海洋曲霉和海洋木霉抗植物病原菌活性次級代謝產物研究進展
2020-12-09 05:31:47黃瑞環韓小斌劉京祝乾湘張遠淑王丹張成省趙棟霖
江蘇農業學報 2020年5期
黃瑞環 韓小斌 劉京 祝乾湘 張遠淑 王丹 張成省 趙棟霖
摘要:植物病原菌是引起植物病害的主要因素之一,給農業生產造成了巨大的經濟損失。真菌病害是植物病害中最大的一類,占植物病害的70%~80%;細菌病害發生面積廣、危害性極其嚴重,并且難以防治。長期以來,植物病原菌的防治依賴于化學農藥,造成了病原菌的抗藥性增強、農藥殘留和環境污染等問題。近年來,研發環境友好、高效、低毒和不易產生抗藥性的生物農藥成為熱點。由于海洋真菌生活在生境獨特的海洋環境中,能夠產生許多結構新穎、生物活性高的次級代謝產物,因而成為具有新穎結構的農用活性天然產物的重要來源。其中,海洋曲霉和海洋木霉是海洋真菌天然產物領域研究較多的屬。本文綜述了1992年至今海洋曲霉和海洋木霉來源的具有抗植物病原菌活性的42個新天然產物,包括聚酮類、萜類、肽類和生物堿類等結構類型,以期為生物農藥的研發提供先導化合物方面的資料。
關鍵詞:海洋曲霉;海洋木霉;次級代謝產物;植物病原菌;抗菌活性
中圖分類號:S432.2+9文獻標識碼:A文章編號:1000-4440(2020)05-1332-10
Abstract:Phytopathogen is one of the main factors that cause plant diseases, resulting in tremendous economic losses in agricultural production. Fungal diseases account for the largest category of all phytopathogens, with a ratio of 70% to 80%. Bacterial diseases occur in a wide area with extreme harmfulness, and are difficult to control. For a long time, the control of phytopathogens relied on chemical pesticide, resulting in problems such as the strengthening of pesticide resistance of phytopathogens, pesticide residues and environmental pollution. In recent years, research and development of biopesticides that are environmentally friendly, efficient, low-toxic and hard to get pesticide resistance have become the hot spots. As marine-derived fungus live in unique marine environment, they can produce many secondary metabolites with novel structures and high biological activities, thus they have become important sources for active natural products with novel structures in agriculture. Among them, the genus of marine-derived Aspergillus sp. and marine-derived Trichoderma sp. have been studied a lot in the field of marine fungus-derived natural products. This article summarized 42 new natural products with anti-phytopathogenic activities of marine-derived Aspergillus sp. and the genus of marine-derived Trichoderma sp. since 1992, including structure types like polyketones, terpenes, peptides and alkaloids, so as to provide lead compounds data for biopesticides development.
Key words:marine-derived Aspergillus sp.;marine-derived Trichoderma sp.;secondary metabolites;phytopathogens;antimicrobial activity
植物病害會阻礙農作物正常生長,導致農作物大量減產、質量下降,從而給農業生產造成重大經濟損失。據統計,全球每年因植物病害造成的農作物產量減幅達10%~16%[1]。植物病原真菌和細菌是引起植物病害的兩大主要因素,真菌病害是植物病害中最大的一類,引起的植物病害多達3×104種,占植物病害總數的70%~80%[2]。據Fisher等[3]估算,真菌病害每年造成水稻、小麥、玉米、馬鈴薯和大豆等5大糧食作物的產量在全球范圍內減少1.25×108 t,僅玉米、小麥和水稻,每年就因真菌病害減產給全球帶來6.00×1010 美元的經濟損失,嚴重威脅著糧食安全。雖然由細菌引起的植物病害種類、受害植物種類及危害程度不及真菌,但植物細菌病害發生面積廣、難以防治[4]。目前已知的植物細菌病害有500種以上,其中作物青枯病、軟腐病、潰瘍病、葉枯病、葉斑病、根癌病和環腐病都是世界性的重要細菌病害[5]。目前,植物真菌病害和細菌病害的防治主要依賴于化學農藥。近年來,中國農藥的總使用量迅速增加,由2001年的1.275 0×106 t迅速增加至2014年的1.807 0×106 t,農藥的單位面積使用量約為世界平均水平的2.5倍[6]。化學農藥長期和大量使用導致病原菌的抗藥性增強,并造成農藥殘留、環境污染以及土壤理化性質和微生物群落破壞等一系列問題[7],因此亟待開發新型生物農藥。與化學農藥相比,微生物農藥來源于自然界,對環境友好、高效、低毒且不易產生抗藥性,因此相關研究越來越受到人們的關注[8]。
隨著陸地環境污染和菌株資源重復利用研究的深入,研究者將尋找微生物農藥的目光轉向了海洋。與陸地微生物相比,海洋微生物生活在高壓、高鹽、低溫、寡營養、有限光照等極端環境中,其代謝方式和途徑不同于陸地微生物,因此海洋微生物的次級代謝產物往往結構新穎、生物活性高,成為新穎活性天然產物的重要來源[9]。海洋真菌由于遺傳背景復雜、代謝產物種類多且產量高,因而成為海洋微生物天然產物研究的焦點[10],其中海洋曲霉和海洋木霉是海洋真菌新天然產物來源的2個重要屬。本文綜述1992年至今海洋曲霉和海洋木霉中具有抗植物病原菌活性的次級代謝產物,以期為進一步開發微生物天然產物農藥提供依據。
1海洋曲霉屬(Aspergillus sp.)和木霉屬(Trichoderma sp.)真菌次級代謝產物研究現狀
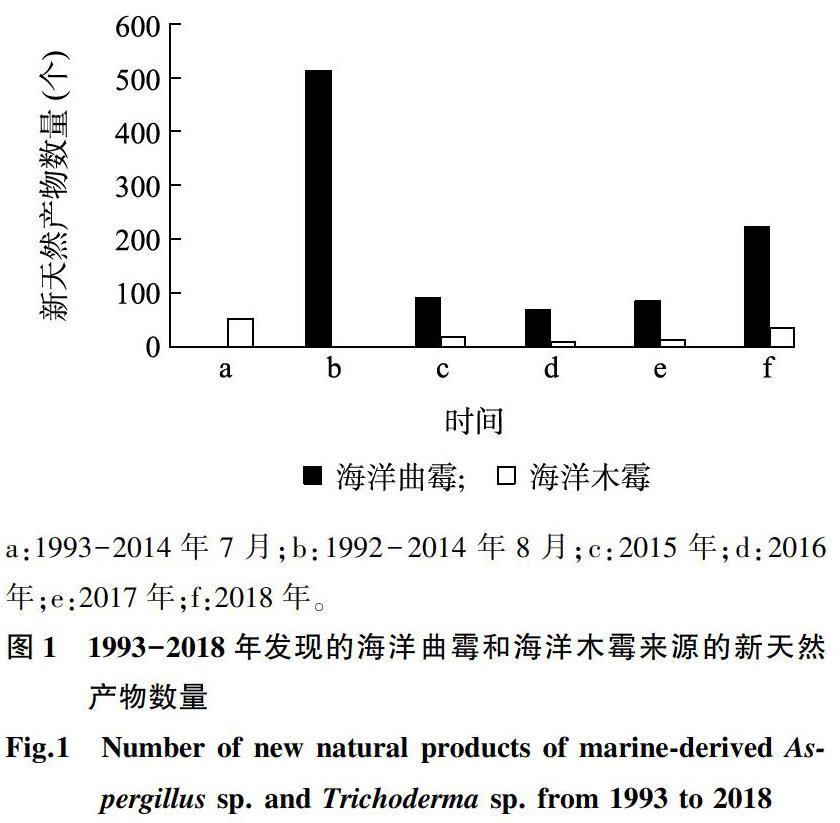
海洋曲霉來源的天然產物研究始于1992年,Numata等報道了首個海洋曲霉來源的新天然產物[11],揭開了海洋曲霉次級代謝產物研究的篇章。截至2018年,研究報道的海洋曲霉來源的新天然產物數量已經超過了979個,包括生物堿類、聚酮類、萜類、甾體類、鹵代物類、脂肪酸類、肽類、糖苷類等多種類型,這些天然產物具有抗癌、抗菌、清除自由基和抗蟲等多種生物活性[10,12-15]。由圖1可以看出,2018年發現的海洋曲霉來源的新天然產物數量多達233個,表明海洋曲霉來源的新天然產物較豐富,并且具有很強的可挖掘性,今后仍是海洋真菌類新天然產物的重要來源屬之一。海洋木霉來源的天然產物的研究始于1993年,但在此后的幾年發展較為緩慢,自2010年以來的相關研究報道占總數的一半以上,是近年來比較活躍的研究領域,僅次于曲霉、青霉來源的天然產物研究[16]。截至2018年,累計發現的海洋木霉來源的海洋木霉新天然產物數量已經超過了127個,且2018年發現的海洋木霉新天然產物數量達35個,其化合物結構類型包括聚酮類、肽類、萜類、脂類等,其中包括一些特殊的三環結構物質、三硫衍生物結構等特殊的骨架結構物質,并且具有抗細菌、抗真菌、殺蟲及對腫瘤細胞具有毒性等生物活性[12-15,17]。
目前,海洋曲霉和海洋木霉來源的天然產物研究主要集中在醫藥領域,關于其防治植物病原菌的報道相對較少。因此,加強海洋曲霉和海洋木霉來源的天然產物在農用抗菌方面的研究,進而用其研發微生物天然產物農藥具有很大潛力。
2海洋曲霉屬真菌抗植物病原菌活性次級代謝產物
2.1抗植物病原真菌活性次級代謝產物
海洋曲霉次級代謝產物農用抗菌活性的研究主要集中在抗植物病原真菌方面。海洋曲霉次級代謝產物豐富、結構類型多樣,海洋曲霉屬是海洋真菌中具有抗植物病原真菌活性的優勢屬之一[18]。目前已有較多報道表明,海洋曲霉活體菌株、菌株提取物及單體化合物具有抑制植物病原真菌的作用。
2010年,沈碩[19]從福建沿海海灘采集的生物樣品中分離到1株海洋曲霉(Aspergillus sp. 1002F2),其水溶性提取物對高粱炭疽病病菌(Colletotrichum graminicola)和番茄早疫病病菌(Alternaria solani)的抑制活性較高,半最大效應質量濃度(EC50)分別為1.34 g/L和0.94 g/L。黃庶識等[20]從廣西北部灣海泥中分離得到短棒曲霉(A. clavatonanicus)MF-13,發現其對荔枝霜疫霉(Peronophythora litchi)、荔枝炭疽病病菌(C. gloeosporioides)、水稻紋枯病病菌(Rhizoctonia solani)和水稻稻瘟病病菌(Magnaporthe grisea)等4種植物病原菌絲的生長均有強的抑制作用。2011年,許蘭蘭等[21]進一步研究發現,MF-13的發酵濾液對荔枝霜疫霉(P. litchi)的抑制效果較好,菌絲生長抑制率達到94%以上,對荔枝炭疽病病菌(C. gloeosporioides)的抑制效果次之,菌絲生長抑制率在70%以上。2013年,祝耀華[22]在福建漳江口的紅樹樣品中分離到135株真菌,其中共有22株對茶葉病原真菌有抑菌活性,曲霉屬是主要屬類,占4.3%。王丹[23]從海南海域樣品中分離的雜色曲霉(A. versicolor)D5、煙曲霉(A. fumigatus)D17、曲霉(Aspergillus sp.)D20對甘藍黑斑病病菌(A. brassicicola)、茶輪斑病病菌(Pestalotiopsis theae)、金橘沙皮病病菌(Diaporthe medusaea)、赤星病病菌(A. alternata)等4種植物病原真菌具有不同程度的抑制作用。
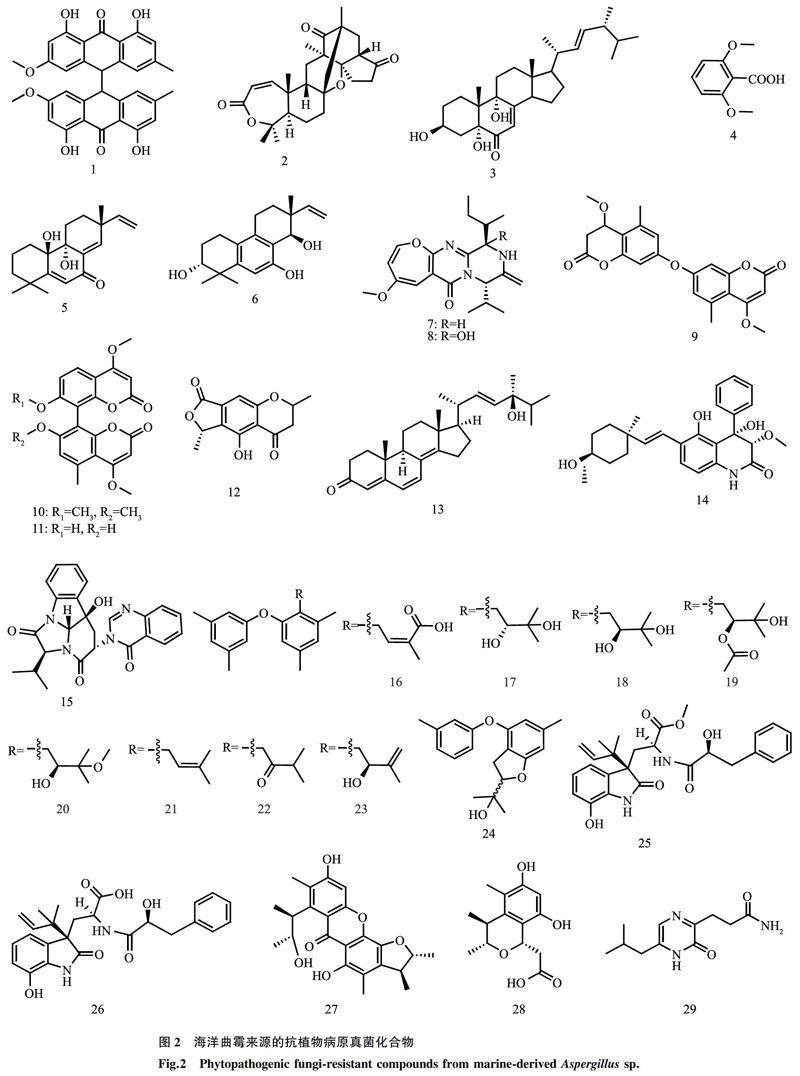
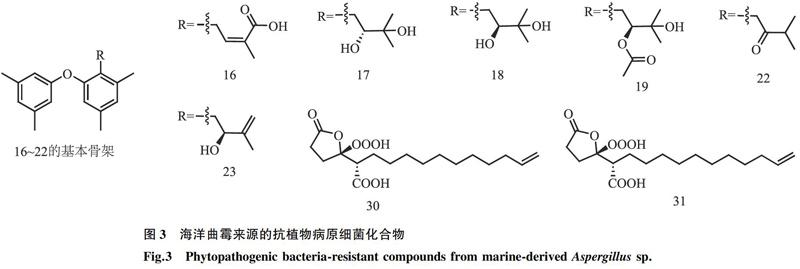
孫好芬[24]從熱帶馬尾藻中分離出1株內生真菌文氏曲霉(A. wentii EN-48),并從中分離得到1個雙核蒽醌類化合物Physcion-10,10′-bianthrone(編號:1)(圖2),發現其對白菜黑斑病病菌(A. brassicae)有一定的抑制活性,抑菌圈直徑為6.0 mm。Cohen等[25]從海綿中分離了1株奇突曲霉(A. insuetus OY-207),其代謝產物混源萜類Insuetolides A(編號:2)有抑制粗糙鏈孢霉(Neurospora crassa)的活性,最小抑菌濃度(MIC)為140 μmol/L。陳卓[26]從海水中分離到1株黃曲霉(編號:MCCC3A00246),采用刃天青顯色法對其代謝產物進行活性跟蹤,結果表明,從該黃曲霉代謝產物中分離到的化合物3β,5α,9α-三羥基-(22E, 24R)-麥角甾-7, 22-二烯-6-酮(編號:3)和2, 6-二甲氧基苯甲酸(編號:4)對黑曲霉(A. niger)有顯著的抑菌作用,抑制率均在90%以上。2016年,Li等[27]從深海海泥中分離到1株文氏曲霉(A. wentii SD-310),并從中鑒定出2個萜類化合物(編號:5、6),結果表明,這2個萜類化合物對禾谷鐮刀菌(Fusarium graminearum)具有顯著的抑菌效果,MIC分別為2.0 μg/ml、4.0 μg/ml,優于陽性對照兩性霉素B(MIC為8.0 μg/ml)。Wang等[28]從深海海泥中分離出1株雜色曲霉(A. versicolor SCSIOO 05879),其產物生物堿化合物Versicolides QA(編號:7)和B(編號:8)對植物炭疽病病菌(C. acutatum)的MIC為1.6 μmol/L(陽性對照放線菌酮對植物炭疽病病菌的MIC為6.4 μmol/L)。5個聚酮類新化合物4,4′-二甲氧基-5,5′-二甲基-7,7′-二豆香豆素(編號:9)、雙香豆素類(Kotanin,編號:10)、Orlandin(編號:11)、色酮衍生物(編號:12)和24-羥基麥角甾-4, 6, 8, 22-四烯-3-酮(編號:13)分離自紅樹內生真菌棒曲霉(A. clavatus)R7[29],其中化合物13對植物病原真菌尖孢鐮刀菌(F. oxysporum)、香蕉炭疽病病菌(C. musae)和青霉菌(Penicillium italicum)具有顯著的抑制作用,MIC分別為224.73μmol/L、195.79μmol/L和61.18 μmol/L,優于陽性對照三唑酮(MIC分別為340.43 μmol/L、272.39 μmol/L和170.24 μmol/L);化合物12對香蕉炭疽病病菌(C. musae)的抑菌效果顯著(MIC為203.07 μmol/L),優于陽性對照;化合物9~11極大地抑制了尖孢鐮刀菌(F. oxysporum),對尖孢鐮刀菌的MIC分別為253.81 μmol/L、235.85 μmol/L和252.47 μmol/L,其抗真菌活性均優于陽性對照。二氫喹啉酮類化合物(編號:14)和喹諾唑啉酮類化合物(編號:15)分離自紅樹欖李新鮮葉中的內生真菌雜色曲霉(A. versicolor)MA-229[30],化合物14對小麥全蝕病病菌(Gaeumannomyces graminis)具有同陽性對照(兩性霉素B)相同的抑菌效果,MIC均為32 μg/ml;在對小麥赤霉病病菌(F. graminearum)的抑菌效果上,化合物15(MIC為16.0 μg/ml)要優于陽性對照(MIC為64.0 μg/ml)。2018年,Li等[31]從海洋藻類內生真菌(A. tennesseensis)中分離到2個具有異戊二烯基二苯醚結構的新化合物Diorcinol L(編號:16)、(R)-diorcinol B(編號:17)和7個已知化合物(S)-diorcinol B(編號:18)、9-acetyldiorcinol B(編號:19)、Diorcinol C(編號:20)、Diorcinol D(編號:21)、Diorcinol E(編號:22)、Diorcinol J(編號:23)、Dihydrobenzofuran derivative(編號:24),這些化合物對赤星病病菌(A. alternata)、玉米小斑病病菌(Cochliobolus heterostrophus)、小麥全蝕病病菌(G. graminis)、圍小叢殼真菌(Glomerella cingulata)、毛霉(Mucor hiemalis)和根黑腐爛病病菌(Thielaviopsis basicola)分別顯示出不同程度的抗菌活性,MIC為2~64 μg/ml。楊遂群[32]在青島海域的海藻和紅樹中分離鑒定得到3株海洋曲霉:阿拉巴馬曲霉(A. alabamensis)、聚多曲霉(A. sydowii)和構巢曲霉(A. nidulans),并采用發酵優化和共培養等方法對其次級代謝產物進行研究,新的吲哚二酮嗎啉類衍生物AL1(編號:25)和AL2(編號:26)分離自阿拉巴馬曲霉EN-547,新的化合物橘青霉素類衍生物SC1(編號:27)和SC2(編號:28)通過將聚多曲霉EN-534和橘青霉EN-535進行共培養后分離鑒定得到,氧雜蒽酮類衍生物ND2(編號:29)分離自構巢曲霉MA-143,化合物25~29對多種植物病原菌均具有顯著的抑菌效果,其中化合物25和26對小麥紋枯病病菌(R. cerealis)、蘋果炭疽病病菌(C. gloeosporioides)等5種病菌的MIC為16~64 μg/ml;化合物SC1(編號:27)和SC2(編號:28)對小麥赤霉病病菌(F. graminearum)等3種病菌的MIC為16~64 μg/ml;化合物ND2(編號:29)對番茄枯萎病病菌(F. oxysporum)等3種病原菌的MIC為8~64 μg/ml。
1株棘孢木霉(T. asperellum)分離自南極沉積物[43],從中分離鑒定的6個抗菌肽類代謝產物Asperelines A~F(編號:32~37)(圖5)具有乙酰化的N末端,并含有不常見的脯氨醇殘基的C末端新穎結構,抗菌活性結果表明,這6個抗菌肽類代謝產物對稻瘟病病菌(M. grisea)和番茄早疫病病菌(A. solani)有弱抑菌活性。2010年,劉旭[44]研究了分離自山東威海海域的鴨毛藻共生真菌哈茨木霉(T. harzianum)EN-85的次級代謝產物,并對其粗提物和單體化合物進行了活性篩選,結果表明,該粗提物對白菜黑斑病病菌(A. brassicae)和蘋果輪紋病病菌(Botryosphaeria dothidea)具有中等抑制作用,分離的化合物中已知化合物Cyclonerodiol FS3(編號:38)和新化合物FS4(編號:39)對蘋果輪紋病病菌(B. dothidea)和棉花枯萎病病菌(F. oxysporium)有弱抑制活性。2019年,鄒積雪等[45]對分離自大連海域松節藻的長枝木霉(T. longibrachiatum)DL5-4進行實驗室規模化發酵培養,從中分離到3個化合物2′,3′-dihydrosorbicillin(編號:40)、Sohirnones A(編號:41)和Sorbicillin(編號:42),發現這3個化合物對圍小叢殼真菌(G. cingulata)有顯著的抑制作用,抑菌圈直徑分別為13.3 mm、12.5 mm和12.6 mm,均強于陽性對照(兩性霉素B的抑菌圈直徑為12.3 mm)。
在以上總結的11個具有抗植物病原真菌活性的海洋木霉次級代謝產物(單體化合物)中,包括6個肽類化合物、2個倍半萜類衍生物和3個聚酮類化合物,分別來源于棘孢木霉、哈茨木霉和長枝木霉。
4展望
本文共總結了具有抗植物病原菌活性的海洋曲霉和海洋木霉,其中紅樹來源的最多,占54.1%,其次是海藻來源的,占27.8%,海洋動物、海泥來源的均占6.6%。從具有抗植物病原菌活性的海洋曲霉和海洋木霉來源看,海藻、紅樹是這2種海洋真菌的重要宿主。在海洋曲霉和海洋木霉次級代謝產物中,被報道具有抗植物病原菌活性的化合物有42個,其結構類型多樣,包括聚酮類、萜類、肽類、生物堿類等,其中聚酮類(占比為57.1%)、萜及混源萜類(占比為14.3%)和肽類(占比為14.3%)是被報道具有抗植物病原菌活性較多的結構類型。
海洋曲霉次級代謝產物在對植物病原真菌抗性上的報道相對于對植物病原細菌抗性上的報道來說較多,但與目前從海洋曲霉中發現的新天然產物的數量相比,占比很小,不足5%。其主要原因可能是目前海洋曲霉天然產物研究的側重點在醫藥領域,在農用抗菌活性上的研究不夠深入;相對于已知化合物而言,科學家們對新化合物的興趣更大,對一些已知化合物的活性篩選也大多停留在抗癌和抗人體病原菌等方面,對其農用抗菌活性篩選的研究較少。海洋木霉在農用抗菌上的報道更多的是在活性菌株和提取物層面,對單體化合物的報道還停留在淺層,還需要結合抑菌機制和田間試驗等進行多層次、多方面的深入研究。筆者所在課題組近年來致力于海洋真菌農用活性物質的研究,從山東、海南沿海分離獲得300余株海洋真菌,針對果斑病病菌(A. avenae)、番茄潰瘍病病菌(Clavibater michiganensis)、白菜軟腐病病菌(Erwinia carotovora)、黃瓜角斑病病菌(P. syringae)、青枯雷爾氏菌(R. solanacearum)、柑橘潰瘍病病菌(Xanthomonas campestris)和水稻白葉枯病病菌(Xanthomonas oryzae pv. oryzae)等主要植物病原細菌,以及煙草赤星病病菌(A. alternate)、甘藍黑斑病病菌(A. brassicicola)、花生冠腐病病菌(A. niger)、黃瓜灰霉病病菌(Botrytis cinerea)、蘋果輪紋病病菌(B. dothidea)、葡萄炭疽病病菌(C. gloeosporioides)、金橘沙皮病病菌(D. medusaea)、稻瘟病病菌(M. grisea)、茶輪斑病病菌(P. theae)、煙草黑脛病病菌(P. parasitica var. nicotianae)、蘋果腐爛病病菌(Valsa mali)、小麥紋枯病病菌(R. cerealis)等植物病原真菌進行了抗菌活性篩選,發現獲得的曲霉屬真菌和木霉屬真菌對不同植物病原菌均表現出不同程度的抗菌活性,對植物病原真菌的抗菌活性更為明顯。而對于植物病原細菌而言,具有抗青枯病病菌和水稻白葉枯病病菌活性的海洋真菌提取物則更難篩選,與之相反的是,黃瓜角斑病病菌、黃瓜果斑病病菌和番茄潰瘍病病菌對海洋真菌提取物更為敏感[46-47]。雖然海洋真菌在防治植物病原菌上具有巨大潛力,但不可否認的是,其應用也具有一定的限制因素。首先,部分植物病原菌活性模型需要的樣品量較大,而海洋微生物活性物質的產量較小,無法滿足實驗室和盆栽試驗所需的產量,對于田間試驗更是“望洋興嘆”;其次,海洋生物資源的采集、純化和制備等成本較高,需要足夠的經費支持,而農藥研發對于成本要求尤其苛刻,這是阻礙其產業化發展的另一難題。以上問題并不是沒有解決辦法,農藥研發與醫藥研發有很大的不同,不需要極高的純度和各種異構體的拆分,發酵產物經過簡單純化即可使用[48]。基于此,可以通過生物合成方法提高活性菌株產生活性成分的產量,并結合有機合成擴大其應用前景。
海洋面積遼闊,海洋曲霉和海洋木霉資源豐富,而目前被分離鑒定的只是很少一部分。隨著對海洋資源的大力開發和利用,海洋曲霉和海洋木霉將為微生物天然產物農藥的研發提供重要的先導化合物來源。
參考文獻:
[1]CHAKRABORTY S, NEWTON A C. Climate change, plant diseases and food security: an overview[J]. Plant Pathology, 2011, 60(1):2-14.
[2]向梅梅. 植物病原真菌分子生物學研究進展[J]. 仲愷農業工程學院學報,2001,14(4):52-58.
[3]FISHER M C, HENK D A, BRIGGS C J, et al. Emerging fungal threats to animal, plant and ecosystem health[J]. Nature, 2012, 484 (7393): 186-194.
[4]彭煒. 植物細菌性病害和病原細菌分類研究進展[C]//中國植物保護學會. 2010年中國植物保護學會學術年會論文集. 鶴壁:中國植物保護學會,2010:166-175.
[5]何晨陽,陳功友. 我國植物病原細菌學的研究現狀和發展策略[J]. 植物保護,2010,36(3):12-14.
[6]王佳新,李媛,王秀東,等. 中國農藥使用現狀及展望[J]. 農業展望,2017,13(2):56-60.
[7]陳曉明,王程龍,薄瑞. 中國農藥使用現狀及對策建議[J]. 農藥科學與管理,2016,37(2):4-8.
[8]袁瑞. 微生物農藥在植物病蟲害防治中的應用策略探討[J]. 農業與技術,2017,37(10):17.
[9]李俊峰,韓曉紅,段效輝. 海洋微生物活性物質研究進展[J]. 氨基酸和生物資源,2014,36(4):12-16.
[10]趙成英,劉海珊,朱偉明. 海洋曲霉來源的新天然產物[J]. 微生物學報,2016,56(3):331-362.
[11]NUMATA A, TAKAHASHI C, MATSUSHITA T, et al. Fumiquinazolines, novel metabolites of a fungus isolated from a saltfish[J]. Tetrahedron Letters, 1992, 33 (12): 1621-1624.
[12]CARROLL A R, COPP B R, DAVIS R A, et al. Marine natural products[J]. Natural Product Reports, 2020, 37(2):175-233.
[13]CARROLL A R, COPP B R, DAVIS R A, et al. Marine natural products[J]. Natural Product Reports, 2019, 36(1): 122-173.
[14]BLUNT J W, CARROLL A R, COPP B R, et al. Marine natural products[J]. Natural Product Reports, 2018, 35(1): 8-53.
[15]BLUNT J W, COPP B R, KEYZERS R A, et al. Marine natural products[J]. Natural Product Reports, 2017, 34(3): 235-294.
[16]宋銀平. 海洋藻棲木霉次生代謝調控的研究[D]. 煙臺:中國科學院煙臺海岸帶研究所,2016.
[17]朱統漢,陳正波,王文玲,等. 非曲霉和青霉來源的海洋真菌新天然產物研究進展[C]//中國藥學會. 中國藥學大會暨第十四屆中國藥師周論文集. 石家莊:中國藥學會,2014:3222-3225.
[18]王丹,芶劍渝,韓小斌,等. 海洋真菌次級代謝產物在植物保護中的研究與應用[J].中國生物防治學報,2019,35(1):146-158.
[19]沈碩. 活性海洋真菌的鑒定及其次級代謝產物的研究[D]. 福州:福建農林大學,2009.
[20]黃庶識,許蘭蘭,黃曦,等. 3株抗水稻和荔枝病原菌的海洋真菌的分離鑒定[J]. 基因組學與應用生物學,2010,29(4):63-70.
[21]許蘭蘭,黃曦,李昆志,等. 海洋真菌的篩選及其對離體荔枝果霜霉病和炭疽病的防效[J]. 中國生物防治學報,2011,27(2):214-220.
[22]祝耀華. 紅樹林沉積物的真菌多樣性及其代謝產物的生物活性研究[D]. 汕頭:汕頭大學,2013.
[23]王丹. 海洋真菌農用生物活性篩選及木賊鐮刀菌D39次級代謝產物研究[D]. 北京:中國農業科學院,2019.
[24]孫好芬. 兩株熱帶馬尾藻內生真菌次生代謝產物研究[D]. 青島:中國科學院海洋研究所,2012.
[25]COHEN E, KOCH L, THU K M, et al. Novel terpenoids of the fungus Aspergillus insuetus isolated from the mediterranean sponge Psammocinia sp. collected along the coast of Israel[J]. Bioorganic & Medicinal Chemistry, 2011,19(22):6587-6593.
[26]陳卓. 高通量抗菌活性篩選模型的構建及兩株海洋真菌次級代謝產物的研究[D]. 廈門:廈門大學,2014.
[27]LI X D, LI X M, LI X, et al. Aspewentins D-H, 20-nor-isopimarane derivatives from the deep sea sediment-derived fungus Aspergillus wentii SD-310[J]. Journal of Natural Products, 2016, 79(5): 1347.
[28]WANG J F, HE W J, HUANG X L, et al. Antifungal new oxepine-containing alkaloids and xanthones from the deep-sea-derived fungus Aspergillus versicolor SCSIO 05879[J]. Journal of Agricultural and Food Chemistry, 2016, 64(14): 2910-2916.
[29]LI W S, XIONG P, ZHENG W X, et al. Identification and antifungal activity of compounds from the mangrove endophytic fungus Aspergillus clavatus R7[J]. Marine Drugs, 2017, 15(8): 259.
[30]羅寒,李曉棟,李曉明,等. 紅樹林來源內生真菌雜色曲霉Aspergillus versicolor MA-229 次級代謝產物研究[J]. 中國抗生素雜志,2017,42(4):334-340.
[31]LI Z X, WANG X F, REN G W, et al. Prenylated diphenyl ethers from the marine algal-derived endophytic fungus Aspergillus tennesseensis[J]. Molecules, 2018, 23(9): 2368.
[32]楊遂群. 五株海藻及紅樹林來源真菌次級代謝產物的分子多樣性挖掘與生物活性研究[D]. 北京:中國科學院大學,2018.
[33]SWATHI J, SOWJANYA K M, NARENDRA K, et al. Isolation, identification & production of bioactive metabolites from marine fungi collected from coastal area of Andhra Pradesh, India[J]. Journal of Pharmacy Research, 2013, 6(6): 663-666.
[34]WANG R, GUO Z K, LI X M, et al. Spiculisporic acid analogues of the marine-derived fungus, Aspergillus candidus strain HDf2, and their antibacterial activity[J]. Antonie van Leeuwenhoek: Journal of Microbiology and Serology, 2015, 108(1): 215-219.
[35]解樹濤,宋曉妍,石梅,等. 康寧木霉(Trichoderma koningii)SMF2分泌的Peptaibols類抗菌肽Trichokonins抑菌活性研究[J]. 山東大學學報(理學版),2006,41(6):140-144.
[36]GAL H I, ATANASOVA L, KOMON Z M, et al. Marine isolates of Trichoderma spp. as potential halotolerant agents of biological control for arid-zone agriculture[J]. Applied and Environmental Microbiology, 2011, 77(15):5100-5109.
[37]陶晶晶. 幾株海洋生境芽孢桿菌和木霉菌農藥潛力的研究[D]. 青島:青島科技大學,2011.
[38]賈煒,田黎,陶晶晶,等. 一株南極生境來源木霉菌株抗菌及誘導植物抗鹽抗寒作用的初步研究[J]. 極地研究,2011,23(3):189-195.
[39]孫健健. 海洋生境木霉與芽孢桿菌的生防應用潛力研究[D]. 青島:青島科技大學,2013.
[40]何海兵. 兩株禾谷鐮刀菌拮抗真菌的次級代謝產物研究[D]. 杭州:浙江大學,2016.
[41]李闖. 3株海洋生境木霉與農藥相關性狀的研究[D]. 青島:青島科技大學,2017.
[42]黃瑞環,芶劍渝,韓小斌,等. 煙草主要病害拮抗菌的篩選鑒定及除草活性分析[J]. 煙草科技,2019,52(12):17-22.
[43]REN J W, XUE C M, TIAN L, et al. Asperelines A-F, peptaibols from the marine-derived fungus Trichoderma asperellum[J]. Journal of Natural Products, 2009,72(6):1036-1044.
[44]劉旭. 蒲枝凹頂藻、鴨毛藻及鴨毛藻共生真菌次生代謝產物研究[D]. 青島:中國科學院研究生院(海洋研究所),2010.
[45]鄒積雪,季乃云. 松節藻來源長枝木霉DL5-4化學成分及其生物活性研究[J]. 化學與生物工程,2019,36(3):18-22.
[46]HUANG R H, GOU J Y, ZHAO D L, et al. Phytotoxicity and anti-phytopathogenic activities of marine-derived fungi and their secondary metabolites[J]. RSC Advances, 2018, 8(66):37573-37580 .
[47]ZHAO D L, WANG D, TIAN X Y, et al. Anti-phytopathogenic and cytotoxic activities of crude extracts and secondary metabolites of marine-derived fungi[J]. Marine Drugs, 2018, 16 (1):36.
[48]吉沐祥,王曉琳,黃潔雪,等.草莓枯萎病菌拮抗細菌JX-13的鑒定及生防效果評價[J].江蘇農業學報,2019,35(3):586-593.
(責任編輯:徐艷)
