DIS3對人骨髓瘤細胞周期及腫瘤相關蛋白表達的影響研究
2020-12-09 03:41:16王蘭蘭
中國比較醫(yī)學雜志 2020年11期
鄒 亮,程 輝,張 婷,周 英,關 軍,程 平,王蘭蘭
(武漢市第一醫(yī)院血液內(nèi)科,武漢 430022)
多發(fā)性骨髓瘤(multiple myeloma, MM)是一種漿細胞惡性增殖性疾病,以骨髓內(nèi)惡性漿細胞的克隆增生為其主要特征[1]。目前,針對多發(fā)性骨髓瘤的治療手段雖多,且均能延長了患者的生存時間,但由于各種各樣的并發(fā)癥,導致患者的生存質(zhì)量較差,因此,尋求新的治療方法迫在眉睫[2-3]。研究發(fā)現(xiàn),幾乎所有腫瘤細胞均存在細胞周期異常;因此,腫瘤也被稱為細胞周期病。腫瘤細胞最明顯的特征為不依賴于細胞外生長信號的刺激而能反復自發(fā)的進行細胞周期循環(huán),細胞增殖增加、凋亡細胞減少,最終誘發(fā)細胞不可控的異常增殖而導致腫瘤形成[4]。全基因組測序結果顯示,DIS3基因具有較強的突變特性,DIS3基因突變是導致MM發(fā)生的原因之一[5],但其對骨髓瘤細胞的周期變化及具體作用機制尚不明確。細胞外調(diào)節(jié)蛋白激酶(extracellular regulated protein kinases, ERK)ERK1/2,是將信號傳導的關鍵因子,磷酸化的ERK1/2由胞質(zhì)轉位到核內(nèi),介導ATF,Ap-1,c-fos和c-Jun等因子的活化,參與細胞增殖、分化、凋亡,維持細胞穩(wěn)態(tài)等多種生物學反應[6-7]。
本實驗中,我們通過構建DIS3過表達或干擾載體,觀察過表達或干擾DIS3對細胞周期及腫瘤相關蛋白的表達影響,旨在為臨床治療MM提供潛在的策略。
1 材料和方法
1.1 實驗細胞
人骨髓瘤細胞株NCI-H929(貨號:CRL-9068)、RPMI 8226(貨號:CCL-155)和U266(貨號:TIB-196)購自ATCC。
1.2 主要試劑與儀器
人RPMI-1640細胞培養(yǎng)基、胎牛血清、青-鏈霉素雙抗及Lipofectamine 2000均購于美國Invitrogen生命技術公司;真核表達載體PEGFP-N1由武漢華聯(lián)科生物技術有限公司分子實驗室保存并提供;細胞RNA提取試劑盒、反轉錄試劑盒購自寶日醫(yī)生物技術(北京)有限公司;SYBR Green染料購自Lumiprobe Corporation公司;CycletestTMPlus DNA Reagent Kit(細胞周期檢測)購自BD Biosciences;蛋白提取試劑盒購自上海碧云天生物技術有限公司;一抗:Cyclin B1(貨號:ab2949)、P21(貨號:ab109199)、CDK2(貨號:ab32147)、MYC(貨號:ab32072)、RAS(貨號:ab52939)、TP53(貨號:ab131442)、BRAF(貨號:ab167415)、p-ERK1/2(貨號:ab201015)、GAPDH(貨號:ab9485)以及山羊抗兔二抗IgG(貨號:ab6734)購自美國Abcam公司;主要儀器:CO2培養(yǎng)箱(Thermo,美國);倒置顯微鏡(Nikon,日本);實時熒光定量PCR儀(Bio-RAD,美國);離心機(Eppendorf,德國);流式細胞儀(Beckman,美國)。
1.3 實驗方法
1.3.1 細胞的復蘇、培養(yǎng)及轉染
人骨髓瘤細胞NCI-H929、RPMI 8226和U266分別培養(yǎng)于含10%胎牛血清的RPMI-1640培養(yǎng)基中,培養(yǎng)至對數(shù)期時,按照前期本課題組相應轉染方案[8]進行轉染,轉染48 h后收集細胞進行轉染效率檢測。
1.3.2 PI單染法檢測細胞周期變化
3種細胞均分成5組:對照組、DIS3-siRNA陰性對照(NC)組、DIS3-過表達空載組、DIS3-過表達組、DIS3-siRNA組。按每孔104個細胞進行接種,每孔加100 μL細胞培養(yǎng)液后,置于5% CO2的恒溫培養(yǎng)箱進行培養(yǎng)。48 h后收集各組細胞,混勻后轉移至流式專用管中;離心棄上清沉淀后,用無水乙醇于-20℃冰箱內(nèi)固定細胞24 h,分別加入RNase A和碘化丙啶(propidium iodide,PI)溶液,避光染核10 min。用流式細胞儀進行檢測。
1.3.3 qRT-PCR檢測各指標的mRNA表達水平
根據(jù)NCBI上公布的各基因序列分別設計qRT-PCR用引物(表1),并由上海生工代為合成。48 h后收集各組細胞,分別提取各組細胞樣本RNA,反轉錄,擴增后,以GAPDH為內(nèi)參基因,以2-△△CT計算各基因的相對表達量。

表1 引物序列Table 1 Primer sequence
1.3.4 Western blot
48 h后收集各組細胞,采用BCA蛋白濃度檢測試劑盒對所提蛋白進行濃度檢測;上樣、電泳、封閉后,分別加入兔抗人一抗(Cyclin B1,1∶500、P21, 1∶1000、CDK2,1∶500、MYC,1∶500、RAS,1∶500、TP53,1∶800、BRAF,1∶1000、p-ERK1/2,1∶1000和GAPDH,1∶2000),4℃過夜;加入相應的二抗IgG(羊抗兔,1∶5000),室溫孵育30 min后,以GAPDH為內(nèi)參蛋白,采用Quantity One圖像分析軟件進行灰度比分析。
1.4 統(tǒng)計學方法

2 結果
2.1 DIS3過表達或干擾對細胞周期的影響
3種細胞的細胞周期變化檢測結果顯示(圖1),與空載體組比較,DIS3過表達組細胞發(fā)生了明顯G0/G1期阻滯現(xiàn)象,表現(xiàn)為G0/G1期細胞占比顯著增加(P<0.01);與NC組比較,DIS3-siRNA明顯促進了細胞周期進程,表現(xiàn)為G0/G1期細胞占比的顯著降低(P<0.05,P<0.01)。
2.2 DIS3過表達或干擾對細胞周期相關指標Cyclin B1、P21、CDK2表達的影響
3種細胞的細胞周期相關基因Cyclin B1、P21、CDK2表達檢測結果顯示(圖2A),與空載體組比較,DIS3過表達組Cyclin B1、CDK2表達水平顯著降低(P<0.05),P21表達水平顯著升高(P<0.01);與NC組比較,DIS3-siRNA明顯促進了Cyclin B1、CDK2的表達而抑制了P21的表達(P<0.01)。而三指標在蛋白水平上的表達與mRNA水平表達趨勢基本一致(圖2B)。
2.3 DIS3過表達或干擾對腫瘤相關指標MYC、RAS、TP53及BRAF表達的影響
3種細胞的腫瘤相關基因MYC、RAS、TP53及BRAF表達檢測結果顯示(圖3A),與空載體組比較,DIS3過表達組MYC、RAS、TP53及BRAF表達水平顯著降低(P<0.05,P<0.01);與NC組比較,DIS3-siRNA明顯促進了MYC、RAS、TP53及BRAF的表達(P<0.01)。MYC、RAS、TP53及BRAF在蛋白水平上的表達與mRNA水平表達趨勢基本一致(圖3B)。
2.4 DIS3過表達或干擾對ERK1/2信號通路的影響
3種細胞的ERK1/2信號通路活化檢測結果顯示(圖4),與空載體組比較,DIS3過表達能顯著抑制ERK1/2信號通路的激活,表現(xiàn)在p-ERK1/2的表達明顯降低(P<0.05);與NC組相比,DIS3沉默則顯著增加了ERK1/2信號通路的活性,表現(xiàn)為p-ERK1/2的表達顯著升高(P<0.05)。
3 討論

注:與空載組比較,**P<0.01;與NC組比較,#P<0.05,##P<0.01。圖1 流式細胞術檢測細胞周期變化Note. Compared with the empty vector group,**P<0.01. Compared with the NC group,#P<0.05,##P<0.01.Figure 1 Cell cycle changes were detected by flow cytometry

注:A:各指標mRNA水平的表達;B:各指標蛋白水平的表達;與空載組比較,*P<0.05,**P<0.01;與NC組比較,#P<0.05,##P<0.01。圖2 細胞周期相關指標Cyclin B1、P21、CDK2的表達變化Note. A, mRNA expression. B, Protein expression. Compared with the empty vector group,**P<0.01. Compared with the NC group,#P<0.05,##P<0.01.Figure 2 Changes in the expression of cell cycle-related indicators Cyclin B1, P21 and CDK2

注:A:各指標mRNA水平的表達;B:各指標蛋白水平的表達;與空載組比較,*P<0.05,**P<0.01;與NC組比較,#P<0.05,##P<0.01。圖3 腫瘤相關指標MYC、RAS、TP53及BRAF的表達變化Note. A, mRNA expression. B, Protein expression. Compared with the empty vector group,*P<0.05,**P<0.01. Compared with the NC group,#P<0.05,##P<0.01.Figure 3 Changes in the expression of tumor-related indicators MYC, RAS, TP53 and BRAF
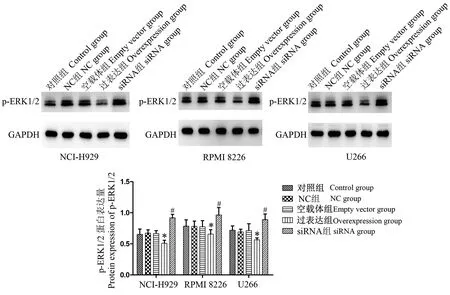
注:與空載組比較,*P<0.05;與NC組比較,#P<0.05。圖4 p-ERK1/2的表達變化Note. Compared with the empty vector group,*P<0.05. Compared with the NC group,#P<0.05.Figure 4 Changes of p-ERK1/2 expression
生理條件下,細胞的增殖與凋亡處于動態(tài)平衡中,細胞周期的異常會導致細胞增殖與凋亡的失衡,是誘發(fā)腫瘤發(fā)生的關鍵環(huán)節(jié)[9]。多發(fā)性骨髓瘤(MM)主要表現(xiàn)為惡性漿細胞無節(jié)制地增生、廣泛浸潤和大量單克隆免疫球蛋白的出現(xiàn)及沉積導致正常多克隆漿細胞增生和多克隆免疫球蛋白分泌受到抑制,從而出現(xiàn)臨床上的異質(zhì)性癥狀及相應并發(fā)癥[10]。DIS3是一種新型的MM抑制基因,其編碼蛋白DIS3能夠?qū)λ械腞NA進行修飾[11-12]。相關臨床研究發(fā)現(xiàn),DIS3突變的MM患者總體中位生存期大大縮短[12-13]。因此,研究DIS3的表達調(diào)控對MM細胞周期的影響具有重要的意義。在本實驗中,DIS3過表達能顯著抑制3種人骨髓瘤細胞的周期進程,表現(xiàn)為顯著的G0/G1期細胞阻滯、細胞周期調(diào)控蛋白Cyclin B1、CDK2的表達減少及P21的表達升高。反之,通過siRNA技術抑制DIS3的表達則從一定程度上促進了癌細胞的周期進程,表現(xiàn)為G0/G1期細胞占比減少、細胞周期調(diào)控蛋白Cyclin B1、CDK2的表達增加、P21表達減少,提示對DIS3進行表達調(diào)控能顯著影響骨髓瘤細胞的細胞周期進程。在以果蠅為研究對象的實驗中發(fā)現(xiàn),DIS3的缺失會抑制細胞的有絲分裂,進而導致幼蟲的生長阻滯及某些組織器官的發(fā)育減緩,提示DIS3的外切酶功能對于細胞有絲分裂是不可或缺的,DIS3的異常會導致細胞的有絲分裂的延遲,表現(xiàn)出非整倍性和過濃縮的染色體[14]。DIS3是結腸癌的新型候選癌基因,沉默DIS3可影響結腸癌細胞的生物學行為[15]。因此,恢復DIS3的表達水平進而維持其功能的穩(wěn)定性對骨髓瘤細胞的惡性增殖可能具有抑制作用,在本實驗中,我們發(fā)現(xiàn)DIS3的抑制作用可能與改變骨髓瘤細胞的細胞周期進程密切相關。此外,我們還發(fā)現(xiàn)DIS3過表達抑制了3種骨髓瘤細胞中癌基因MYC、RAS、TP53及BRAF蛋白水平的表達,提示癌細胞潛在的惡性生物學特征得到了抑制。細胞外調(diào)節(jié)蛋白激酶1/2(ERK1/2)信號轉導通路是絲裂原活化蛋白激酶(MAPK)成員之一[16]。在細胞增殖、分裂、凋亡及惡性轉化等過程中均具有重要的作用,尤其在多種癌細胞中,ERK1/2常處于異常高表達或激活狀態(tài),且被認為是多種藥物的潛在作用靶點[17-19]。例如,在膽囊癌細胞中,青蒿素抑制細胞增殖、抑制CDK4和Cyclin D1表達以及誘導p16表達所造成的細胞G1期阻滯被認為與ERK1/2信號轉導通路的抑制相關[20]。在乳腺癌的體外實驗中發(fā)現(xiàn),厄貝沙坦影響MCF-7細胞的周期,使得細胞的G0/G1期細胞比例增多,而S期細胞比例下降,進而抑制細胞的生長,這一作用被證明與p-ERK1/2蛋白的表達下調(diào)有關[21]。而作為抗腫瘤的化療藥物,卡鉑主要通過誘導細胞周期阻滯抑制腫瘤細胞生長,其對卵巢癌HO-8910細胞作用研究表明,其通過抑制ERK1/2激活,誘導細胞S和G1期阻滯,進而抑制細胞生長[22]。而本研究中,我們同樣發(fā)現(xiàn),DIS3過表達同樣抑制了ERK1/2的激活,而DIS3沉默促進了ERK1/2的活化。
綜上所述,過表達DIS3能通過抑制ERK1/2通路的活性進而抑制骨髓瘤細胞的細胞周期,降低癌基因的表達水平。后期我們將在體內(nèi)水平進行進一步研究,以期為臨床治療MM提供潛在的靶點。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
