pn值無明顯變化,中度干旱脅迫下Pn值略有提升,重度干旱脅迫下Pn值顯著提高,且Pn隨PAR的下降幅度減小。
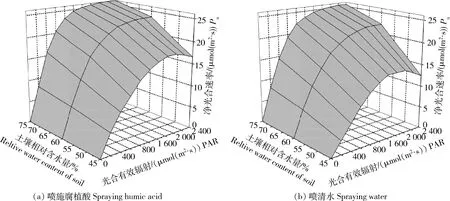
圖1 不同處理下燕麥凈光合速率對(duì)光合有效輻射的響應(yīng)Fig.1 Response of net photosynthetic rate of oats to photosynthetically active radiation under different drought stress
2.2 燕麥光響應(yīng)特征參數(shù)實(shí)測(cè)值方差分析
由表1可知,隨著干旱脅迫程度加劇,燕麥葉片的光飽和點(diǎn)(LSP)和光補(bǔ)償點(diǎn)(LCP)逐漸增大,而最大凈光合速率(Pnmax)和暗呼吸速率(Rd)逐漸減小,重度干旱脅迫與正常供水相比達(dá)到顯著差異(P<0.05)。正常供水條件下噴施HA與CK相比,LSP、LCP、Pnmax和Rd值均無顯著差異;中度干旱脅迫下噴施HA相比于CK,只有LCP和Pnmax達(dá)到顯著差異;而在重度干旱脅迫條件下噴施HA后,LCP值降低31.09%,Pnmax值提高15.21%,Rd值提高62.5%,且相比于CK差異均達(dá)到顯著水平(P<0.05)。綜上說明,噴施HA可以在重度干旱脅迫條件下有效提高燕麥葉片的光能利用率,使其葉片在不同光合有效輻射下表現(xiàn)出較好的光合性能。
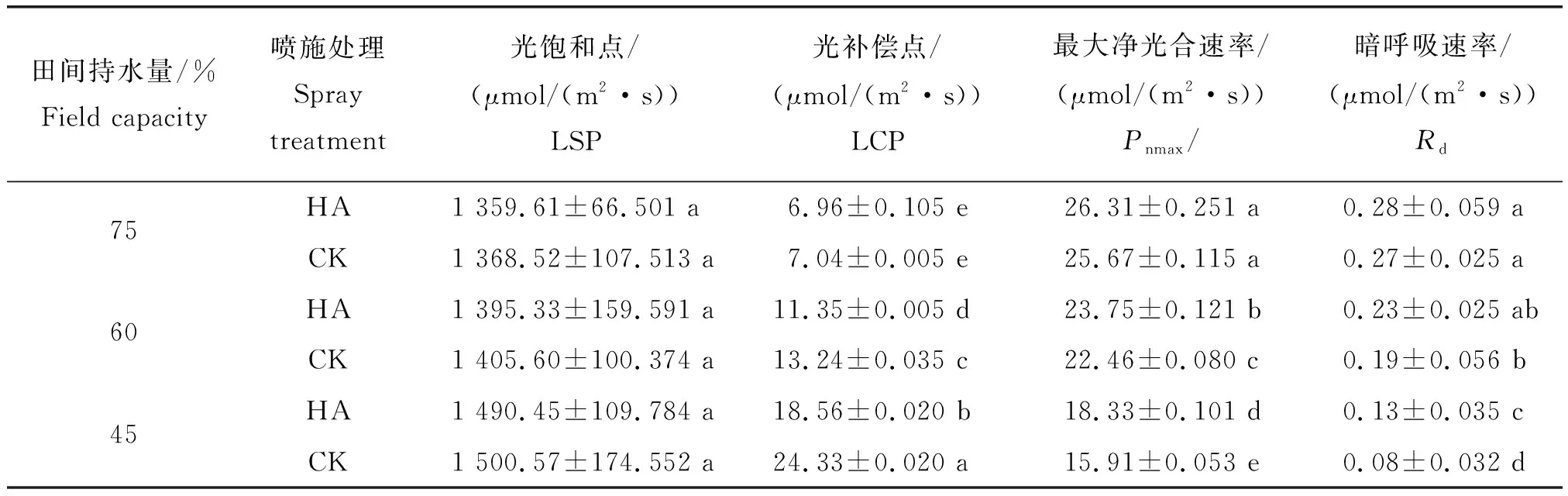
表1 不同干旱處理(田間持水量)下燕麥的光響應(yīng)特征參數(shù)實(shí)測(cè)值方差分析Table 1 Analysis of variance of measured values of light response characteristic parameters of oats under different drought treatment (field capacity)
2.3 燕麥光響應(yīng)曲線模型擬合效果對(duì)比分析
由圖2,圖3,圖4和圖5可知, 4種光響應(yīng)模型的擬合效果差異顯著,其中只有直角雙曲線修正模型在較高PAR下能夠很好的擬合Pn下降趨勢(shì),并且可以很好地?cái)M合出實(shí)測(cè)曲線中的凈光合速率穩(wěn)定狀態(tài)。而其他3種模型在較高PAR時(shí)的擬合點(diǎn)都要比實(shí)測(cè)值高,這是因?yàn)橹苯请p曲線、非直角雙曲線和指數(shù)模型擬合的光響應(yīng)曲線在較高PAR擬合點(diǎn)時(shí)都是一條沒有極值的漸近線,因此不能很好的擬合各處理在飽和光強(qiáng)后的Pn變化過程。分析比較4種光響應(yīng)曲線的宏觀擬合情況發(fā)現(xiàn),直角雙曲線模型的擬合效果最差,只有直角雙曲線修正模型在PAR>1 400 μmol/(m2·s)時(shí)幾乎與實(shí)測(cè)曲線完全擬合,這是其他3種模型不能做到的,因此擬合效果最佳。

圖2 直角雙曲線模型擬合分析燕麥光響應(yīng)曲線Fig.2 Analysis of light response curves of oats by rectangular hyperbola model

圖3 非直角雙曲線模型擬合分析燕麥光響應(yīng)曲線Fig.3 Analysis of light response curves of oats by non-rectangular hyperbola model

圖4 指數(shù)模型擬合分析燕麥光響應(yīng)曲線Fig.4 Analysis of light response curves of oats by exponential model

圖5 直角雙曲線修正模型擬合分析燕麥光響應(yīng)曲線Fig.5 Analysis of light response curves of oats by modefied rectangular hyperbola model
2.4 燕麥光響應(yīng)特征參數(shù)在不同擬合模型下的對(duì)比分析
由表2可知, 4種模型均能較好地?cái)M合燕麥葉片光響應(yīng)過程,但與直角雙曲線模型相比,指數(shù)模型、非直角模型和直角雙曲線修正模型的擬合效果更好,且只有這3種模型的R2>0.99,然而R2值大并不能說明燕麥葉片在不同處理下光合作用的實(shí)際情況,只能宏觀地表現(xiàn)出擬合的曲線是否相近實(shí)測(cè)曲線。綜合比較只有直角雙曲線修正模型擬合度最高,其R2=0.999,并且擬合的參數(shù)LSP、LCP、Pnmax及Rd值與實(shí)測(cè)值最為接近,其余3種模型擬合的結(jié)果中均無光飽和點(diǎn)LSP,并且LCP、Pnmax和Rd值也都高于實(shí)測(cè)值。因此,綜合比較 4 種模型對(duì)燕麥葉片光響應(yīng)參數(shù)的擬合度(R2值),結(jié)果為直角雙曲線修正模型>指數(shù)模型>非直角雙曲線模型>直角雙曲線模型。由此可見,直角雙曲線修正模型更適用不同處理下燕麥葉片光響應(yīng)過程與特征參數(shù)的擬合。
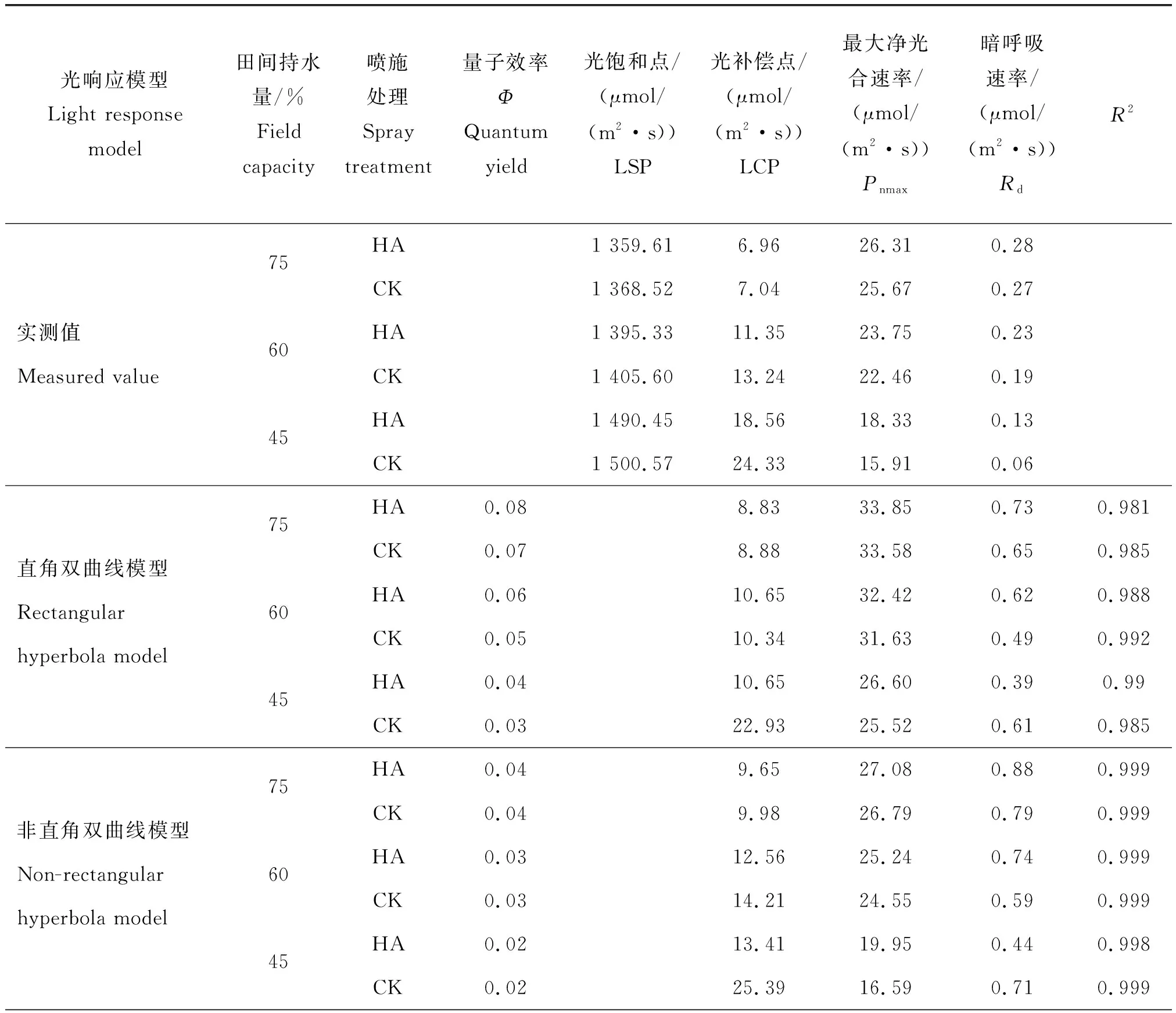
表2 不同干旱處理(田間持水量)下燕麥的光響應(yīng)特征參數(shù)模擬擬合值Table 2 simulated fitted values of light response characteristic parameters of oats under different drought treatments (field capacity)

表2(續(xù))
2.5 燕麥光響應(yīng)曲線擬合
由圖6可知,Tr和Gs均隨PAR的增加呈上升趨勢(shì),而Ci隨PAR的增加呈下降趨勢(shì),且三者在PAR增加到1 400 μmol/(m2·s)時(shí)達(dá)到穩(wěn)定狀態(tài),即飽和點(diǎn);相同PAR水平下,Tr、Gs和Ci均隨干旱脅迫程度的加深而降低,正常供水條件下噴施HA,三者的變化與CK趨勢(shì)相同,甚至曲線的某個(gè)階段出現(xiàn)重合,中度干旱脅迫下噴施HA與CK相比,趨勢(shì)變化略有不同;而重度干旱脅迫條件下噴施HA使三者的響應(yīng)曲線變化趨勢(shì)發(fā)生明顯改變,Tr和Gs的提升速率加快,Ci的下降速率明顯減緩。綜上可知,干旱脅迫導(dǎo)致燕麥葉片氣孔導(dǎo)度、蒸騰作用及胞間二氧化碳濃度降低,噴施HA能有效緩解干旱脅迫造成的影響,尤其在重度干旱脅迫下效果最佳。
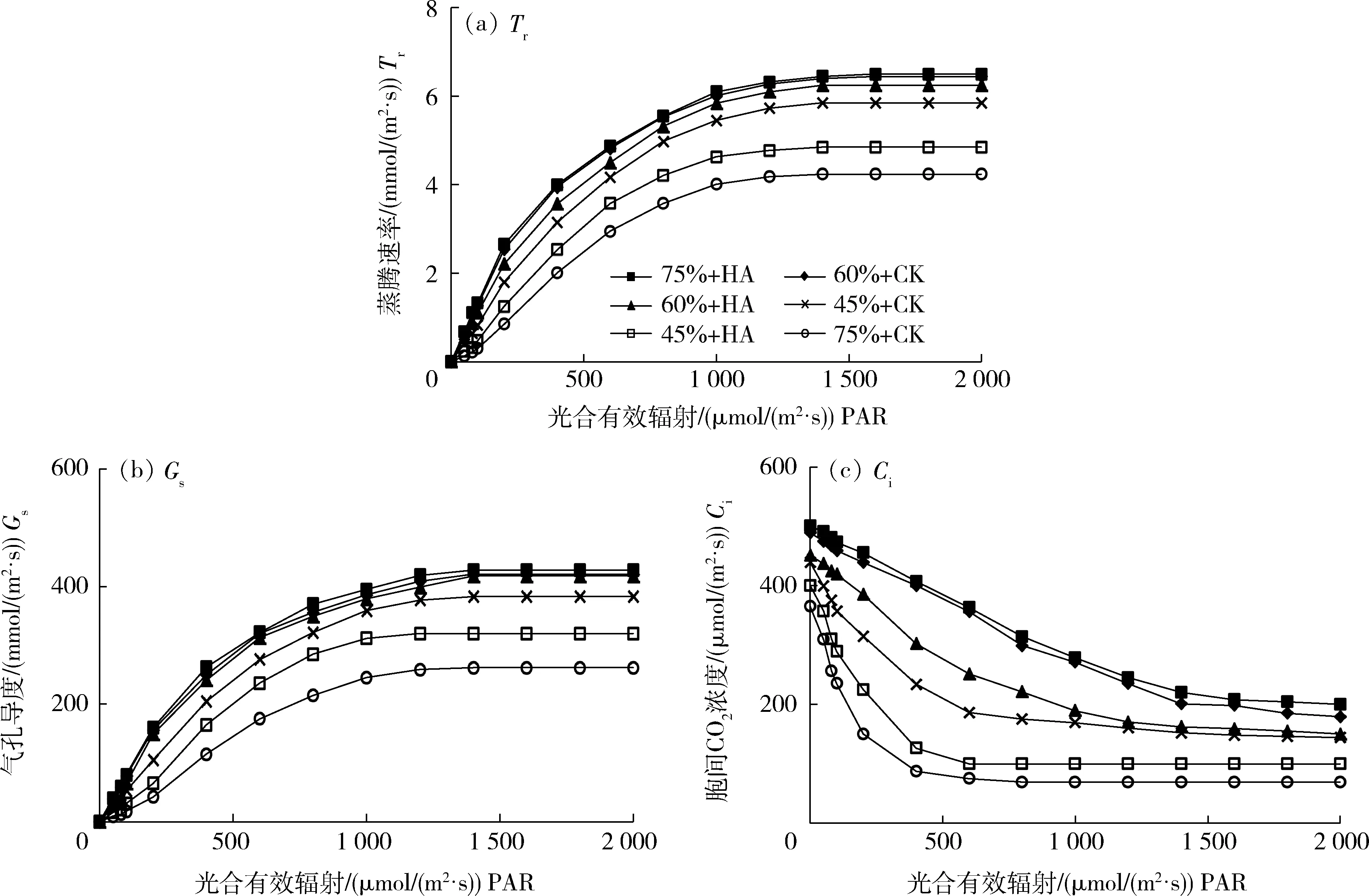
圖6 不同處理下燕麥的光合參數(shù)響應(yīng)曲線Fig.6 Response curves of photosynthetic parameters of oats under different treatments
2.6 不同處理對(duì)燕麥產(chǎn)量的影響
由圖7可知,不同處理之間燕麥的籽粒產(chǎn)量和生物產(chǎn)量存在顯著差異。干旱脅迫導(dǎo)致其籽粒產(chǎn)量和生物產(chǎn)量逐漸降低,與正常供水相比,重度干旱脅迫下籽粒產(chǎn)量和生物產(chǎn)量均顯著下降(P<0.05),中度干旱脅迫下其籽粒產(chǎn)量降低比例未達(dá)到顯著水平;正常供水和中度干旱脅迫條件下噴施HA與CK相比無顯著差異,而重度干旱脅迫條件下噴施HA后,燕麥籽粒產(chǎn)量和生物產(chǎn)量較CK分別提高8.42%和23.15%,差異均顯著(P<0.05)。綜上說明,腐植酸在干旱脅迫下可以提高燕麥籽粒產(chǎn)量和生物產(chǎn)量,且重度干旱脅迫下腐植酸的增產(chǎn)效果最佳。

圖7 不同處理下燕麥的籽粒產(chǎn)量(a)和生物產(chǎn)量(b)Fig.7 Grain yield (a) and biological yield (b) of oats under different treatments
2.7 相關(guān)性分析
由表3可知,燕麥籽粒產(chǎn)量與葉片光飽和點(diǎn)LSP和光補(bǔ)償點(diǎn)LCP呈極顯著負(fù)相關(guān)關(guān)系,與葉片最大凈光合速率Pnmax和暗呼吸速率Rd呈極顯著正相關(guān)關(guān)系;燕麥生物產(chǎn)量與葉片光飽和點(diǎn)LSP和光補(bǔ)償點(diǎn)LCP呈極顯著負(fù)相關(guān)關(guān)系,與葉片最大凈光合速率Pnmax和暗呼吸速率Rd呈極顯著正相關(guān)關(guān)系。
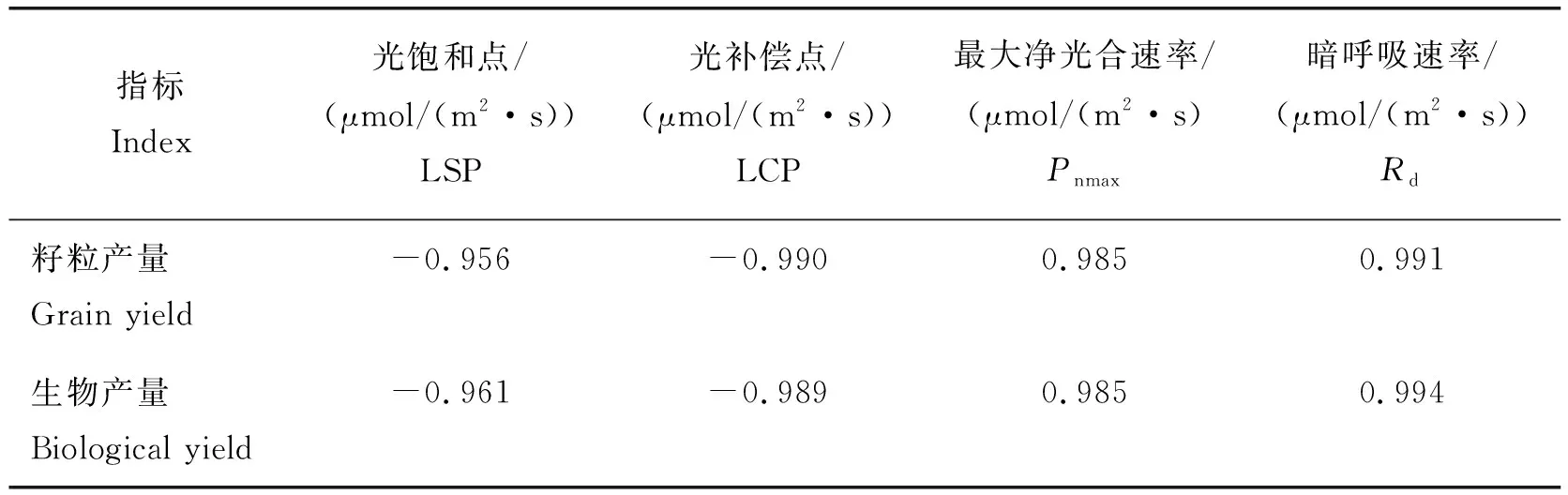
表3 燕麥產(chǎn)量與光響應(yīng)特征參數(shù)相關(guān)性分析Table 3 Correlation analysis of oat yield and light response characteristic parameters
3 討 論
3.1 不同處理下燕麥光響應(yīng)曲線的變化
光照是影響植物生長(zhǎng)的重要環(huán)境因子之一。Pn-PAR曲線反映植物凈光合速率隨光照強(qiáng)度的變化規(guī)律,從曲線中找到光合作用的最大光合速率、表觀量子效率、光飽和點(diǎn)、光補(bǔ)償點(diǎn)和暗呼吸速率等生理參數(shù),有助于判定植物光合機(jī)構(gòu)運(yùn)轉(zhuǎn)狀況、光合作用能力和光合作用效率及其受環(huán)境變化的影響程度,因此Pn-PAR曲線的測(cè)定是植物光合生理生態(tài)學(xué)研究的重要指標(biāo)之一[23-27]。光合作用模型就是描述Pn與PAR的相互關(guān)系,對(duì)探究植物光合作用響應(yīng)機(jī)制和評(píng)價(jià)光合效率具有重要意義[14]。國(guó)內(nèi)外學(xué)者提出了不同的光響應(yīng)模型并進(jìn)行了擬合,由于推導(dǎo)機(jī)理和方式的不同,各個(gè)模型都存在一定的優(yōu)缺點(diǎn)[28]。本試驗(yàn)中擬合的4種模型中非直角雙曲線模型和指數(shù)模型都能很好的擬合PAR<1 400 μmol/(m2·s)的光響應(yīng)過程,但只有直角雙曲線修正模型能夠擬合出高PAR值下Pn的飽和點(diǎn)及下降趨勢(shì),直角雙曲線模型擬合的最差(R2<0.99),產(chǎn)生這樣的原因是因?yàn)橹苯请p曲線模型、非直角雙曲線模型和指數(shù)模型是一條沒有極值的漸進(jìn)線,不能擬合出曲線的歪曲程度[29-30]。有研究表明,在不同的逆境脅迫條件下,擬合光響應(yīng)曲線時(shí)所適用的擬合模型是不同的[28],因此從本試驗(yàn)的結(jié)果中得知,相比于其他3種模型而言,直角雙曲線修正模型更適用于干旱脅迫環(huán)境下燕麥的光響應(yīng)曲線擬合。
李瑞姣等[31]研究表明,輕、中度干旱脅迫下日本莢蒾葉片Pn雖低于對(duì)照但差異逐漸減小,而長(zhǎng)期重度干旱脅迫下Pn均為負(fù)值,進(jìn)一步說明該植物對(duì)輕、中度干旱有一定的耐受性。同樣在本試驗(yàn)中,從光響應(yīng)曲線模型中看出,在PAR<1 400 μmol/(m2·s)時(shí),3個(gè)水分梯度下燕麥的Pn值均隨PAR的增加而增大,正常供水和中度干旱脅迫下Pn達(dá)到光飽和點(diǎn)后趨于穩(wěn)定,而重度干旱脅迫下Pn在光飽和點(diǎn)后有明顯的下降趨勢(shì)。因此推測(cè),燕麥對(duì)中度干旱脅迫有較好的耐受性與適應(yīng)性,而重度干旱脅迫引起植物水分虧缺,使得光合作用受到抑制[32]。
HA在重度干旱脅迫條件下可以顯著提升小麥和馬鈴薯等作物的Pn,而在輕度或中度干旱脅迫下Pn值基本保持不變[33]。本研究結(jié)果中,在正常供水和中度干旱脅迫條件下噴施HA,燕麥葉片Pn隨PAR的變化趨勢(shì)無明顯改變,而在重度干旱脅迫下,Pn的下降幅度明顯減小,說明HA在不同干旱脅迫條件下對(duì)植物葉片Pn的影響不同。同時(shí)本研究發(fā)現(xiàn),與CK相比,HA在較高PAR下對(duì)Pn變化趨勢(shì)的影響較大,而在較低PAR下Pn幾乎保持一致(圖1),具體原因還需進(jìn)一步研究得知。
3.2 不同處理下燕麥光響應(yīng)特征參數(shù)的變化
各項(xiàng)光響應(yīng)特征參數(shù)表明,隨著土壤含水量的降低,植物葉片的光飽和點(diǎn)LSP與光補(bǔ)償點(diǎn)LCP表現(xiàn)出一定程度的升高趨勢(shì),因?yàn)楦珊得{迫導(dǎo)致葉片光合能力受阻,需要一定程度光補(bǔ)償效應(yīng)來提高其光能利用效率[34],這與本研究結(jié)果相吻合。而重度干旱脅迫下噴施HA相比CK,LCP顯著降低(P<0.05),說明燕麥葉片對(duì)重度干旱環(huán)境的適應(yīng)性減弱[35],降低了葉片對(duì)光的利用能力,而HA可以提升葉片的光補(bǔ)償效應(yīng),幫助其適應(yīng)干旱環(huán)境。研究結(jié)果中,噴施HA使葉片的Pnmax有顯著提高(P<0.05),同時(shí)有效緩解重度干旱所引起的光抑制現(xiàn)象。植物Pnmax主要是由電子傳遞速率和Rusbico活性決定的[35],說明在嚴(yán)重干旱條件下HA可有效提高燕麥葉片的電子傳遞速率和Rusbico活性,因此導(dǎo)致其Pnmax增大,這與陸燕元等[36]在甘薯中的研究結(jié)果一致。光合作用的表觀量子效率(Φ)是反映植物對(duì)光能利用效率的重要指標(biāo),傳統(tǒng)的確定方法是利用直線方程擬合弱光強(qiáng)PAR≤200 μmol/(m2·s)的光響應(yīng)數(shù)據(jù)得到的直線斜率表示。已有研究表明,在適宜的生長(zhǎng)條件測(cè)定一般植物的表觀量子效率在0.03~0.05[37]。但是重度干旱脅迫下燕麥葉片的量子效率(Φ)<0.03,小于一般植物的光合量子效率,而HA可有效提升植物葉肉細(xì)胞活躍度,進(jìn)而促進(jìn)細(xì)胞對(duì)光能的利用效率,因此可顯著提高燕麥的光合量子效率(P<0.05)。
3.3 不同處理下燕麥光合參數(shù)響應(yīng)曲線的變化
引起植株葉片Pn下降的原因有氣孔因素和非氣孔因素,氣孔因素指葉片受逆境脅迫導(dǎo)致氣孔導(dǎo)度(Gs)降低,胞間二氧化碳濃度(Ci)不能滿足光合作用的需要,非氣孔因素是由于葉片葉綠體的活性和Rusbico酶活性降低引起的光合作用降低[38]。本研究中,干旱脅迫使燕麥葉片的Gs和Ci降低,說明Pn的下降由氣孔因素引起。而HA在干旱脅迫下對(duì)Gs和Ci都有提升,說明腐植酸可以通過調(diào)節(jié)葉片氣孔運(yùn)動(dòng)來促進(jìn)其光合作用[16]。同時(shí)本研究發(fā)現(xiàn),HA可加快Gs和Tr隨PAR的提升速率,減緩Ci的降低速率,更加印證HA對(duì)葉片氣孔的調(diào)節(jié)作用。
4 結(jié) 論
燕麥葉片中Pn隨PAR增大逐漸升高,當(dāng)PAR接近1 400 μmol/(m2·s)時(shí)趨于平穩(wěn),重度干旱脅迫下Pn在飽和點(diǎn)后出現(xiàn)明顯下降趨勢(shì),噴施HA可以減小Pn的下降幅度;光響應(yīng)特征參數(shù)中,與正常供水相比,重度干旱脅迫導(dǎo)致燕麥葉片的Pnmax和Rd值顯著提升,LSP和LCP顯著降低。噴施HA與CK相比,重度干旱脅迫下Pnmax和Rd值顯著下降,LSP和LCP顯著升高;與CK相比,重度干旱脅迫下噴施HA,燕麥葉片Tr和Gs隨著PAR增大加速升高,Ci緩慢降低;與CK相比,重度干旱脅迫下HA顯著提高燕麥產(chǎn)量。綜上表明燕麥葉片可以適應(yīng)中度干旱脅迫,HA可以緩解重度干旱脅迫對(duì)燕麥葉片光合系統(tǒng)造成的傷害。
猜你喜歡
童話王國(guó)·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網(wǎng)絡(luò)安全與數(shù)據(jù)管理(2022年1期)2022-08-29 03:15:20
導(dǎo)航定位學(xué)報(bào)(2022年4期)2022-08-15 08:27:00
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀(jì)智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:36
成都醫(yī)學(xué)院學(xué)報(bào)(2021年2期)2021-07-19 08:35:14
新世紀(jì)智能(數(shù)學(xué)備考)(2020年9期)2021-01-04 00:25:14
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
