
外源氮添加對2種豆科牧草功能性狀的影響
2020-12-09 13:53:06張中信岳奇奇錢申吳靜陶冶
江蘇農業學報 2020年5期
張中信 岳奇奇 錢申 吳靜 陶冶
摘要:以2種常見豆科牧草紫花苜蓿(Medicago sativa)和紫云英(Astragalus sinicus)為材料,設置對照、低氮和高氮(0 kg/hm2、50 kg/hm2、100 kg/hm2)3個梯度的外源氮添加處理,對比研究了2種豆科植物苗期形態、生物量分配、含水量及葉片光合色素含量等功能性狀對外源氮添加的響應及性狀間的權衡特征。結果表明:隨施氮量增加,紫花苜蓿形態性狀及含水量根冠比,紫云英光合色素特征及葉寬、株高、葉厚均有顯著變化(P<0.05),但變化趨勢大多不相同。2種豆科牧草形態性狀總體表現為低促高抑效應,而葉綠素b和類胡蘿卜素含量則表現為施氮后明顯降低。相比之下,氮添加對2種豆科牧草生物量根冠比、含水量、葉綠素a及總葉綠素含量均無顯著影響(P>0.05)。主成分分析結果表明,紫花苜蓿和紫云英前2個主成分所反映的功能性狀協同性在種間差異較大。可見,2種豆科牧草生長和光合色素特征對外源氮添加具有明顯的差異化響應,并表現出不同的協同性。
關鍵詞:氮;紫花苜蓿;紫云英;功能性狀;協同性
中圖分類號:Q948文獻標識碼:A文章編號:1000-4440(2020)05-1197-08
Abstract:In this study, two common leguminous forage plants, Medicago sativa and Astragalus sinicus, were used as materials, and three nitrogen addition treatments, i.e., control (0 kg/hm2), low nitrogen level (50 kg/hm2) and high nitrogen level (100 kg/hm2), were set up. Moreover, the responses of functional traits including morphology, biomass allocation, water content and photosynthetic pigments of two leguminous forage seedlings to exogenous nitrogen addition, and the trade-off characteristics among functional traits were comparably analyzed. The results indicated that with the increase of nitrogen application rate, the morphological traits and root-shoot ratio of water content of M. sativa, the photosynthetic pigment characteristics, leaf width, plant height and leaf thickness of A. sinicus were significantly changed (P<0.05), but the change trends were different in most cases. The morphological traits of two leguminous forage plants presented a low-promotion and high-inhibition effect, while the chlorophyll b and carotenoid contents decreased obviously after nitrogen input. In contrast, nitrogen addition showed no significant effect on root-shoot ratio of biomass, water content, chlorophyll a and total chlorophyll contents of two leguminous forage plants (P>0.05). The results of principal component analysis showed that the synergism of functional traits reflected by the first two principal components differed greatly among the two species. Consequently, the growth and photosynthetic pigment characteristics of the two leguminous forage plants exhibited differential responses to exogenous nitrogen addition, and also showed different covariant patterns.
Key words:nitrogen;Medicago sativa;Astragalus sinicus;functional traits;synergism
氮素是植物生長的必需元素,也是植物生長的主要限制性元素之一[1-4]。隨著工農業發展、人口增長及全球環境變化,東亞地區氮沉降量逐漸增高,已成為繼歐、美之后的第3大氮沉降集中區[2]。大氣氮沉降可以給土壤帶來一定量的外源性氮輸入,對植物生長有一定的促進作用,且適量的氮添加也能部分緩解干旱對植物生長的影響[4-6],對農田作物生長有一定益處。但當氮沉降達到或超過一定量時,氮在土壤及植物體內積累過多,植物生長會受到限制[2]。不僅如此,高濃度的氮沉降會給陸地及水生生態系統的生產力、穩定性以及生物多樣性帶來顯著的負面影響[1,3-4,7-9]。因此,研究氮沉降等外源性氮輸入對不同功能群植物的影響有助于深入揭示植物適應對策,并對厘清農田、森林等不同陸地生態系統對氮沉降的綜合響應具有重要意義。
非固氮植物是研究氮沉降效應的主體材料。大部分非固氮植物的生長本身受到氮素的限制,因此對氮添加的響應極為敏感[10-11]。而具有與根瘤菌共生固氮能力的植物,如豆科植物,其共生體所固定的氮,足以滿足植物自身生長需求[12]。盡管如此,在氮沉降量日益增大的背景下,這些豆科植物依然要面對外源氮輸入帶來的影響,這是不可忽視的現實問題。有研究者發現,一定量(0~40 kg/hm2)的氮添加能夠促進3年生紫花苜蓿(Medicago sativa)的生長并提高光合性能[13],但是不同量的外源性氮輸入對豆科牧草苗期生長和生理的影響尚不明晰。
植物在與環境相互作用過程中,逐漸形成了很多外部形態和內在生理方面的適應對策[14-15]。功能性狀是指對生態功能的形成產生影響,并對個體適合度和自身表現產生影響的性狀,例如植株高度、葉片大小、生物量分配、光合能力等[14-17]。功能性狀之間具有表型整合或表型協變關系,能客觀地反映植物對自身各功能之間權衡及對環境變化的適應性[18]。氮添加能改變植物生長環境,而植物也能通過功能性狀及其組合的變化調整環境變化(包括氮沉降)影響下的響應策略[2]。例如,氮添加使植物莖和葉生物量分配增加,進而顯著降低根冠比,且在營養生長與繁殖上也會出現生物量權衡分配[10]。因此,采用多功能性狀整合的方法有利于揭示豆科植物對氮增加的響應和適應策略。
本研究以2種豆科常見牧草紫花苜蓿和紫云英(Astragalus sinicus)為研究對象,設置3種氮施加梯度(最高為100 kg/hm2),采用大棚盆栽培養的方法,測定植物苗期形態指標、生物量、含水量及光合色素含量,采用單因素方差分析、相關性分析和主成分分析法,對比研究2種豆科植物多種功能性狀對氮施加的響應以及功能性狀間的相關性和協調性,以綜合了解氮沉降等外源性氮輸入對豆科植物生長影響,為氮沉降量逐漸增加情況下的植物生長權衡策略解析及生態系統管理提供科學依據。
1材料與方法
1.1試驗材料與試驗設計
紫花苜蓿和紫云英種子采自安慶市宜秀區牧草種植地,盆栽試驗在安慶師范大學多功能溫室中開展。選擇農田施用量最大的尿素為氮源[13],制成水溶劑后使用。設置3個用量梯度,分別為高用量(100 kg/hm2)、低用量(50 kg/hm2)和不施加氮素的對照(CK,0 kg/hm2)。每個處理5個重復,共30盆。種子破除休眠后,選取顆粒飽滿的種子,均勻播入直徑20 cm的花盆,覆土澆水。待幼苗生長10 d后定苗(每盆12株),并開始氮施加處理,每2 d用噴壺給每盆植物施加100 ml氮素溶液,對照施加等量水。試驗于2018年5月11日開始,35 d后取樣測定相關指標。
1.2植株生長特征的測定
每盆隨機選取6株植物,先測定植株高度,再將地上部和地下根系分開稱鮮質量(電子天平精度為0.000 1 g)。隨后在每株上選取3枚健康葉片,用直尺、電子游標卡尺等工具測量葉長、葉寬、葉厚,計算葉長寬比。將每盆6株植物地上部分和地下部分分別混合,放入75 ℃烘箱中烘24 h,再稱其干質量。用公式計算地上部含水量和地下部含水量,含水量=[(鮮質量-干質量)/干質量]×100%。再以此計算含水量的根冠比[(根含水量/地上含水量)×100%]和生物量的根冠比[(根干質量/地上部干質量)×100%]。
1.3光合色素含量的測定
每盆選取6株植株,每株選擇3枚健康葉片,共18枚葉片作為1個重復。葉片剪成條狀,用電子天平(精度為0.000 1 g)稱取0.2 g樣品,采用95%乙醇法萃取光合色素。用刻度試管定容至20 ml,在黑暗條件下靜置24 h,期間搖晃2次。利用分光光度法測定上清液在波長為645 nm、663 nm及470 nm處的吸光值,根據Lichtenthaler方法計算葉綠素a、葉綠素b、類胡蘿卜素含量及總葉綠素含量[19],并計算葉綠素a/b值。
1.4數據處理
采用單因素方差分析對比葉長、葉寬、葉厚、葉長寬比、葉綠素a含量、葉綠素b含量、類胡蘿卜素含量、葉綠素a含量與葉綠素b之比、地上部含水量、地下部含水量、含水量根冠比及生物量根冠比在不同氮添加量處理間的差異性,利用Levenes test進行方差分析,使用Duncans法進行多重比較。用Pearson相關性分析檢驗氮施加影響下2種豆科植物功能性狀之間的相關性。用主成分分析(PCA)解析不同功能性狀的協同變化關系[18,20]。單因素方差分析、Pearson相關性分析以及主成分分析采用SPSS 20.0(SPSS Inc. USA)軟件完成,常規數據分析采用Excel 2016完成。
2結果與分析
2.12種豆科牧草形態性狀對不同用量氮的響應
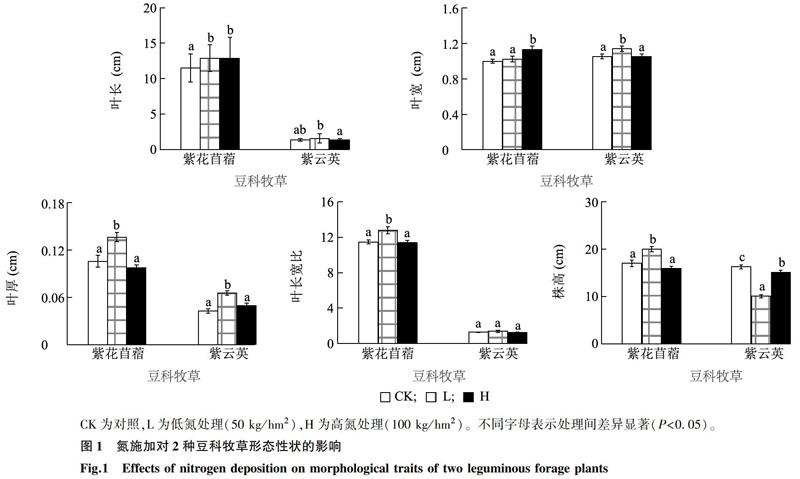
不同氮用量處理下,2種豆科植物形態特征具有不同程度的響應趨勢(圖1)。隨氮施加量的增加,紫花苜蓿的葉長和葉寬也顯著增大(P<0.05),即H(高氮組)>L(低氮組)>CK(對照組);而株高、葉厚及葉長寬比則呈先增后降趨勢,表明出現了高用量抑制的現象。對紫云英而言,其葉長、葉寬及葉厚也呈現先增后降的趨勢,但其葉長寬比在不同施氮用量下無顯著差異。此外,紫云英株高在氮施加條件下顯著降低,其中低用量下株高降低最明顯,表明氮施加后紫云英以橫向生長為主。
2.22種豆科植物含水量與生物量分配對不同用量氮的響應
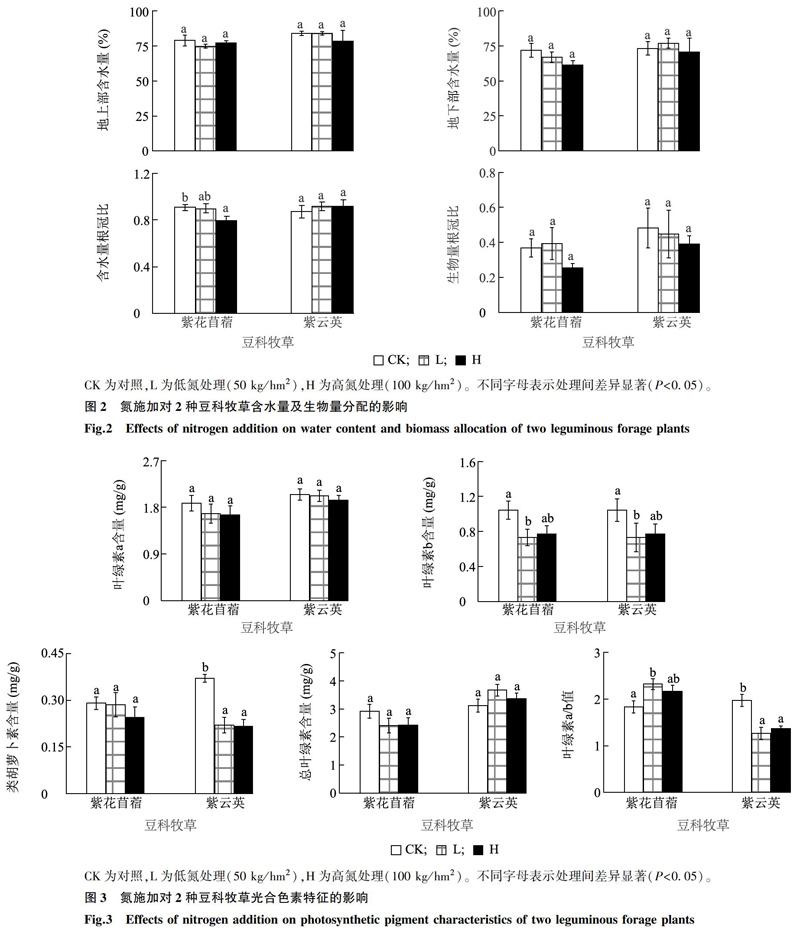
隨施氮量的增加,紫花苜蓿和紫云英的地上部含水量、地下部含水量及生物量根冠比均無顯著差異(圖2),但紫花苜蓿含水量根冠比在高氮下顯著低于對照組(P<0.05),表明施氮沒有改變2種豆科植物生物量分配,但抑制了紫花苜蓿根的水分吸收和利用。
2.32種豆科植物光合色素特征對不同用量氮的響應
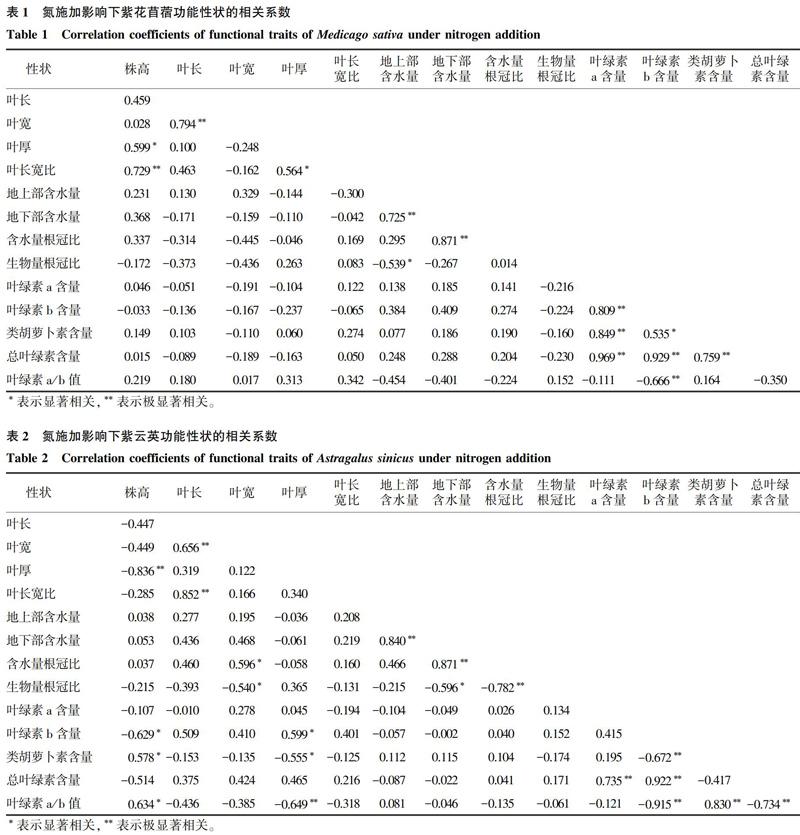
隨著施氮量增加,紫花苜蓿葉綠素a、類胡蘿卜素含量和總葉綠素含量變化均不顯著(圖3);其葉綠素b含量表現為CK>H>L,即施氮顯著降低了葉綠素b含量;對照組紫花苜蓿葉綠素a/b值顯著低于低氮處理組(P<0.05)。對紫云英而言,其葉綠素a和總葉綠素含量在處理間均無顯著差異,但其葉綠素b、類胡蘿卜素、葉綠素a/b值均表現為施氮后顯著降低。
2.4不同施氮量下2種豆科牧草功能性狀的相關性
相關性分析結果表明,紫花苜蓿葉綠素a/b值與葉綠素b含量呈極顯著負相關關系,總葉綠素含量與葉綠素a、葉綠素b、類胡蘿卜素含量呈極顯著正相關關系,類胡蘿卜素含量與葉綠素a和葉綠素b含量分別呈極顯著和顯著正相關關系(表1)。生物量根冠比與地上部含水量呈顯著負相關關系,含水量根冠比與地下部含水量之間及地下部含水量與地上部含水量之間均極顯著正相關關系。葉長寬比與株高和葉厚分別呈極顯著和顯著正相關關系,葉寬與葉長也呈極顯著正相關關系。
紫云英葉綠素a/b值與株高、類胡蘿卜素含量分別呈顯著和極顯著正相關關系,與葉厚、葉綠素b含量以及總葉綠素含量呈極顯著負相關關系(表2)。葉綠素b含量與葉厚呈顯著正相關關系,總葉綠素含量與葉綠素a、葉綠素b含量呈極顯著正相關關系,類胡蘿卜素含量與葉綠素b含量呈極顯著負相關關系。生物量根冠比與葉寬、地下部含水量及含水量根冠比呈顯著或極顯著負相關關系。含水量根冠比與葉寬和地下含水量分別呈顯著和極顯著正相關關系。葉厚與株高呈極顯著負相關關系,葉長與葉寬呈極顯著正相關關系。
2.5不同施氮量下2種豆科牧草功能性狀的權衡特征
由表3可知,在紫花苜蓿PCA第1主成分中,地上部含水量、地下部含水量、葉綠素a含量、葉綠素b含量、類胡蘿卜素含量和總葉綠素含量的變化具有協同性(特征值的絕對值>0.5)。在第2主成分中,株高、葉厚、葉長寬比和葉綠素a/b值有一定的協同性,葉長、葉寬、生物量根冠比和含水量根冠比與其他性狀并沒有明顯的協同變化趨勢。
對紫云英功能性狀的PCA分析結果表明,第1主成分中株高、葉長、葉寬、葉厚和葉長寬比、葉綠素b含量、類胡蘿卜素含量、總葉綠素含量和葉綠素a/b值表現出較強的協變趨勢,但株高、類胡蘿卜素含量及葉綠素a/b值與其他性狀的協變方向相反。在第2主成分中,葉寬、地上部含水量、地下部含水量、生物量根冠比、含水量根冠比具有協同變化特征,但生物量根冠比與其他性狀的協變方向相反。此外,紫云英葉綠素a含量與其他性狀均無明顯的協變性。
由此可見,2種豆科牧草形態、生物量分配、含水量及葉片光合色素等功能性狀對氮施加的協變趨勢有較大差異,體現了明顯不同的權衡特征。
3討論
增加土壤養分會改變植物多方面的適應對策,其中形態學適應是一種最基本的機制[21]。氮添加使土壤可利用性氮增加,在一定程度上會促進植物生長。本研究結果顯示,低氮處理組紫花苜蓿的株高顯著高于其他2組,這與李雙娥等[22]的研究結果一致,即適量的氮促進了海南紅豆(Ormosia pinnata)與馬占相思(Acacia mangium)地上部分的累積,過量氮反而抑制植物地上部分的生長[4,10]。然而紫云英的株高為對照組>高氮處理組>低氮處理組,可能是由于紫云英植株高度對氮的響應更加敏感,且出現了橫向(匍匐)生長。紫花苜蓿葉長與葉寬表現為高氮處理組>低氮處理組>對照組,表明氮對葉片的生長有明顯的促進作用[23]。而紫云英葉長與葉寬隨著氮用量的增加先增后降,這與大豆葉片面積響應氮添加的結果相一致[24]。紫花苜蓿的葉厚與葉長寬比和紫云英的葉厚隨著氮用量的增加先增后降,是典型的低促高抑現象。
植株體內不同器官的有機物分配往往會根據外界環境的變化而改變,以提高自身對環境的適應能力[10]。根系是豆科植物生長的重要結構和物質基礎。本研究中紫云英的根冠比順序為對照組>低氮處理組>高氮處理組,即施氮降低了植物根冠比,這與木荷(Schima superba)等木本植物響應氮添加的結果一致[25];同時,該現象也與荒漠草本植物對氮的響應相符,即植物在氮施加后,地上部分的生長比地下根系快,進而導致根冠比降低[26]。這可能是因為對照組植物受到了氮的限制,進而分配給地下根系更多生物量以吸收營養,保證植物正常營養供應,且這種限制會在氮施加后逐漸解除[10]。紫花苜蓿的根冠比順序為低氮處理組>對照組>高氮處理組,即根冠比先增后降,其中高量氮添加下根冠比下降趨勢更明顯,這也符合氮添加后植物根冠比下降的普遍規律[4,10]。但一些研究結果表明,在較低的施氮水平下,施氮增加了紫花苜蓿的地下生物量[27],說明植物對體內生物量分配比例的調控也受到氮施用量的影響,低用量促進而高用量抑制。兩種植物含水量的變化并不顯著,表明氮用量對植物含水量的影響有限。
植物葉片是光合作用的主要器官,光合色素含量是衡量光合作用強弱的重要指標,環境因子的變化會影響植物光合色素含量[28-34]。本研究中對照組紫云英類胡蘿卜素含量大于施氮組,表明氮對紫云英類胡蘿卜素的合成有抑制作用。葉綠素a是光合作用中心色素分子,具有把光能轉換為電能,進行電子傳遞并最終轉化成化學能的功能,而葉綠素b具有吸收光能并傳遞光能的能力[32]。本研究中,紫云英和紫花苜蓿的葉綠素b含量大小順序為對照組>高氮處理組>低氮處理組,而葉綠素a變化不顯著,這可能是由于低氮促進葉綠素b的合成,而高氮抑制其合成。葉綠素a在遮光條件下會加速向葉綠素b轉化,使葉綠素a/b值降低,有利于提高植物對短波藍紫光(弱光)的利用率。而葉綠素a/b值升高則表明,植物在強光下光合速率會提高,能明顯抵抗光抑制[32,35]。在本研究中,紫花苜蓿葉綠素a/b值低氮處理組顯著大于對照組,而紫云英為對照組大于施氮處理組,表明施氮后紫花苜蓿對強光的利用率提高,而紫云英則對弱光的利用率較高。
紫花苜蓿和紫云英株高、葉寬、含水量根冠比、類胡蘿卜素含量、葉綠素a/b值對不同氮用量的響應表現出較為明顯的差異性,這可能是由種間差異性導致的,即不同物種對氮添加的響應不同。厘清這種多功能性狀的差異性,就需要進行性狀整合分析。研究結果表明,紫花苜蓿和紫云英植株功能性狀之間具有一定相關性,也表現出一定的協變特征,尤其在葉片光合色素方面。表型協變是植物性狀之間互相作用而協同變化的現象[36],它可以影響功能性狀間的變化而適應環境的變化,從而形成功能性狀間的最佳組合[18,37],是物種進化的重要標志。紫花苜蓿的含水量特征(地上含水量、地下含水量)與光合性狀(葉綠素a、葉綠素b、類胡蘿卜素、總葉綠素含量)之間表現出較強的整合格局,反映了紫花苜蓿的含水量和光合色素特征之間具有適應性的協同作用,是氮施加影響過程中植物體發育性狀間相互權衡的結果。紫云英的形態性狀(株高、葉長、葉寬、葉厚和葉長寬比)和光合生理性狀(葉綠素b含量、類胡蘿卜素含量、總葉綠素含量和葉綠素a/b值)之間也有較強的協變趨勢,其中株高、葉綠素a/b值和類胡蘿卜素含量與其他性狀具有反向協變性,說明紫花苜蓿地上部分功能性狀具有明顯的整體性;紫云英生物量根冠比與其他性狀的協變趨勢相反,這表明紫云英在氮施加下會通過形態結構和生理特征響應,優先調配資源給地上部分,形成地上部分和地下部分的不一致性,也即產生了地上部分和地下部分的權衡。
4結論
不同量氮施加對紫花苜蓿形態性狀、含水量根冠比、葉綠素b含量、葉綠素a/b值和紫云英的形態性狀及光合色素特征均具有顯著影響。兩物種間部分功能性狀(如葉厚、葉綠素a含量、葉綠素b含量、地上地下含水量等)對氮添加的響應具有相同趨勢,但大部分功能性狀存在種間差異性。不僅如此,2種物種功能性狀對氮素施加的協同適應性也存在種間差異。研究證實2種不同豆科牧草對外源氮添加具有差異化的響應特征和性狀協變趨勢,其機理需進一步研究。
參考文獻:
[1]BOBBINK R, HICKS K, GALLOWAY J, et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis[J]. Ecological Applications, 2010, 20(1):30-59.
[2]毛晉花,邢亞娟,馬宏宇,等. 氮沉降對植物生長的影響研究進展[J]. 中國農學通報, 2017, 33(29):42-48.
[3]劉學軍,張福鎖. 環境養分及其在生態系統養分資源管理中的作用: 以大氣氮沉降為例[J]. 干旱區研究, 2009, 26(3):306-311.
[4]常運華,劉學軍,李凱輝,等. 大氣氮沉降研究進展[J]. 干旱區研究, 2012, 29(6):972-979.
[5]張維,趙文勤,謝雙全,等. 模擬降水和氮沉降對準噶爾盆地南緣梭梭光合生理的影響[J]. 中國生態農業學報, 2018, 26(1):106-115.
[6]肖迪,王曉潔,張凱,等. 模擬氮沉降對五角楓幼苗生長的影響[J]. 北京林業大學學報, 2015, 37(10):50-57.
[7]CLARK C M, TILMAN D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands[J]. Nature, 2008, 451(7179):712-715.
[8]張燕,崔學民,樊明壽. 大氣氮沉降及其對草地生物多樣性的影響[J]. 草業科學, 2007, 24(7):12-17.
[9]蘇潔瓊,李新榮,回嶸,等. 氮沉降對荒漠化草原草本植物物種多樣性和群落組成的影響[J]. 西北植物學報, 2012, 32(4):795-801.
[10]毛晉花,邢亞娟,閆國永,等. 陸生植物生物量分配對模擬氮沉降響應的 Meta 分析[J]. 生態學報,2018,38(9):3183-3194.
[11]萬宏偉,楊陽,白世勤,等. 羊草草原群落6種植物葉片功能特性對氮素添加的響應[J]. 植物生態學報, 2008, 32(3):611-621.
[12]趙葉舟,王浩銘,汪自強. 豆科植物和根瘤菌在生態環境中的地位和作用[J]. 農業環境與發展, 2013, 30(4):7-12.
[13]李雪芬. 氮添加對紫花苜蓿生長特性及光合生理特性的影響[J]. 北方園藝, 2015(2):158-164.
[14]劉曉娟,馬克平. 植物功能性狀研究進展[J]. 中國科學: 生命科學, 2015, 45:325-339.
[15]何念鵬,劉聰聰,張佳慧,等. 植物性狀研究的機遇與挑戰: 從器官到群落[J]. 生態學報, 2018, 38(19): 6787-6796.
[16]LAVOREL S, GRIGULISK, LAMARQUE P, et al. Using plant functional traits to understand the landscape distribution of multiple ecosystem services[J]. Journal of Ecology, 2011, 99:135-147.
[17]LIENIN P, KLEYER M. Plant trait responses to the environment and effects on ecosystem properties[J]. Basic and Applied Ecology, 2012, 13(4):301-311.
[18]邱東,張靜,吳楠,等. 安徽大龍山國家森林公園東亞小金發蘚配子體與孢子體個體性狀及其相互關系[J]. 植物研究, 2019, 39(6):835-845.
[19]LICHTENTHALER H K, WELLBURN A R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents[J] . Biochemical Society Transactions, 1983, 11:591-592.
[20]CONCHITA A, CARLOS M H. Patterns made of patterns: Variation and Covariation of leaf nutrient concentration and between populations of Prunus mahaleb[J]. New Phytologist, 2001, 150:629-640.
[21]PATTERSON T B, GUY R D, DANG Q L. Whole-plant nitrogen- and water-relations traits, and their associated trade-offs, in adjacent muskeg and upland boreal spruce species[J]. Oecologia, 1997, 110(2):160-168.
[22]李雙娥,李義勇,方熊,等. 不同氮添加和添加方式對南亞熱帶4個主要樹種幼苗的生長的影響[J]. 植物生態學報, 2019, 39(10):950-961.
[23]武新紅,彭克勤,梁宗鎖,等. 氮磷對菘藍營養生長期根、冠生長的影響[J]. 西北農業學報, 2008, 17(4):274-278.
[24]宋英博. 施氮對大豆葉片生理指標的影響[J]. 中國農學通報, 2010, 26(15):243-246.
[25]李明月,王健,王振興,等. 模擬氮沉降條件下木荷幼苗光合特性、生物量與C、N、P分配格局[J]. 生態學報, 2013, 33(5):1569-1577.
[26]周曉兵,張元明,王莎莎,等. 3種荒漠植物幼苗生長和光合生理對氮增加的響應[J]. 中國沙漠, 2011, 31(1):82-89.
[27]ESECHIE H A, AL-BARHI B, AL-GHEITY S, et al. Root and shoot growth in salinity-stressed alfalfa in response to nitrogen source[J]. Journal of Plant Nutrition, 2007, 25(11):2559-2569.
[28]袁孟玲,岳堃,王紅,等. 增強UV-B輻射對芒果成年樹光合作用及其產量與常規品質的影響[J].南方農業學報,2018,49(5):930-937.
[29]高祺,李明,樸琳,等. 拔節期弱光和漬水脅迫對春玉米光合作用、根系生長及產量的影響[J].江蘇農業學報,2018,34(6):1276-1286.
[30]郝正剛,王志恒,魏玉清,等.外源鈣鎘處理對甜高粱幼苗葉片光合作用的影響[J].江蘇農業科學,2019,47(19):75-80.
[31]DE E, DONG D, ZENG X, et al. Direct effect of acid rain on leaf chlorophyll content of terrestrial plants in China[J]. Science of the Total Environment, 2017, 605/606:7-769.
[32]李合生. 現代植物生理學[M]. 3版. 北京: 高等教育出版社, 2012:100-140.
[33]李君,婁運生,馬莉,等. 夜間增溫和水分管理耦合對水稻葉片光合作用和熒光特性的影響[J].江蘇農業學報, 2019, 35(3):506-513.
[34]王銳潔,劉筱,楊淑君,等. 氮沉降背景下遮陰對虎耳草生長和光合作用的影響[J].南方農業學報,2019, 50(2):330-337.
[35]占吉成,王利軍,黃衛東. 弱光環境下葡萄葉片的生長及其在強光下的光合特性[J]. 中國農業大學學報, 2004, 7(3):75-78.
[36]KLINGENBER G, PETER C. Morphological integration and developmental modularity[J]. Annual Review of Evolution and Systematics, 2008, 39(1):115-132.
[37]施宇,溫仲明,龔時慧. 黃土丘陵區植物葉片與細根功能性狀關系及其變化[J]. 生態學報, 2011, 31(22):6805-6814.
(責任編輯:張震林)
