休眠卵孵化對策與食物濃度對萼花臂尾輪蟲(Brachionus calyciflorus)兩品系干雌體種群增長的影響
2020-11-24 09:30:32邢康南牛翠娟
生態學報 2020年21期
邢康南,牛翠娟
生物多樣性與生態工程教育部重點實驗室,北京師范大學生命科學學院, 北京 100875
單巢綱輪蟲生活史為典型的世代交替,休眠卵的形成是其生活史中非常重要的一部分,輪蟲依靠休眠卵來適應不良環境的變化,對延續種群、保留基因有重要意義。因此,與休眠相聯系的有性生殖也被看作是對不良環境的適應[1-3]。一個基因要被保留下來,必須在種群中傳遞下去,而環境中的不利因素對這一過程有阻礙作用。由于休眠期的存在,帶有基因變異的休眠卵的形成和這些具有基因變異后代的出生在時間上分隔開來,分隔時間或長或短。輪蟲休眠卵庫就像“種子貯存庫”一樣,當環境恢復到適宜輪蟲生存時, 它們可以在短時間內孵化出來并依靠孤雌生殖迅速繁衍擴大種群,有利于種群避開不利的生態因子,使該物種的基因在棲息地中保留下來,而不至于滅絕。此外,休眠卵耐干旱的特性是輪蟲在全球范圍內廣泛分布的原因,因為高耐旱能力有助于休眠卵的散布,例如借助風力或鳥類、昆蟲等動物的銜食而散布到更廣泛的范圍以致全球范圍內,這樣可使輪蟲從不適宜生存的環境中解放出來,盡快在適宜生存的環境孵化、增殖[4-5]。
對單巢綱輪蟲休眠卵的早期研究主要集中于休眠卵形成、孵化過程及其影響因素等方面,隨著研究的逐漸深入,休眠卵的孵化時間引起了研究者的關注[6- 8]。有性生殖產生的休眠卵會沉入自然水體底泥中的休眠卵庫進行休眠,休眠時長在輪蟲種間和種內均有差異[8]。以往通常認為,休眠卵會休眠至下一個生長季,乃至數年之后才萌發,相對于輪蟲僅能持續數天至幾周的壽命而言,休眠時間較長[6,9- 11]。然而,一些實驗室研究發現部分休眠卵在產生后的幾天之內便開始孵化,稱為早孵化現象(early hatching)[11- 15]。雖然目前少有確鑿證據顯示野外環境下也存在休眠卵的早孵化現象,但是Gilbert[16]預測這個現象在自然環境中可能十分普遍,并且具有重要意義。原因有兩點,一是為使種群獲得盡可能大的短期適合度,即使已經開始進行有性生殖,種群也應當保持優良的孤雌生殖,而休眠卵的早孵化恰好對此有利;二是在生長季開始了一段時日之后,種群的基因多樣性在自然選擇作用下減小,此時休眠卵的早孵化能為種群帶來新的遺傳變異,其中有可能包含更適應此時環境的基因型[16]。此外,有研究者提出輪蟲休眠卵的孵化時間是一個具有“兩頭下注對策”的生活史特征[10,17],即輪蟲的單克隆種群能通過產生具有不同孵化對策的休眠卵來分攤風險,有研究表明孵化對策多樣性與該克隆群所處環境的不可預測性呈出明顯的正相關[18-20]。
目前,關于休眠卵孵化對策的研究主要集中于對孵化對策的影響因素上[7-8,10],尚不清楚不同輪蟲或同種輪蟲的不同品系在孵化對策上是否存在偏好;孵化對策又如何影響其干雌體后代克隆群的增長與繁殖[11],對于這些問題的定量研究有助于了解早孵化現象對輪蟲種群適合度的影響。
本研究擬采用萼花臂尾輪蟲(Brachionuscalyciflorus)的兩個品系H1、D1為實驗動物模型,通過比較觀測兩種孵化對策下的干雌體單克隆種群的增長及有性生殖率(混交率)的差異,來探究孵化對策對后代種群增長的影響。H1、D1兩品系均采集自北京西海(116°38′87.28″E, 39°94′94.31″N),但是在自然水體中的生長時間不同:H1的生長季為8—9月,是窄溫度生態位品系;而D1的生長季為5—10月,是寬溫度生態位品系[21]。兩品系來源地4—11月部分環境指標數據見表1。本研究可為深入了解輪蟲“休眠”的功能作用及其生活史的適應性進化提供基礎數據,并可為水產養殖中重要餌料生物輪蟲的培養提供一定的理論參考。

表1 北京西海4—11月水體部分環境因子觀測數值
1 材料與方法
1.1 兩種孵化對策的休眠卵和食物的制備
本研究的兩個萼花臂尾輪蟲品系均為本實驗室自野外采集,在實驗室內保種培養的樣本。兩種孵化對策的休眠卵制備方法為:將從克隆群中收集的休眠卵直接孵化,數天之內萌發的可認定為早孵化休眠卵;將收集的休眠卵置于4℃下避光保存40 d后再放到相同孵化條件下萌發,模擬經歷了較長休眠期的晚孵化休眠卵。
輪蟲的食物為蛋白核小球藻(Chlorellapyrenoidesa)(購自中國科學院水生生物研究所淡水藻種庫),經無菌接種后,擴大培養于SE培養液(根據中國科學院水生生物研究所淡水藻種庫的配方進行配置)中,恒溫光照培養(25℃,3000 lx,L∶D=24∶0)。選取指數增長期的藻液,在4℃,7000 r/min離心6 min,收集的濃縮藻液,使用分光光度計測定其原始濃度,4℃儲存備用。
1.2 實驗處理過程
取各品系的休眠卵,兩種孵化對策組各約80枚,分別置于盛有30 mL COMBO培養液的燒杯中孵化,孵化及后續的種群培養條件均為培養溫度20℃、光照強度3000 lx、光周期L∶D=16∶8。挑選健康的、出生2h以內的幼體于6孔板中進行單克隆培養,每孔培養體積為5 mL,培養起始密度為0.2 ind/mL。每個品系各設置4個實驗組:晚孵化-高食物濃度(LH-HF(2×106個細胞/mL)),晚孵化-低食物濃度(LH-LF(5×105個細胞/mL)),早孵化-高食物濃度(EH-HF),早孵化-低食物濃度(EH-LF),每個實驗組10個重復。每12h換一次培養液并觀察一次,統計每個孔中非混交雌體(amictic female,AF)數量、混交雌體(mictic female,MF)數量和個體總數(N),計算混交率MR “MR=MF/N”。統計時,帶有非混交卵的成熟個體計入非混交雌體數量,帶有雄卵或休眠卵的成熟個體計入混交雌體數量,所有個體均計入總數。
1.3 數據分析
對各實驗組種群增長曲線進行Logistic方程擬合,得出方程中參數的估計值。擬合使用的公式為:N=K/(1+ea-rt),其中N為種群數量觀測值,待定參數K為環境容納量,r為內稟增長率,a為積分常數。對混交率、內稟增長率數據進行反正弦轉換。用S-W正態分布檢驗和Levene方差齊性檢驗檢測數據的正態分布和方差齊性,若符合,則對各品系的內稟增長率、最大種群數量、開始進行有性生殖時的種群密度和混交率進行雙因素方差分析,進一步用LSD進行兩兩比較;若數據經一定轉換后仍不滿足進行方差分析的要求,則對數據進行非參數檢驗。實驗統計結果以平均值±標準誤(Mean ± SE)表示,差異顯著性水平設定為0.05,全部統計分析采用SPSS 20.0軟件,并采用Origin 9.0作圖。
2 結果
2.1 不同處理組種群數量隨時間的變化
2.1.1種群數量增長曲線
兩品系不同處理組的種群增長曲線如圖1所示。
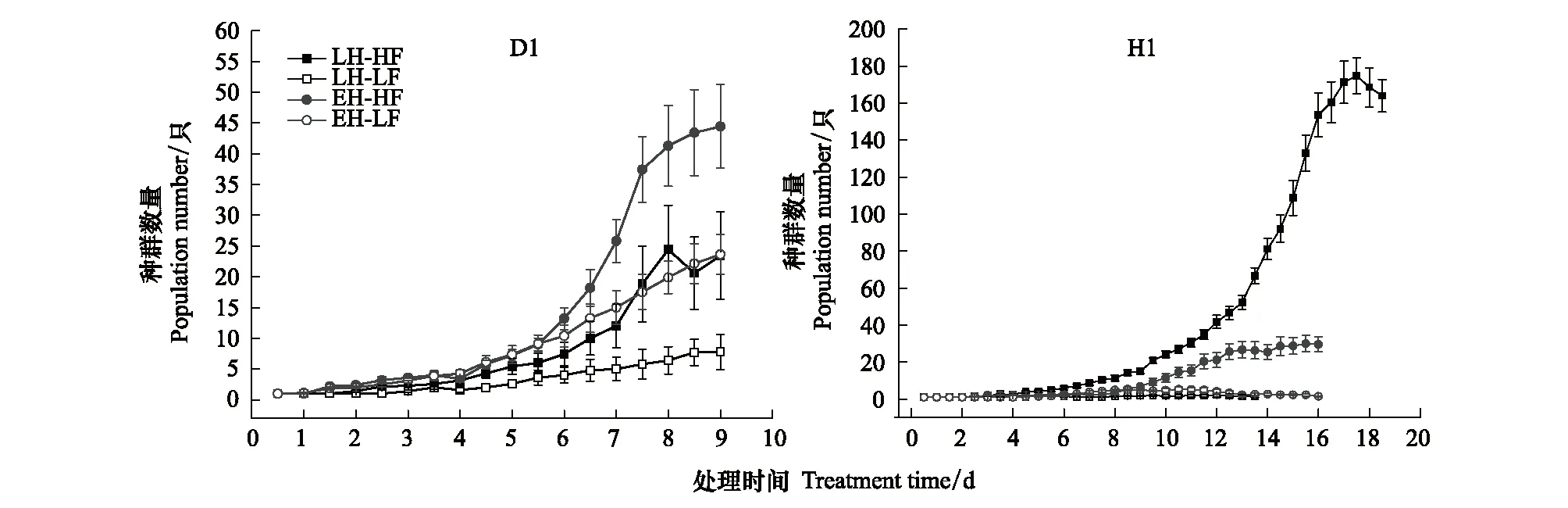
圖1 不同實驗處理組的種群數量增長曲線(數據表示為:Mean±SE)Fig.1 Population growth curve of different experimental groups( Data are shown as Mean±SE)D1:寬溫度生態位品系;H1:窄溫度生態位品系;LH-HF:晚孵化-高食物濃度; LH-LF:晚孵化-低食物濃度; EH-HF:早孵化-高食物濃度; EH-LF:早孵化-低食物濃度
2.1.2Logistic方程擬合
對各實驗組種群增長曲線進行了Logistic方程擬合,獲得K、r的估計值和擬合優度R2,如表2所示。除H1品系的低食物濃度下兩實驗組擬合優度較低外,其余各組均獲得較好地擬合效果。
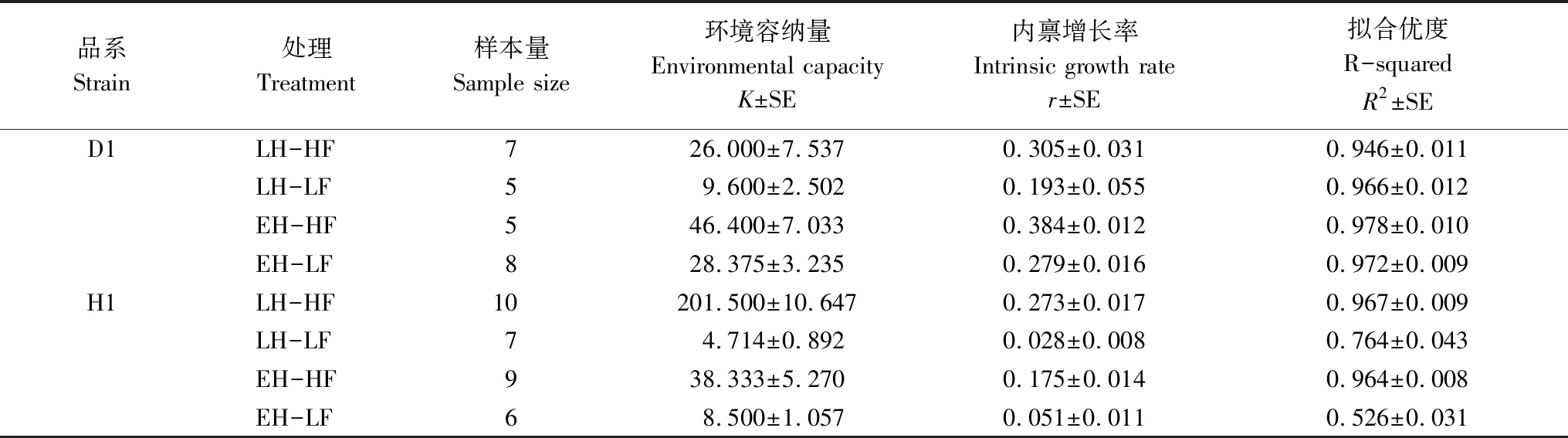
表2 Logistic方程擬合結果
2.1.3內稟增長率
要實現這些目標、路經和模式,任務比較艱巨,其核心人物主要有內部資源管理、內容生產管理、渠道管理、用戶管理和運營績效分析。
對各組內稟增長率數據進行了反正弦轉換后,D1仍不符合進行參數檢驗的要求,因此進行了曼-惠特尼 U 檢驗,發現孵化對策對D1品系的種群內稟增長率沒有顯著影響,食物濃度對種群內稟增長率有顯著影響,高食物濃度更有利于種群增長,如圖2;對H1各組數據進行雙因素方差分析,發現孵化對策和食物濃度兩因素對H1品系的內稟增長率的綜合影響是顯著的(P<0.001),并且兩因素之間存在交互作用(P<0.001),孵化對策對H1品系的內稟增長率影響不顯著(P=0.235),食物濃度對H1品系的內稟增長率影響顯著(P<0.001),進一步將四組數據在單因素下用LSD進行兩兩比較,統計結果如圖2。
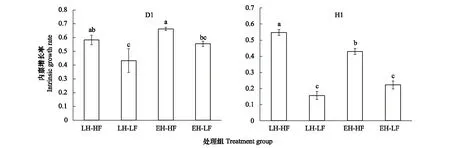
圖2 各處理組內稟增長率的比較(數據表示為:Mean±SE)Fig.2 The intrinsic growth rate of each treatment( Data are shown as Mean±SE)無相同上標字母表示具有統計學顯著差異P<0.05
2.1.4最大種群數量
為使數據滿足方差齊性,符合進行雙因素方差分析的要求,對最大種群數量數據進行了平方根轉換。
如表3所示,孵化對策和食物濃度兩因素對D1品系的最大種群數量具有顯著的綜合影響(P=0.002),而兩者之間不存在交互作用(P=0.911),兩個因素單獨對最大種群數量起作用。其中,孵化對策對D1品系的最大種群數量影響顯著(P=0.001), 食物濃度對D1品系的最大種群數量也有顯著影響(P=0.004,圖3)。
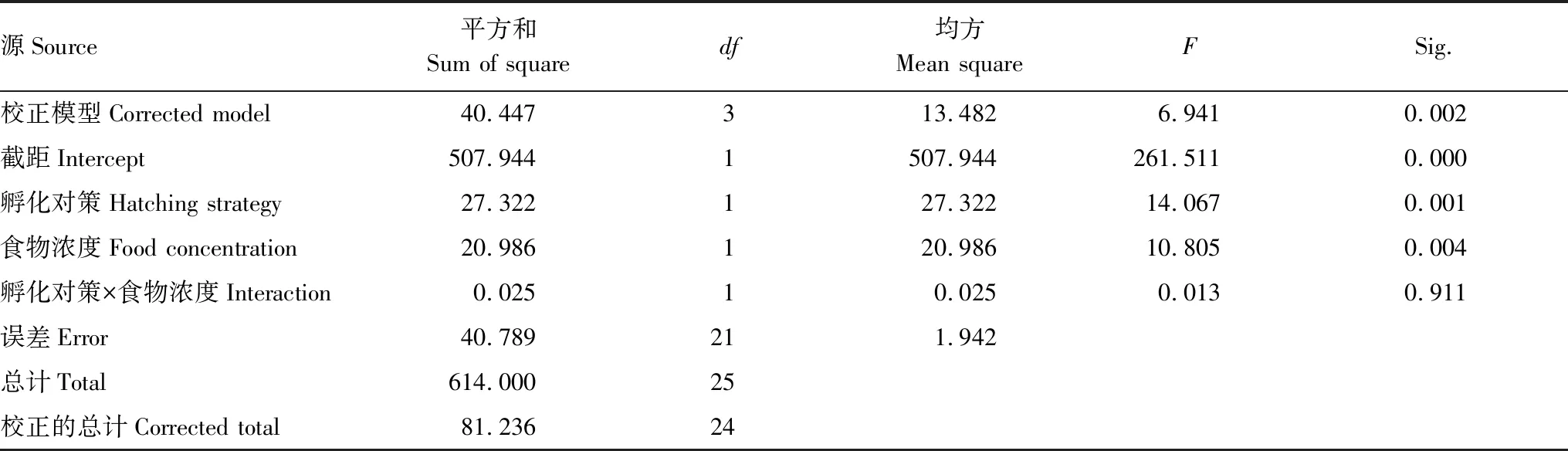
表3 孵化對策與食物濃度對品系D1最大種群數量影響的雙因素方差分析結果
孵化對策和食物濃度兩因素對品系H1的最大種群數量具有顯著的綜合影響(P<0.001,表4),且兩者之間存在交互作用(P<0.001)。孵化對策對H1品系的最大種群數量影響顯著(P<0.001),食物濃度對H1品系的最大種群數量影響也顯著(P<0.001,圖3)。

表4 孵化對策與食物濃度對品系H1最大種群數量影響的雙因素方差分析結果
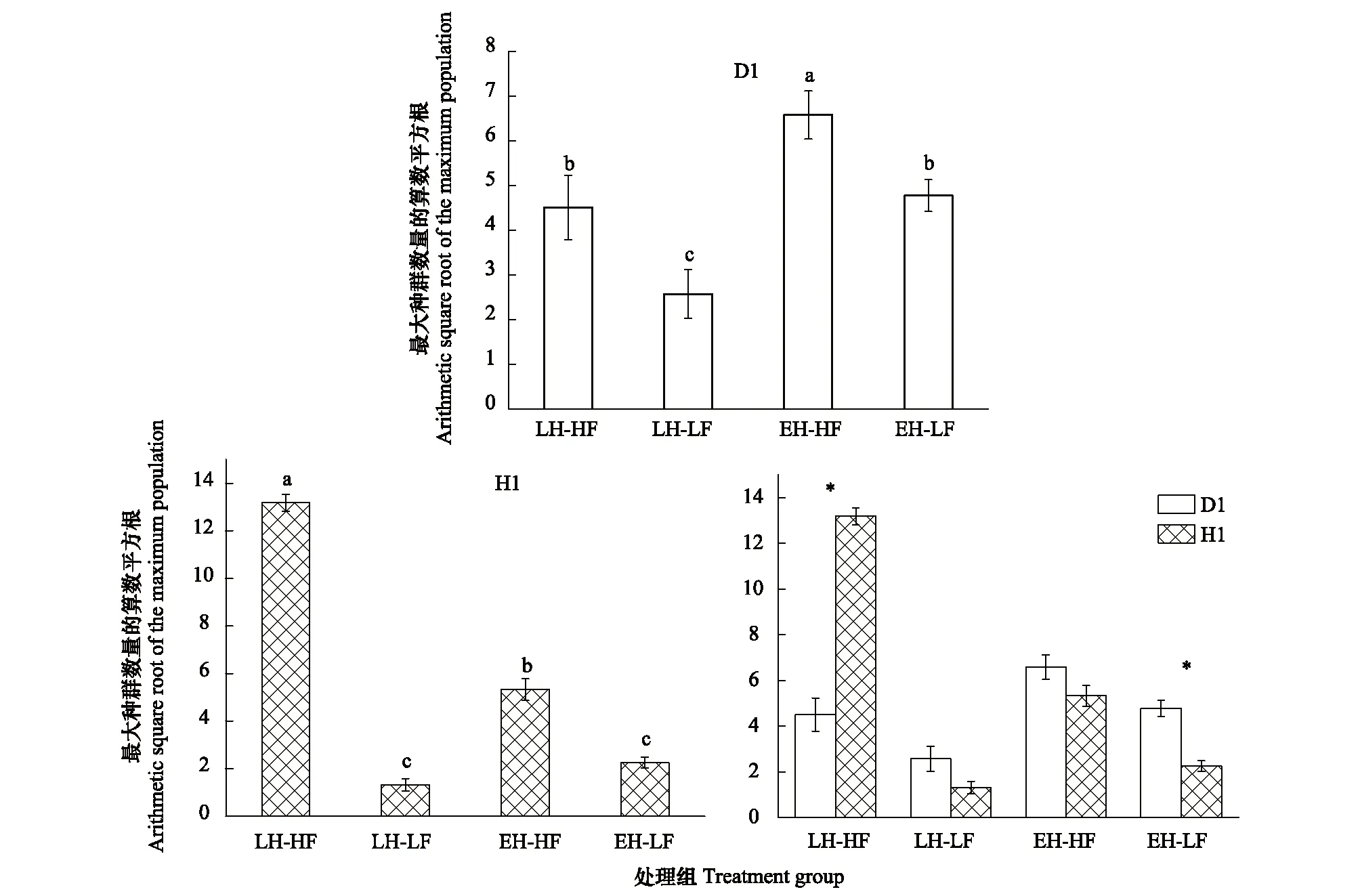
圖3 各處理組最大種群數量的比較(數據表示為:Mean±SE)Fig.3 The maximum population size of each treatment (Data are shown as Mean±SE)無相同上標字母表示具有統計學顯著差異P < 0.05
2.2 有性生殖
2.2.1種群混交率隨時間變化曲線
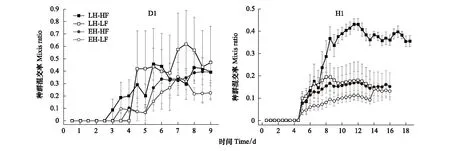
圖4 各處理組混交率隨時間的變化曲線(數據表示為:Mean±SE)Fig.4 Mixis ratio curve of each treatment (Data are shown as Mean±SE)
2.2.2各實驗組種群開始進行有性生殖時的種群密度
對各組數據進行算數平方根轉換后,“低食物濃度”各組數據仍不符合進行雙因素方差分析的要求,因此僅探究了高食物濃度下“孵化對策”這一因素對種群開始進行有性生殖時的密度有何影響。對D1數據進行了單因素方差分析(P=0.041),對H1數據進行了曼-惠特尼U檢驗(P=0.022),結果如圖5所示。
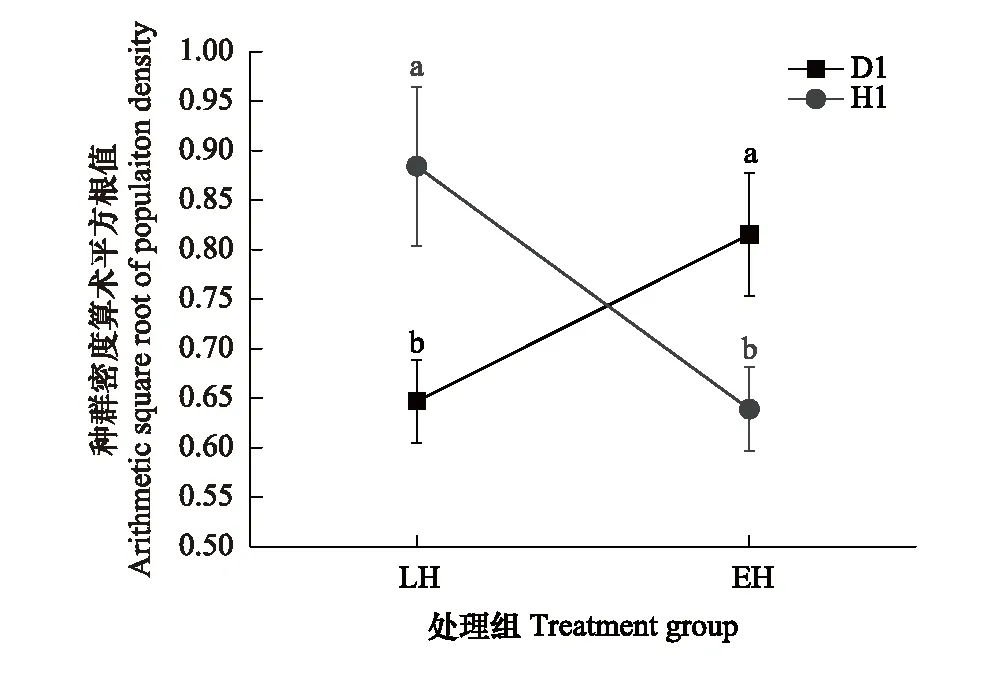
圖5 不同處理組種群開始進行有性生殖時的種群密度比較(數據表示為:Mean±SE)Fig.5 Comparison of the population density at the beginning of sexual reproduction in different treatment groups(Data are shown as Mean±SE)無相同上標字母表示具有統計學顯著差異P<0.05
2.2.3各實驗組種群在最高種群密度下的混交率
對各組混交率數據進行算數平方根轉換后,“低食物濃度”數據仍不符合進行雙因素方差分析的要求,因此仍然僅探究高食物濃度下“孵化對策”這一因素對種群在最高種群密度下的混交率有何影響。對D1、H1數據進行了單因素方差分析,“孵化對策”對D1種群在最高種群密度下的混交率影響不顯著(P=0.855),對H1種群在最高種群密度下的混交率有顯著影響(P=0.044,圖6)。
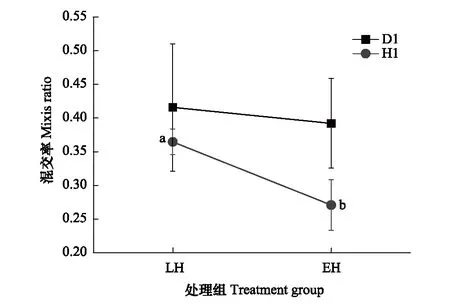
圖6 不同處理種群在最高種群密度下的混交率(數據表示為:Mean±SE)Fig.6 Mixis ratio at peak population density of different experimental groups(Data are shown as Mean±SE)無相同上標字母表示具有統計學顯著差異P<0.05
3 討論
本研究主要觀測了萼花臂尾輪蟲(B.calyciflorus)的兩個品系H1、D1在不同休眠卵孵化對策和不同食物濃度下干雌體形成的單克隆種群增長的差異。本研究發現,休眠卵孵化對策確實對其干雌體克隆種群的增長產生了影響,但具體影響的結果與各品系在自然環境中的生長季的時間有關。同一品系中,內稟增長率和最大種群數量的分析結果表明孵化對策會對種群最終能占領的環境資源多少產生影響,早孵化更有利于生長季時間較長的D1品系的種群增長,晚孵化更有利于生長季時間較短的H1品系的種群增長。
在長期自然選擇壓力下,不同環境、不同品系的輪蟲在與休眠相關的性狀(如休眠期存活率、休眠時長和孵化率等)上會呈現出不同的特點[22-23]。休眠期的長短可能受諸多因素的影響[17-18,22],不同孵化對策所占的比例與萌發后種群增長的表現也會相應受到影響。有研究表明,在底泥環境安全的前提下,水體環境不適宜種群生長的時間越長,休眠卵的休眠期也越長[17]。也就是說,環境適宜的生長季越短,種群越傾向于選擇較長的休眠期,較晚孵化的休眠卵形成的種群能夠獲得更高的種群適合度,本研究的結果支持這一發現。
本研究使用的D1和H1為不同溫度生態位的萼花臂尾輪蟲品系,一年之中適宜寬溫度生態位的品系(D1)生長的時間較長,該品系中早孵化的休眠卵在萌發時很可能仍處于環境適宜的生長季內,因此后代種群能夠以低混交率快速擴增,在適宜生境下參與競爭。而水體中窄溫度生態位品系(H1)生長的時間較短,若過早萌發則可能面臨著生長季的結束,采取晚孵化對策才是一個相對保險的選擇。另外,影響H1品系的最大種群數量的兩個因素(孵化對策和食物濃度)之間存在交互作用,以及Logistic方程擬合的結果(表2)都說明, H1品系對食物濃度的要求較高,當食物缺乏時,食物會超越孵化對策成為影響種群增長的主要因素。H1品系的生長季在夏末秋初,早孵化的個體可能面臨已存在的高種群密度個體對食物的競爭和秋季食物生產將隨光照和溫度下降而降低的雙重壓力,因而晚孵化更有利。
在一個生長季中,產生足夠多的休眠卵可在一定程度上保證種群的長期適合度[24],但進行有性生殖對種群在當下生長季的短期適合度又有明顯代價[25,26],種群需要在二者之間進行權衡[27]。種群開始進行有性生殖時的種群密度可以反映出在增長早期對密度信號響應的靈敏性,再結合種群密度較高(達到環境容納量)時的有性生殖率,可以反映出種群在不同時期通過有性生殖產生休眠卵的傾向性。本研究中,在種群增長的早期,D1品系早孵化的種群比晚孵化的更能經受較高的種群密度,先以較低的有性生殖率迅速擴大種群(圖5),優先保證種群在當下生長季的短期適合度,而不急于產生休眠卵保種,這可能和D1品系生長季較長有關。H1品系恰恰與D1相反,早孵化的種群比晚孵化的種群更傾向于盡早產生休眠卵(圖5)以應對接下來可能面臨的環境脅迫,優先保證種群的長期適合度,對種群增長造成的代價由增長后期相對較低(圖6,相對于晚孵化休眠卵后代種群)的混交率來彌補。
總體上,本研究有助于我們了解休眠卵在孵化時間上的兩頭下注對策與種群自身特性、種群的原始生境之間存在的聯系,很可能是輪蟲種群對生境時間變異的適應性表現,也使我們能夠進一步了解休眠的功能作用,并可以為水產養殖中合理利用和培養餌料生物提供參考。即便是在同一輪蟲品系中,甚至是同一克隆群中,也存在著休眠卵孵化對策的差異[15,19],但原因以及不同孵化對策如何影響后代種群動態還需要更多的定量研究。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:46
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
江蘇安全生產(2020年3期)2020-04-21 05:44:14
華人時刊(2019年13期)2019-11-17 14:59:54
云南教育·中學教師(2019年6期)2019-08-13 07:03:28
基層中醫藥(2018年11期)2019-01-31 05:26:52
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
