布魯氏桿菌在巨噬細胞內可以長期存在的原因
2020-12-07 06:06:23耿慧娟
中西醫結合心血管病電子雜志 2020年28期
耿慧娟
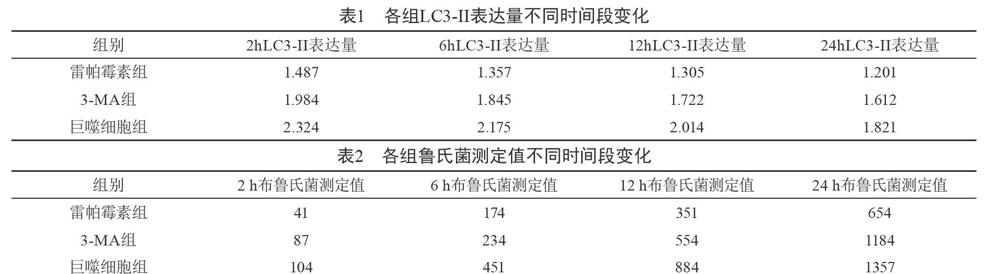
【摘要】目的 觀察分析布魯氏桿菌對巨噬細胞容易感染原因。方法 采用同組體外培養的巨噬細胞進行分組,并分別采用抑制劑3-MA與自噬激活劑雷帕霉素對于巨噬細胞進行處理,并預留一組不做處理,分為巨噬細胞組、3-MA組與雷帕霉素組,處理完成后進行綠色熒光表達的重組豬種布魯氏菌接種,并在接種完成后對于兩組細胞內部的巨噬細胞數量與布魯氏菌數量進行對比,觀察兩種細胞數量變化與細胞內LC3-II的含量變化。結果 在出現布魯氏菌感染后,巨噬細胞發生了一定的自噬作用,雷帕霉素組巨噬細胞內的LC3-II的含量明顯上升且高于其他組;同時雷帕霉素組內的布魯氏菌數量明顯高于另外兩組,而在2 h、6 h、12 h、24 h四個時間段內對比我們發現,各組的布魯氏菌數量均出現上漲的情況,且隨著時間的推移巨噬細胞內的LC3-II的含量明顯增加。結論 在布魯氏菌感染巨噬細胞的過程中,巨噬細胞體內的LC3-II的含量會出現明顯上升,提示自噬作用的發生,但是隨著時間的推移,此種自噬作用雖然能夠殺滅部分布魯氏菌,但是也會使得布魯氏菌在巨噬細胞內部出現增殖的情況,當達到感染數量飽和時,巨噬細胞將無法抑制布魯氏菌的大量增殖。
【關鍵詞】布魯氏桿菌;巨噬細胞;雷帕霉素;3-MA;LC3-II;自噬作用
【中圖分類號】R37 【文獻標識碼】A 【文章編號】ISSN.2095.6681.2020.28..02
細胞的自噬過程是一種細胞基于自我消化與降解的高度保守行為,多是由于真核細胞生物體內為維持機體的環境穩定所進行的一種過程[1]。同時自噬行為的發生往往也是機體內部固有的免疫應答重要標志,在進行自噬的過程中,會對于機體內部細胞中存在的病原體進行有效的清除,同時也能夠在抗原的提呈過程中起到一定的作用[2]。但是近年來有一些文獻指出[3],自噬雖然能夠有效的對于外來入侵的病原體與微生物進行有效的殺滅與清除,但是部分胞內寄生菌與病毒往往能夠通過自身或者機體對于自噬作用進行一定的改變或者逃避,甚至進而在機體內借助大量增殖,引發相關疾病。本次研究中的布魯氏菌就是一種主要寄生與巨噬細胞與胎盤滋養層細胞內部的包內寄生革蘭氏陰性菌種,此類菌種能夠通過其表面的O-鏈LPS對于巨噬細胞表面脂伐產生一定作用,進而進入巨噬細胞內部,改變內部pH值的同時,使自身大量繁殖[4]。但是對于自噬細胞遭受相關感染的原因與在感染過程中是否存在一定的抑制布魯氏菌增長的作用一直未曾明晰。本次研究中巨噬細胞進行分組,并分別采用抑制劑3-MA與自噬激活劑雷帕霉素對于巨噬細胞進行處理,并預留一組不做處理,觀察分析布魯氏桿菌對巨噬細胞容易感染原因,現報告如下。
1 資料與方法
1.1 一般資料
本次研究選取的重組豬種布魯氏菌與小鼠巨噬細胞等實驗材料均來源于外院實驗室采購保存,研究中所采用的相關實驗材料有:LC3-II抗體試劑、3-MA抑制劑、自噬誘導劑Ra、LC3-IIELISA檢測試劑盒。
1.2 實驗方法
具體實驗方法如下:①首先進行重組豬種布魯氏菌與布魯氏菌的培養,均按照1%的比例于培養基中接種,隨后37℃培養24 h,培養完成后進行收獲并置于冰箱內保存與菌落計數;②隨后將巨噬細胞進行常規傳代培養并置于96孔細胞培養板中,2×106個/孔,直至細胞長至單層,隨后選擇終濃度為10nM的Ra與10nM的3-MA進行作用,作用2h后棄細胞液并加入培養基,單孔100uL/孔;③將培養好的布魯氏菌以1∶500的比例接種于96孔細胞培養板中,并分為巨噬細胞組、3-MA組與雷帕霉素組,隨后進行培養在2 h、6 h、12 h、24 h四個時間段加入蛋白酶抑制劑,并隨后進行3此低溫凍融;④對于標本進行離心5 min并取上清液,進行LC3-II含量測定。
1.3 觀察指標
對比三組兩種細胞數量變化與細胞內LC3-II的含量變化。
2 結 果
2.1 檢測結果一覽
在出現布魯氏菌感染后,巨噬細胞發生了一定的自噬作用,雷帕霉素組巨噬細胞內的LC3-II的含量明顯上升且高于其他組;同時雷帕霉素組內的布魯氏菌數量明顯高于另外兩組;而在2 h、6 h、12 h、24 h四個時間段內對比我們發現,各組的布魯氏菌數量均出現上漲的情況,且隨著時間的推移巨噬細胞內的LC3-II的含量明顯增加,見表1、2。
3 討 論
本次研究中所提及的布魯氏菌能夠通過一系列的策略逃避免疫監視,從而改善巨噬細胞內部的環境,從而使得其內部有利于其自身的復制與增殖。國內有研究認為胞內病原微生物主要能夠通過抑制自噬小體的形成或者抑制吞噬小體與溶酶體的融合來干擾機體內部自噬作用的發生,本次研究中采用了3-MA與雷帕霉素對于巨噬細胞的自噬能力進行了放大或者降調,且并為對于巨噬細胞的活力造成影響,同時基于自噬能力的大小對于布魯氏菌生長的影響進行了對比,發現了兩者之間的關聯性[5]。
同時本次研究中對于細胞內LC3-II的表達量進行了一定的測定,此類指標的含量往往是反映細胞內部自噬作用能力的重要指標,通過對于此指標的測定能夠有效的測定自噬作用與布魯氏菌的關聯性[6]。我們發現,布魯氏菌感染后巨噬細胞內LC3-II的表達量明顯增加,并對于巨噬細胞的自噬能力進行了有效的增強,同時也導致了布魯氏菌在巨噬細胞內部的增殖,隨著實驗時間的推進我們不難發現,隨著細胞內LC3-II的表達量下降,細胞內部的布魯氏菌數量卻一直在增加,主要是由于在細胞數量有限的情況下,當布魯氏菌的數量飽和后,自噬細胞雖然啟動了自噬作用,LC3-II為溶酶體降解,并使得布魯氏菌被部分殺滅,但是存留的布魯氏菌已然在其內部大量增殖,這也解釋了布魯氏桿菌對巨噬細胞容易感染的原因所在。
綜上所述,在布魯氏菌感染巨噬細胞的過程中,巨噬細胞體內的LC3-II的含量會出現明顯上升,提示自噬作用的發生,但是隨著時間的推移,此種自噬作用雖然能夠殺滅部分布魯氏菌,但是也會使得布魯氏菌在巨噬細胞內部出現增殖的情況,當達到感染數量飽和時,巨噬細胞將無法抑制布魯氏菌的大量增殖。
參考文獻
[1] 何金科,童志霞,李天森,等.布魯氏菌LPS及VirB2缺失株對巨噬細胞SUMO-1表達及Ubc-9轉錄水平的影響[C].中國動物傳染病學報,2018,14(5):29-34.
[2] 鄧肖玉,易繼海,李啟峰,等.泛素相關修飾蛋白SUMO-2對RAW264.7細胞內布魯氏菌16M存活的影響[J].中國畜牧獸醫,2019,46(8):2257-2264.
[3] 許 達.馬耳他布魯氏菌OmpR基因缺失株的構建及其對布魯氏菌生長與復制的影響[J].中國預防獸醫學報,2018,40(12):1105-1111.
[4] 王姝懿,趙學亮,孫 柯,等.布魯氏菌A19疫苗株全基因組測序及比較基因組學分析[J].中國人獸共患病學報,2019,35(11):1002-1008.
[5] 任 健,章偉東,王玲莉,等.2016—2018年十堰地區布魯氏桿菌感染患者流行趨勢分析[J].公共衛生與預防醫學,2019,30(4):61-64.
[6] 王 成,向 導,田 艷,等.一例布魯氏桿菌病的緊急流行病學調查報告[J].獸醫導刊,2018,22(8):122.
