中華鱘的性腺發育與退化問題研究
2020-12-10 03:55:18張富鐵劉煥章
水生生物學報 2020年6期
高 欣 張富鐵 常 濤 劉煥章
(中國科學院水生生物多樣性與保護重點實驗室, 中國科學院水生生物研究所, 武漢 430072)
中華鱘(Acipenser sinensis)是一種低等的硬骨魚類, 是國家一級重點保護野生動物。中華鱘被認為是長江水生動物保護的旗艦物種和長江水生態系統健康狀況的指示物種。近年來, 中華鱘多次被監測到未進行野外繁殖, 其生存前景堪憂。
有關中華鱘沒有繁殖的原因存在多方面的爭議, 包括三峽工程調度導致水文條件改變, 不能滿足中華鱘繁殖的需求; 以及中華鱘繁殖親本太少,難以自然配對繁殖, 等等。最新奇的一個論點是黃真理和王魯海[1]提出的:在葛洲壩水利樞紐修建以后, 中華鱘的性腺退化嚴重, 是導致中華鱘種群數量減少的主要原因, 并宣稱“中科院水生所有關否定中華鱘性腺退化的方法和材料, 是不充分和不可靠的”。這是一個關乎中華鱘保護的嚴重問題, 如果沒有正確的源流認識, 任由謬論誤導, 將極大地影響中華鱘未來的保護工作。
經過仔細研讀中國科學院水生生物研究所(以下簡稱中科院水生所)的相關文獻, 并分析黃真理等的系列文章就可以發現, 黃真理等沒有親自在長江對中華鱘洄游和繁殖實地調查的第一手資料和數據, 而是選擇性地引用他人文章中的數據, 然后發揮想象力, 編造故事, 自我陶醉。而中科院水生所的研究結果是通過長期實地調查取得的, 具有客觀性和科學性, 經得起推敲和質疑。本文試從中華鱘的性腺發育過程及退化的判斷標準、中華鱘的生活史階段及性腺狀態、葛洲壩與三峽工程對中華鱘繁殖影響等方面, 論述中華鱘的性腺是否可以發育成熟? 是否存在性腺退化現象? 從而為中華鱘的保護提供科學合理的依據。
1 魚類性腺的發育與退化過程
1.1 魚類性腺的發育與退化過程
魚類的生殖系統包括性腺(精巢、卵巢)和生殖導管(輸精管、輸卵管)。魚類的生殖細胞可以分為6個發育時相, 對應地魚類的性腺發育可以劃分為6個時期, 即Ⅰ—Ⅵ期。其中Ⅰ—Ⅲ期為發育不成熟的性腺,Ⅳ—Ⅴ期為成熟性腺。對于一生多次產卵的魚類來說, 性腺發育到Ⅳ期成熟, 可以產卵; 產卵過程中為Ⅴ期性腺, 這一時期極為短暫; 產卵完成后為Ⅵ期性腺。在一般情況下, 產卵后會慢慢恢復到Ⅱ期性腺,然后再發育成熟, 進入新的繁殖循環[2—5]。
性腺退化(Gonadal degeneration)現在一般是指性腺成熟(性腺Ⅳ期), 但因環境條件無法滿足繁殖需求導致不產卵(排精)或者部分產卵(排精), 從而錯過繁殖窗口期后性腺再被吸收的過程。也有研究將完全產卵和排精后, 性腺(Ⅵ期)回復到Ⅱ期的過程視為性腺退化。其特征是性腺逐漸萎縮、營養物質被吸收, 性腺會從Ⅳ或者Ⅵ期回到Ⅱ期[3—7]。性腺成熟與退化可以通過形態學、組織學觀察和血清學檢查來進行判斷。
1.2 中華鱘“性腺退化”的由來
早在1971—1982年開展的葛洲壩工程“救魚問題”論證過程中, 就有人“預測”中華鱘受到葛洲壩阻隔, 不能溯游到長江上游的產卵場, 性腺會退化。柯福恩等[8]首次“證實”了葛洲壩下中華鱘大規模性腺退化的問題。他們認為, 在葛洲壩工程修建后, 下游環境條件發生變化, 造成了大量的中華鱘出現性腺退化現象。他們分析了1984年4—5月和10—11月在葛洲壩下宜昌江段采集的中華鱘樣本,認為存在很高比例的性腺退化, 性腺退化個體占繁殖群體個體總數的48.87%, 雄魚性腺退化個體比例為29.41%, 雌魚性腺退化個體比例達71.43%。特別是在10—11月的樣本中, 除1尾雌魚外其余9尾均存在性腺退化現象。
柯福恩等[8]定義中華鱘性腺退化的形態學標準為“雄鱘: 性腺上脂肪重比正常脂肪重的平均值減少50%以上, 精巢重比正常精巢重平均值減少或者增加10%以下; 或雄魚性腺上的脂肪全部消耗完, 精巢重不超過1.5 kg的性腺為退化性腺。雌鱘: 性腺上脂肪基本消失或剩下很少, 卵徑沒有增大, 卵粒變色、變形, 成熟系數在10以下的卵巢為退化卵巢。”
我們認為, 以性腺上的脂肪多少作為標準判斷中華鱘性腺是否退化是不合適的。研究發現, 對于西伯利亞鱘或者俄羅斯鱘的雄魚, Ⅱ期精巢完全包埋在脂肪中, 性腺上脂肪約為性腺重量的80%—95%; 精巢發育至Ⅲ期時, 脂肪大量減少。雌鱘到Ⅱ期末, 卵巢也被大量脂肪包圍; Ⅲ期時濾泡增長,卵膜層加厚, 性腺變長變大, 卵黃沉積, 脂肪會大量減少, 白色的、灰色的卵母細胞會增加。到IV期時, 無論雌雄鱘魚的性腺上幾乎都完全看不到脂肪[7]。同樣, 中華鱘IV期時, 性腺脂肪已消失或僅有少量絲絡狀的脂肪組織[9, 10]。因此脂肪的減少、尤其Ⅲ期性腺發育過程中脂肪大量減少是鱘魚正常發育過程中必然出現的現象。以此將其作為判斷性腺退化的標準, 容易誤將正常發育的中華鱘Ⅲ期性腺判斷為退化性腺。
此外, 以“卵徑沒有增大, 卵粒變色、變形, 雌鱘性腺成熟系數低于10的即為退化”, 判斷也不嚴謹。因為正常生長Ⅲ期雌鱘的卵徑是較小的(中華鱘為2.0—3.7 mm, 俄羅斯鱘或者西伯利亞鱘為1.6—2.6 mm), 卵母細胞呈現多種顏色(黃、白、灰等), 卵巢中有卵原細胞和Ⅰ—Ⅲ時相的卵母細胞同時存在, 大小和形態各異, 此時中華鱘性腺成熟系數為3.2—13.6[7, 9, 10]。因此, 以此作為性腺退化的判斷依據, 也極易將正常發育的Ⅲ期卵巢誤判為退化性腺。
柯福恩等[8]觀察到所謂退化中華鱘雌鱘“卵粒沒有光澤, 呈灰白色、黃白色、淺褐色且帶白色斑紋, 尤如麻雀蛋殼的花紋, 有的呈糊狀花紋”, “卵粒的卵徑波動在 2.1—3.3 mm, 平均2.75 mm, 成熟系數波動在4.91—9.68, 平均7.62。同一卵巢有的卵粒大小不均, 20尾魚的卵巢都伴隨白色點狀的Ⅱ時相卵粒”, “大多數都是處在第Ⅲ時相的卵母細胞, 但也有第Ⅳ時相的卵母細胞”, “卵膜(放射帶和膠質膜)加厚, 其厚度可增至42.19 μm”。實際上, 這些卵母細胞的大小、顏色、時相及卵膜結構等, 都比較符合雌鱘正常發育Ⅲ—Ⅳ期性腺的特征, 而不是性腺退化。
周春生等[9]和四川省長江水產資源調查組[10]詳細記載了中華鱘性周期中已產卵性腺退化的特征:Ⅵ期時, 性腺松弛充血, 絕大部分成熟卵已排出, 殘存在卵巢中的主要是結締組織和排卵后脫落的濾泡膜。有少量未產出的過熟卵已處于退化過程中,這些卵粒已失去彈性, 體積膨大, 卵徑可達8 mm,形狀為不規則的球形, 呈灰褐色, 卵核和卵黃粒溶解或僅存少量卵黃粒, 卵膜變形, 厚度不均, 卵膜很易破裂。卵巢內除少量第Ⅳ時相卵母細胞外, 主要為第Ⅱ時相的卵母細胞, 黃白色, 數量較多, 卵徑0.3—0.9 mm, 成熟系數2.5—5。Chebanov和Galich[7]也記載: Ⅵ期時, 性腺強烈充血, 結構松散, 開始時有明顯的邊界和膜層。但是, 如果錯過了產卵期,性腺會出現再吸收, 隨著再吸收過程的進行, 成熟卵母細胞界限越來越模糊; 成熟的雌鱘在繁殖溫度下超過20d就會出現過熟卵子和濾泡上皮細胞的再吸收現象, 成熟卵母細胞形狀變得不規則, 邊界消失, 初級卵母細胞越來越清晰可見, 在某些情況下性腺會萎縮。柯福恩等[8]未報道中華鱘性腺有無充血情況, 所謂的退化性腺中卵母細胞時相也不符合。從鱘魚類性腺發育的規律看, 柯福恩等[8]所謂判斷為退化的性腺, 實際上是中華鱘正常的未發育成熟的性腺。
從1984年起, 柯福恩等就以人工繁殖的名義,申請捕撈中華鱘的親魚, 在20世紀90年代每年捕撈數十尾[11]。試問, 既然這些中華鱘的性腺已經“退化”, 那他們捕去干什么用呢?
2 中華鱘的生活史階段及其在長江中的性腺發育過程
2.1 中華鱘在長江中的生活史階段
中華鱘是一種洄游魚類, 具有溯河產卵習性。中華鱘產卵場位于長江上游、金沙江下游, 其繁殖期為秋季10—11月。在產卵場孵化的中華鱘仔魚于長江中生長和發育, 向下游洄游, 于次年4—5月到達長江口, 然后進入中國近海的大陸架水域, 進一步攝食和生長。在一般情況下, 中華鱘雌鱘13—26齡到達性成熟, 雄鱘8—18齡到達性成熟[10—12]。
每年6—7月, 在海中長大、即將成熟的中華鱘個體進入長江口, 溯江而上; 8—9月經過九江; 9月下旬至10月陸續進入湖北江段; 10—11月份, 上溯至四川, 并在產卵場下游附近江段的深潭或坑洼中越冬, 在翌年秋季10—11月再進入產卵場進行繁殖。由于中華鱘進入長江后, 到完成其繁殖全過程需要15個月以上的時間, 因此, 每年的6—11月會在長江不同江段發現處于不同洄游階段的兩類中華鱘繁殖群體。其中在湖北及以下江段發現的中華鱘群體, 通常是當年進入長江等待翌年繁殖的亞成熟個體, 即新股群; 而在上游江段發現的中華鱘群體則同時包括當年進入長江的亞成熟個體(新股群), 和上年進入長江準備當年繁殖的成熟個體(老股群)[10]。根據歷史上合江至屏山江段的捕撈情況來看, 每年10—11月繁殖季節捕獲的主要是性腺發育成熟的老股群, 而性腺處于Ⅲ期的亞成熟個體主要出現在12月以后。隨著葛洲壩工程的修建, 阻隔了中華鱘的洄游路線, 這也導致中華鱘在壩下大量聚集, 并同時存在新老兩個股群, 而新股群的越冬場也由四川江段下移至長江中游江段。
產后親鱘的洄游過程在不同產卵場存在差異。四川省長江水產資源調查組在長江上游原產卵場的調查認為, 產后親鱘大部分迅速降河返回海洋育肥; 而在葛洲壩產卵場, 王成友[13]對4尾精巢Ⅳ期的雄鱘標記放流發現, 其繁殖后并不立即離開產卵場, 而是在產卵場徘徊最長達148d, 這意味著雄鱘從進入產卵場到離開約經歷20個月, 而雌鱘則是立即離開。相關研究顯示, 中華鱘降河洄游的速度較快, 只需花費約15d的時間就可完成整個遷移過程到達海中[14]。
2.2 中華鱘的性腺發育狀態
研究表明, 中華鱘成魚從第一年的夏季進入長江后, 到次年的秋季進行繁殖, 期間是停食的, 其性腺發育所需的能量消耗完全依靠身體積累的脂肪來維持[10]。其在不同的生活史階段, 性腺發育狀態不同; 在繁殖期后, 由于能量消耗極大, 雌鱘需要迅速返回海洋進行攝食。
在一般情況下, 每年進入長江的中華鱘性腺通常處于Ⅱ—Ⅲ期, 其中卵粒在不同個體中呈現黃色、褐色和灰色。灰褐色卵徑較大, 平均2.45—3.07 mm, 黃色卵徑略小, 平均2.19—2.62 mm。性成熟系數為3.2—13.6。卵粒包含在黃色脂肪內, 漁民稱這樣的卵巢為“油子”。而精巢呈乳白色, 性成熟系數較小, 為0.15—0.64。調查發現, Ⅲ期的鱘魚通常6—9月份在河口、江蘇、安徽、九江、湖北等江段中出現, 均為新股群。而9—11月在上游江段發現的鱘魚中, 大部分個體卵巢處于Ⅳ—Ⅴ期,為上年進入長江的老股群。此時卵巢內卵徑在3.71—4.90 mm, 多數為4.5 mm, 性成熟系數為14.8—19.5, 性腺中脂肪很少或無脂肪; 漁民稱該時期的卵巢為“水子”。而精巢擠壓或切割時有白色液體滲出, 性成熟系數在0.62—0.67變動[10]。
據相關研究報道, 沙市和宜昌江段曾分別在1979年和1980年的5月捕獲到少量性腺處于Ⅳ期的“水子”鱘魚[15], 說明這部分鱘魚為上年進入長江,準備當年上溯參與繁殖的群體。由此可以推斷, 在葛洲壩阻隔前, 中華鱘也能在湖北江段達到性腺成熟。
綜上所述, 處于不同生活史階段的中華鱘, 或者說中華鱘的新股群和老股群, 其性腺發育狀態是不同的。據此分析, 柯福恩等[8]1984年4月采集的中華鱘是1983年從長江口上溯的待產親魚, 應該是老股群, 其將在1984年10—11月參加繁殖。4月取樣時正是性腺發育時期, 卵粒直徑2.9—3.3 mm, 成熟系數6.49—13.2(柯福恩等[8]表 2), “大多數都是處在第Ⅲ時相的卵母細胞, 但也有第IV時相的卵母細胞”(柯福恩等[8]性腺切片描述原文), 這表明這些卵巢符合正常發育卵巢的特征(Ⅲ—Ⅳ期), 并不是退化卵巢。
據危起偉等[14]的資料, 1984年中華鱘自然繁殖兩次, 分別是10月16日和11月13日。那么柯福恩等[8]1984年10月19—31日采集的雌鱘樣本中, 理論上包括三個類型的群體: 1983年上溯且已在1984年10月16日產卵的魚(老股群1)、1983年上溯且即將于1984年11月13日產卵的魚(老股群2), 及1984年上溯將在1985年秋季產卵的魚(新股群)。老股群1鱘魚已完成正常的生殖活動, 繁殖后性腺吸收會從Ⅵ期慢慢回復到Ⅱ期, 符合性周期規律。老股群2鱘魚性腺達到成熟即將參與11月的繁殖, 柯福恩等[8]表2中性成熟系數達到23.33的那尾魚屬于老股群2。新股群鱘魚則具有正常發育的尚未成熟的Ⅲ期卵巢。結合中華鱘雌魚產卵后幾小時內立即降河洄游的特點[14], 我們推斷1984年10月16日產卵的雌鱘(老股群1)在10月19—31日不可能出現在產卵場附近。10月采集的10尾雌魚樣本中, 成熟系數達到23.33的雌魚屬于老股群2; 其他的個體推測是未發育成熟、將在1985年秋季產卵的魚(新股群)。因此1984年10月的樣本中也不存在性腺退化現象。
3 葛洲壩水利樞紐對中華鱘性腺發育的影響
3.1 葛洲壩水利樞紐對中華鱘性腺發育時間的影響
1963—1975年, 葛洲壩水利樞紐工程截流前,金沙江三塊石、偏巖子、金堆子和長江上游鐵爐灘、望龍磧等產卵場中華鱘第一次產卵的日期從10月11日至11月1日[10]。葛洲壩截流之后, 1983—2002年, 中華鱘第一次產卵的日期從10月13日至11月7日(圖 1)[12, 16]。從時間范圍來看, 沒有明顯的差異。
為了定量比較, 將10月1日假設為繁殖期的第一天, 以此量化中華鱘初次繁殖時間。葛洲壩工程截流前, 1963—1975年, 中華鱘繁殖時間為11—32d,中值為14d, 平均17d(SD=7d)。葛洲壩截流后, 1983—2002年, 中華鱘繁殖時間為13—38d, 中值為21d, 平均22d(SD=6d)。根據t檢驗分析, 葛洲壩截流前后,中華鱘在長江上游和中游的繁殖時間沒有顯著的差異(P>0.05)。結果表明, 葛洲壩對中華鱘的繁殖時間沒有明顯的影響。這也就意味著葛洲壩修建之后, 宜昌江段滿足中華鱘的性腺發育條件和繁殖條件。洄游距離的改變沒有影響中華鱘性腺發育與產卵繁殖的時間格局。這也說明Huang和Wang[17]依據洄游距離的模型預測完全脫離實際。
3.2 葛洲壩工程對中華鱘性腺發育成熟比例的影響
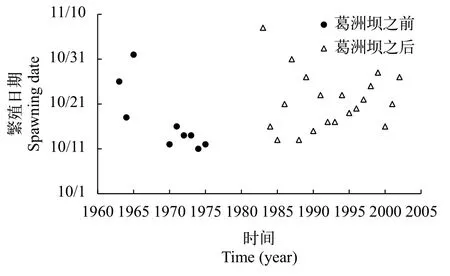
圖 1 1963—2002年長江中華鱘的初次繁殖日期變化(此圖用的是金沙江三塊石、偏巖子、金堆子和長江上游鐵爐灘、望龍磧等產卵場中華鱘第一次產卵的日期) [10, 12, 16]Fig. 1 Changes on spawning date of Chinese sturgeon in the Yangtze River from 1963 to 2002 (The figure shows the first spawning dates of Chinese sturgeon in Sankuaishi, Pianyanzi and Jinduizi sites in Jinsha River and Tielutan and Wanglongqi sites in the upper Yangtze River) [10, 12, 16]
中華鱘親魚需要在長江中滯留1年以上才能完成繁殖活動。長江中會存在當年繁殖的老股群和次年繁殖的新股群, 如果繁殖群體數量保持相對穩定, 在繁殖季節, 正常的性腺成熟個體比例為50%左右[12, 18], 即老股群的個體均可達到性腺成熟。
1981—1998年中華鱘繁殖群體性腺發育成熟個體的平均比例范圍為13.2%—92.6%[12, 18—20], 平均值61.6%(SD=26.8%, 圖 2)。1981年1月4日葛洲壩截流, 1980年6—10月進入長江的新股群, 部分通過葛洲壩到長江上游, 部分在葛洲壩下游發育。因此1981年10—11月的繁殖季節中, 葛洲壩下游繁殖群體性腺發育成熟的個體比例偏少。1982年和1983年性腺成熟個體比例為13.2%和20.5%, 明顯少于50%。這是因為1981年和1982年過度捕撈造成次年性腺成熟個體比例減少[18]。
1983年,我國全面禁止中華鱘的商業捕撈,1984—1990年葛洲壩下游宜昌江段的中華鱘每年捕撈量降為約62—120尾[12]。因此, 1984—1990年,中華鱘性腺成熟個體比例恢復正常, 為37.9%—66.8%,平均53.4%(SD=8.8%); 1991—1998年, 中華鱘性腺成熟個體比例增加至77%—92.6%, 平均86%(SD=5.1%)[12, 20]。這些數據皆證明, 中華鱘繁殖群體中老股群的個體都能達到性腺成熟, 不存在大量性腺退化現象。
1991—1998年的性腺成熟個體比例明顯大于1984—1990年(P<0.05)。性腺成熟個體比例增加的原因是, 葛洲壩水利樞紐修建阻隔了中華鱘的洄游通道, 長江上游產卵場喪失, 中華鱘產卵規模減少。1991年開始, 葛洲壩下游出生個體開始逐漸洄游補充進入繁殖群體, 但是新股群的數量少于老股群的數量, 導致性腺成熟比例增加。
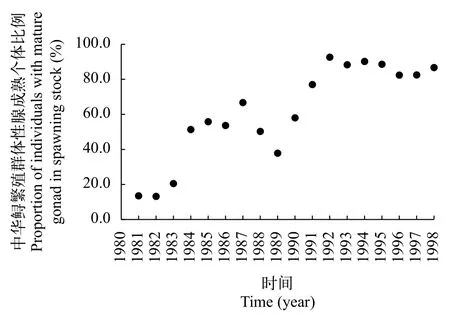
圖 2 1981—1998年長江中華鱘繁殖群體性腺成熟個體比例變化[12]Fig. 2 Proportion changes of individuals with mature gonad in spawning stock of Chinese sturgeon in the Yangtze River from 1981 to 1998 [12]
以野外調查數據[12, 18—20]為依據的分析結果顯示, 嚴重的過度捕撈會降低次年繁殖群體的性腺成熟個體比例; 當捕撈數量降低時, 性腺成熟個體比例會恢復到50%左右; 新股群數量減少會提高性腺成熟個體比例。同時, 結合周春生等[9]、陳金生等[21]研究結果顯示, 在葛洲壩工程修建之后, 在沒有受到過度捕撈影響的情況下, 中華鱘繁殖群體中性腺成熟個體比例是正常的, 沒有所謂嚴重“性腺退化”的現象, 即不影響性腺正常發育成熟。
4 三峽工程對中華鱘繁殖的影響
三峽水利樞紐蓄水運行對中華鱘繁殖活動有明顯的影響(圖 3)。三峽水庫蓄水運行造成中華鱘繁殖期產卵場下游水溫滯后和流量減小[22], 導致中華鱘初次繁殖時間從10月中下旬推遲到11月底, 繁殖次數從每年2次減少到1次(2012年有2次)[16]。2013年、2015年、2017—2019年, 中華鱘在長江中沒有進行繁殖活動。而根據中科院水生所的調查結果顯示, 2017—2019年中華鱘繁殖群體數量已經減少至10—24尾, 難以自然配對繁殖。
5 關于中華鱘性腺退化問題的討論
5.1 柯福恩等對中華鱘性腺退化現象的誤判
柯福恩等[8]提出了中華鱘大規模性腺退化的問題。其研究認為, 在葛洲壩修建以后, 由于產卵環境條件發生改變, 導致中華鱘性腺發生退化,1984年4—5月和10—11月葛洲壩下宜昌江段雌魚性腺退化個體比例為71.43%, 雄魚性腺退化個體比例為29.41%, 性腺退化個體占繁殖群體個體總數的48.87%。
然而仔細分析柯福恩等[8]判斷性腺退化的標準, 無論是以性腺上脂肪的重量, 還是以卵徑的大小和性成熟系數低于10為標準, 都不能準確地判斷是否為性腺退化。正確的判斷必須是經歷IV或者VI期后的性腺, 卵巢充血, 卵母細胞出現再吸收的情況。柯福恩等[8]判斷為性腺退化的雌魚應該是未發育成熟的魚。同時, 柯福恩等[8]沒有提供制定性腺退化標準的理論依據和數據基礎, 也沒有進行數據統計分析來驗證該標準, 因此這些所謂“標準”從理論、數據和統計學等方面都沒有實質的支撐。
中華鱘性腺退化觀點的提出還存在著一定的時代背景, 即20世紀70、80年代, 葛洲壩救魚之爭,國家水產總局力主修建過魚設施, 作為拯救中華鱘等珍貴稀有水生動物的主要保護措施[23, 24]。水利樞紐工程會造成洄游親魚性腺退化的觀點則是支撐修建過魚設施的依據之一[25]。1981年, 張亢西[25]就以俄國鱘和鰉為例, 提出了大壩會造成壩下洄游親魚性腺退化的觀點, 并建議建設“水利樞紐漁業綜合工程”, 例如魚道。因此, 柯福恩等[8]并非提出中華鱘性腺退化的第一人, 而只是“印證”了這一觀點。
5.2 黃真理等[1, 17]、柯福恩等[8]對中華鱘性腺發育過程的錯誤認識
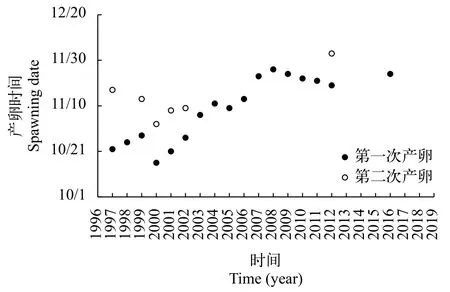
圖 3 1997—2019年三峽工程蓄水前后中華鱘繁殖日期(2014年中華鱘的繁殖時間和地點不確定)Fig. 3 Spawning dates of Chinese sturgeon before and after impoundment of the Three Gorges Reservoir from 1997 to 2019(The spawning date and location are unknown in 2014)
從黃真理等[1, 17]、柯福恩等[8]對中華鱘性腺退化的觀點看, 他們對于中華鱘的性腺發育過程存在若干錯誤的認識。首先, 黃真理和王魯海[1, 17]通過建立性腺發育模型認為, 中華鱘的性腺發育與洄游距離相關; 在葛洲壩工程截流后, 由于洄游距離減少, 所以中華鱘性腺退化。這是十分荒謬的。據柯熏陶[15]報道, 沙市和宜昌江段曾分別在1979年和1980年春季(5月)捕獲到性腺處于Ⅳ期的“水子”鱘魚, 說明在葛洲壩工程阻隔前, 中華鱘就可以在湖北江段達到性腺發育成熟, 與洄游距離無關。在葛洲壩工程阻隔后, 中華鱘在壩下能夠成功繁殖。1984—1998年, 繁殖雌魚個體為年均72尾(26—163尾)或62尾(24—133尾)(常劍波[12]表5.15, 按照雌魚平均懷卵量64.5萬粒的產出率為100%計算)。危起偉等[14]也報道,1998年、2005年和2007年僅一批次繁殖活動中產卵雌魚數量就可達154尾、159尾和176尾。這說明中華鱘在葛洲壩下可以大批量正常發育成熟。對葛洲壩建壩前后中華鱘繁殖的時間對比, 也沒有發現差異。洄游距離的變化沒有影響繁殖時間和性腺成熟。Huang和Wang[17]的性腺發育模型是閉門造車, 空中樓閣, 完全不符合中華鱘的實際情況。以是否符合模型的結果來討論中華鱘的性腺退化問題, 無異于“鄭人買履”,讓實際的數據去迎合理論模型。
此外, 黃真理等[1, 17]、柯福恩等[8]對中華鱘性腺退化的認識也不符合中華鱘的生活史階段的常識。黃真理和王魯海[1]認為: 1981—1995捕撈的中華鱘中, “新股群和老股群都出現性腺退化”, 純屬憑空臆想。柯福恩等[8]認為1984年4月的樣本中有退化的性腺, 這有悖于中華鱘性腺發育的規律。這些老股群魚類還未到產卵期, 從性腺發育的卵徑、時相和成熟系數來看, 都屬于正常III—IV發育過程中的特征。他們還認為1984年10月的樣本中, 新股群也存在性腺退化的現象。如果新股群剛剛進入長江不久, 性腺就退化, 性腺退化的脅迫原因是什么? 如果沒有嚴重的脅迫因子, 性腺就發生退化, 這也有悖于魚類生活史的一般特征。
5.3 黃真理等[1]對中華鱘相關研究數據的曲意歪解
除了對中華鱘性腺發育基本認識的錯誤之外,為了證明黃真理等(Huang和Wang[17])的觀點, 黃真理和王魯海[1]對中華鱘相關研究數據進行曲意歪解。他們認為1981年中華鱘的性腺退化率為70%,其使用的性腺成熟個體數來源于曹文宣等[26]和“胡興祥(1989)”(注: 黃真理和王魯海[1]文中引用的文獻“胡興祥. 葛洲壩魚類救助工作綜述”是不存在的,第七頁是“水生所. 關于葛洲壩樞紐下游中華鱘自然繁殖的調查報告”一文[27])。當時, 曹文宣等和周春生等皆為中科院水生所第一研究室的同事, 1981年中華鱘性腺發育數據來源一致。周春生等[9]已經證實, 僅1982年調查到1尾雌魚的卵巢有不正常現象,并且卵巢上有較多的脂肪。1981年沒有中華鱘出現性腺退化的現象, 性腺退化率為0。曹文宣等[28]也沒有提及中華鱘性腺退化的現象。然而, 黃真理和王魯海[1]的文中卻有意曲解數據, 認為除了11尾性腺發育到IV期或產過卵的中華鱘之外, 其余皆為性腺退化個體, 推算結果為70%的性腺退化率。該結果是嚴重地歪曲事實, 蓄意抬高性腺退化率。實際上, 1981年1月4日葛洲壩截流, 1980年6—10月進入長江的新股群, 部分通過葛洲壩, 部分在葛洲壩下游發育, 因此1981年10—11月繁殖季節葛洲壩下游繁殖群體中老股群數量較少, 導致性腺發育成熟個體比例偏少, 并非性腺退化造成[18]。
關于1996—1998年的性腺退化率為63.5%。黃真理和王魯海[1]引用1997年的實際產卵雌魚數量為18尾(數據來源于危起偉[29])。危起偉[29]文中明確指出, 18尾僅為第一次產卵雌魚數量, 第二次產卵雌魚數量不明。黃真理和王魯海[1]選擇引用數據, 明顯低估雌魚繁殖比例, 以此來提高性腺退化率。
此外, 黃真理和王魯海[1]對陳金生等[21]的數據也進行了選擇性的使用。黃真理和王魯海[1]認為, 陳金生等[21]的“材料和方法部分語焉不詳, 比如調查樣本76尾, 來源模糊”, 而且附件記錄中只有66尾樣本的來源記錄, 還差10尾, 并得出結論陳金生等的復核報告是不可靠的。然而, 在早期的研究中, 黃真理[30]卻引用了陳金生等[21]的76尾數據為依據計算了1989年中華鱘繁殖群體數量。這說明黃真理等為了達到其不同的目的, 會特意選擇性地使用數據。
同樣, 黃真理和王魯海[1]的結果顯示, 葛洲壩截流之后中華鱘性腺退化十分嚴重, 性腺退化率為75.8%。根據該研究結果計算, 性腺成熟個體比例應小于24.2%。然而, 黃真理[30]的《利用捕撈數據估算長江中華鱘資源的新方法》和黃真理等[31]的《葛洲壩截流前后長江中華鱘繁殖群體數量變動研究》在計算中華鱘繁殖群體數量時, 引用的1984—1990年的性腺成熟個體比例均為51.1%—58%(該數據來源于趙燕等[18]和肖慧[20]), 明顯高于24.2%。這是黃真理等選擇性使用數據的證據, 同時也說明采用正確方法計算的結果與實際數據吻合, 用所謂性腺發育理論模型計算的結果則完全脫離實際。
5.4 中華鱘是否可以性腺發育成熟以及未來的保護措施
雖然葛洲壩水利樞紐修建阻隔了中華鱘的洄游通道, 造成中華鱘種群數量銳減[12, 32], 但是宜昌江段產卵場仍可以提供合適的繁殖條件[33], 滿足每次平均34或38尾雌魚完成繁殖活動[12]。這說明一般情況下, 中華鱘在葛洲壩下產卵場可以正常發育成熟, 并成功產卵繁殖。
如果中華鱘的繁殖活動和物種能夠得到有效保護, 中華鱘的種群是有可能以較小的規模長期維持的[32]。然而, 中華鱘的保護效果并不好, 其繁殖群體數量持續減少。導致這種結果的原因是多方面的: 第一, 葛洲壩下中華鱘產卵場受到三峽工程蓄水運行、航道整治等影響[16, 22, 34], 中華鱘繁殖的環境條件常常受到人類活動干擾; 第二, 捕撈對中華鱘種群長期的不利影響[32]。中華鱘同白鱀豚、白鱘、長江江豚一樣, 都是在20世紀80年代后期長江電纜捕魚興起后, 數量急劇減少, 并導致白鱀豚和白鱘功能性絕滅。目前, 長江實施了十年休漁政策, 但是在此前的多年時間里, 長江中捕撈壓力巨大, 對中華鱘繁殖群體影響明顯; 第三, 人工繁殖放流規模小對種群的補充效果不明顯[32, 35—37], 自中華鱘的人工繁殖成功以來, 相對放流量太小, 難以遏制中華鱘種群下降的趨勢。盡管如此, 目前沒有確鑿的證據表明中華鱘的性腺有大規模的退化現象。因此, 中華鱘的野生群體保護仍然具有希望。從目前存在的問題看, 未來中華鱘的保護需要采取多方面的措施, 包括野外產卵場的修復和繁殖條件的滿足, 及更大的群體放流規模, 等等。我們相信,隨著未來保護措施的落實, 中華鱘種群的恢復還是非常有可能的。
6 結語
中華鱘是長江的旗艦物種, 是長江生態系統健康的標志。保護中華鱘是我們大家的責任。保護中華鱘需要科學、客觀地分析其所面臨的困境, 因癥施策, 這樣才能保證措施的有效性。目前有關中華鱘大規模性腺退化的觀點是錯誤的, 柯福恩等[8]對性腺退化判斷的標準不科學, 將正常發育未成熟的III期性腺作為退化性腺處理, 甚至認為中華鱘剛剛進入長江不久即出現性腺退化, 不符合中華鱘性腺發育的規律。黃真理等[1, 17]沿襲了這一錯誤的認識, 并且以調查數據不符合他們的模型結果, 從而否定調查數據的正確性, 嚴重誤導公眾的認識。監測數據顯示, 葛洲壩工程沒有改變中華鱘繁殖的時間格局, 也沒有改變中華鱘性腺成熟的比例, 黃真理等的性腺分析模型是脫離實際的。而且, 黃真理等[1]還將柯福恩等稱為“吹哨人”, 一方面是將其錯誤認識以正確自居, 另一方面是想誤導大眾, 污蔑中科院水生所的正確觀點。該論文已經不再是學術研究和探討, 充滿了惡意的攻擊, 居心叵測。科學論文和期刊本應以科學探索和學術研究為主旨,可以百家爭鳴, 而不是黨同伐異。當然, 現階段中華鱘的保護確實遇到了困境。但是, 我們相信, 如果加強中華鱘野外產卵場的修復和繁殖條件的滿足, 進行更大規格、更大規模的群體放流, 中華鱘的種群還是有可能恢復的。
致謝:
本文承中國科學院水生生物研究所曹文宣院士提出修改意見, 特此致謝!
