云南松母樹和不同采種時間對球果發育及種子質量的影響
2021-01-03 10:05:14吳俊多李蓮芳張青青楊歷雨楊永潔葉桂榮吳柏良李楊濤鄭川玲
四川師范大學學報(自然科學版) 2021年1期
吳俊多, 沈 松, 李蓮芳, 張青青, 楊歷雨, 楊永潔,葉桂榮, 吳柏良, 馬 敬, 李楊濤, 鄭川玲, 楊 博, 劉 進
(西南林業大學 林學院,云南 昆明650224)
云南松(Pinus yunnanensis),松科(Pinaceae)松屬,常綠喬木,天然分布于海拔1 500 ~2 500 m 的云貴高原地區及周邊,具有耐干旱貧瘠土壤和材質優良等特點,是分布區主要的植被類型和重要的鄉土造林用材樹種之一[1-3].種子繁殖是苗木培育的主要方式之一,確定適宜的采種期可有效提高種子質量和播種品質.韓云花等[4]指出,灰楸(Catalpa fargesii)的鮮質量、出種率和千粒質量隨采種期的不同呈規律性變化,10 月底至11 月中旬是最佳采種期;喻方圓[5]指出,每年的9 月8 日至10 月31日,隨采種時間后延,金錢松(Pseudolarix amabilis)種子的出籽率逐漸提高;陳穎等[6]開展喜樹(Camptotheca acuminata)秋、冬和雪后3 個采收期的種子發芽研究,雪后采種的發芽高峰較秋季采種提前6 d,發芽率和發芽勢分別提高34. 8%和148.8%,采種期對種子品質具有重要影響.前人研究表明,林木的果實是較為穩定的遺傳性狀,不同家系的果實性狀呈現顯著的差異[7-8].雙德良等[7]根據油桐(Vernicia fordii)家系間果實形態、數量和產量等性狀的差異現象,進行其優良家系和單株良種選擇;候遠瑞等[8]利用番荔枝(Annona squamosa)果實表型性狀的差異篩選出具有不同優良性狀的家系.
已有研究指出,云南松不同時間采種顯著影響其種實的質量和品質[9-10];徐楊等[11]指出,云南松群體內不同個體的種實性狀具有豐富的遺傳變異.目前結合采種時間和母樹研究種實性狀變化的文獻較少,在已有研究的基礎上,本研究于彌渡第一代云南松無性系種子園混合家系種子培育苗木營造的11 a生人工林中,隨機選擇10株采種母樹為研究對象,從10 月底(10 月27 日)到翌年3 月中旬(3 月19日),共分6次(前3 次采種間隔時間約1 個月,后3次約為半個月)采集球果,在時間和個體差異的尺度上分析球果和種子的變化規律及其對種實的影響,豐富云南松種實成熟相關資料,為云南松以家系為主選擇最佳采種時間提供科學依據.
1 采種地及其林分概況
采種地位于昆明市宜良縣祿豐村國有林場的尖山林區.尖山林區的地理位置為24°33′70″N,103°09′54″E,海拔約1 980 m,屬亞熱帶季風氣候.據資料記載,林區年均氣溫約17.5 ℃,最熱月(8月)和最冷月(1 月)的月平均氣溫分別為22.0 ℃和8.6 ℃,年均降水量995.3 mm,年相對濕度約68%,干、濕季分明,降雨主要集中于每年的5—10月,無霜期約260 d[12-13];處于云南省中部地區,屬云南松的分布中心.
采種的云南松為采用2 a生實生苗造林的11 a生人工林,其種子來源于彌渡第一代無性系種子園自由授粉的混合家系;林地坡度10° ~15°,位于北坡,經過1 次疏伐和修枝撫育,郁閉度0.5 ~0.6,林木生長良好.
2 研究方法
在林分內隨機選擇10 株形質指標優良的I 或II級優勢木作為采種母樹,滿足結實較多、可供多次采種需求.采種前測定母樹胸徑、樹高、枝下高和冠幅.當年成熟和宿存球果的判斷依據如下:云南松的主干和側枝每年生長一個節段(即每年發一輪枝),當年成熟球果位于主稍或側枝頂端第1 輪分枝的下一輪分枝處,宿存球果則分布于其下分枝處,根據宿存球果所在枝輪的位置可區分球果成熟的年份[9,14];首次采集時,當年預成熟球果為綠色,極易辨認,據此統計當年預成熟及宿存1 a 和2 a球果數(表1).首次采種時,根據當年成熟球果的數量和采種次數(6 次:分別為2017 年10 月27 日、12 月02 日、2018 年1 月05 日、2 月10 日、2 月26日和3 月19 日)確定每次采種球果數.球果數量較多的每次采集12 個,部分母樹因球果數量不足,相應減少采集球果數至5 ~10 個.
采種時從上、中、下3 部分以及東、南、西、北4個方向均勻選取球果,如母樹球果數量不足,則優先從不同方向選擇球果.多個球果著生于同一輪枝節的則每次只采1 個,若球果成熟出現種鱗開裂且種子脫落,則不作為采集對象.采種當日將其帶回室內測定球果橫徑和縱徑(橫徑測定球果直徑最大處),每一球果分別裝入信封并編號,置于60 ℃的烘箱中[11],烘至球果種鱗開裂、種子可自由脫落時,將種子取出測定其粒數、質量以及取種時所產生的種翅和種鱗碎屑等雜質的質量(除計數外,于自然風干至恒重時測定),后將球果烘至恒重,測定其烘干質量.
以母樹(M1~M10分別為第1 ~10 株采種母樹的代號)和采種時間(T1~T6分別為第1 ~6 次采種時間的代號)為因素,構成2 因素包含60 個處理組合的完全隨機設計,其中,處理組合1 ~10 為第一批次采種的,11 ~20 為第二批次,以此類推,每個處理組合包含不等概的5 ~12 個重復(即球果數).
相關指標計算公式如下:
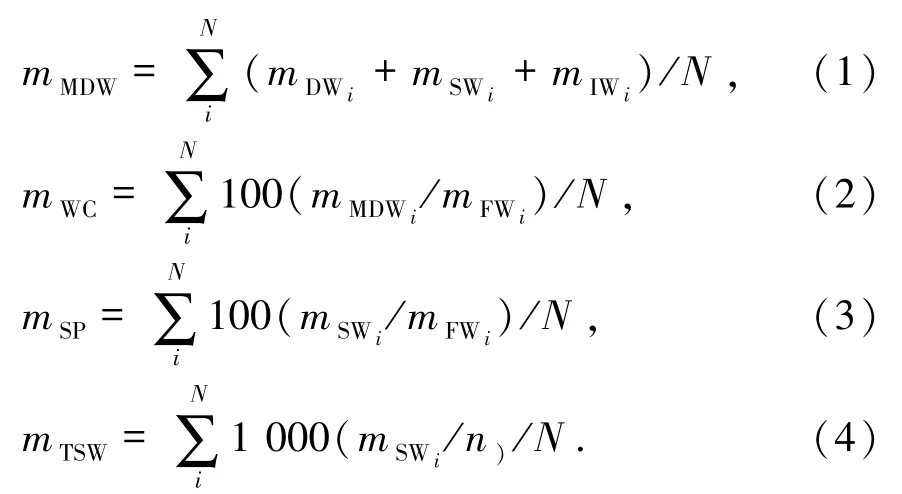
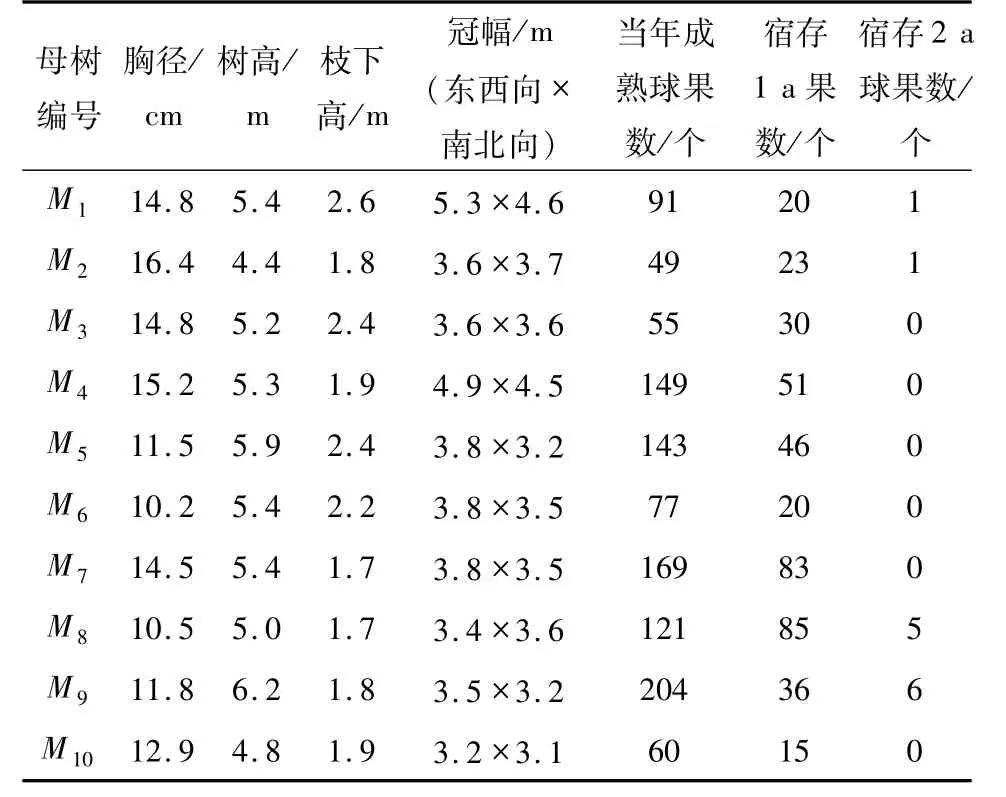
表1 采種母樹的生長指標Tab. 1 Information of maternal trees
其中,mMDW表示平均單個球果的烘干總質量(g/個),mDW為單個球果烘干質量(g/個);mSW為單個球果的種子氣干質量(g),mIW為單個球果的雜質烘干質量(g),mWC是母樹平均單個球果含水質量分數(%),mSP為母樹平均球果出種率(%),mFW為單個球果鮮質量(g/個),mTSW是每一母樹的種子千粒質量(g),n為每一球果種子數(粒),N為每一母樹的球果數(個).
采用Excel 2003 和SPSS 22.0 進行數據整理和分析,若處理組合或因素水平間差異顯著或極顯著,則以Duncan’s檢驗法進行多重比較.
3 結果與分析
3.1 球果尺寸隨母樹和采種時間的變化 處理組合球果的橫、縱徑分別為29. 6 ~46. 6 mm 和51.21 ~85.7 mm(表2).不同處理組合的此2 指標呈現極顯著的差異(P≈0.000 <0.01),其中,處理組合1、11、21、31、41 和51(M1的T1~T6)的橫徑極顯著地大于除處理組合3(M3的T1)、10(M10的T1)、30(M10的T3)、53(M3的T6)和59(M9的T6)外的其他處理組合,即橫徑較大的處理組合集中于M1、M3、M9和M10的4 株母樹;處理組合1、11、21、31、41 和51(M1的T1~T6)的縱徑均顯著或極顯著地大于其他處理組合,與橫徑相同,即M1的球果尺寸最大,且顯著大于其他母樹球果.
母樹是影響球果橫、縱徑的主導因子,其對球果尺寸具有極顯著的差異影響(P≈0.000 <0.01),單株母樹間球果尺寸的變異極為豐富(10 株母樹橫、縱徑分別呈現具有極顯著差異的5 和6 類),表明球果的尺寸大小主要由母樹決定,即受遺傳控制為可選擇的性狀,在云南松遺傳改良中,可通過幼樹選擇繁育,獲得理想尺寸要求的球果.除第一和最后一次采種外,其余時期的球果顯著縮小(表3),這是因為種鱗隨球果成熟而收縮.而最后一次采種時球果橫縱徑增大,是由球果成熟時種鱗開裂(種子未脫落)引起的.從球果橫縱徑的變化,表明每年10 月底后滇中的云南松球果趨于成熟,可準備采種.
3.2 球果的質量及水份含量變化分析處理組合的單個球果干質量和含水質量分數分別為15.4 ~50.4 g/個和8.9% ~51.8%(表4),與橫縱徑相同,不同處理組合的此2 指標也呈現極顯著的差異(P≈0.000 <0.01).其中,處理組合1、11、31、50、51 和59(M1的T1、T2、T4和T6;M9的T6;M10的T5)的球果干質量極顯著地重于其他處理組合,與球果橫縱徑類似,球果干質量最大的處理組合集中于M1、M9和M10等3 株母樹;球果含水質量分數最大的為處理組合1(M1T1),其極顯著地大于除處理組合2、3、4、6、8、9、10、11、21、23、26、27、28 和30(M2、M3、M4、M6、M8、M9和M10的T1;M1的T2;M1、M3、M6、M7、M8和M10的T3)外的其他處理組合(表5),即含水質量分數較大的主要集中于1 ~3批次(10 月26 至次年1 月5 日)所采的球果,該時期球果中水分含量較高,表明成熟初期的球果含水質量分數較高,此時期采種制種過程相對較長.球果含水質量分數也可作為表征種子成熟的指標之一,于含水量下降期開展采種,可提高制種效率.
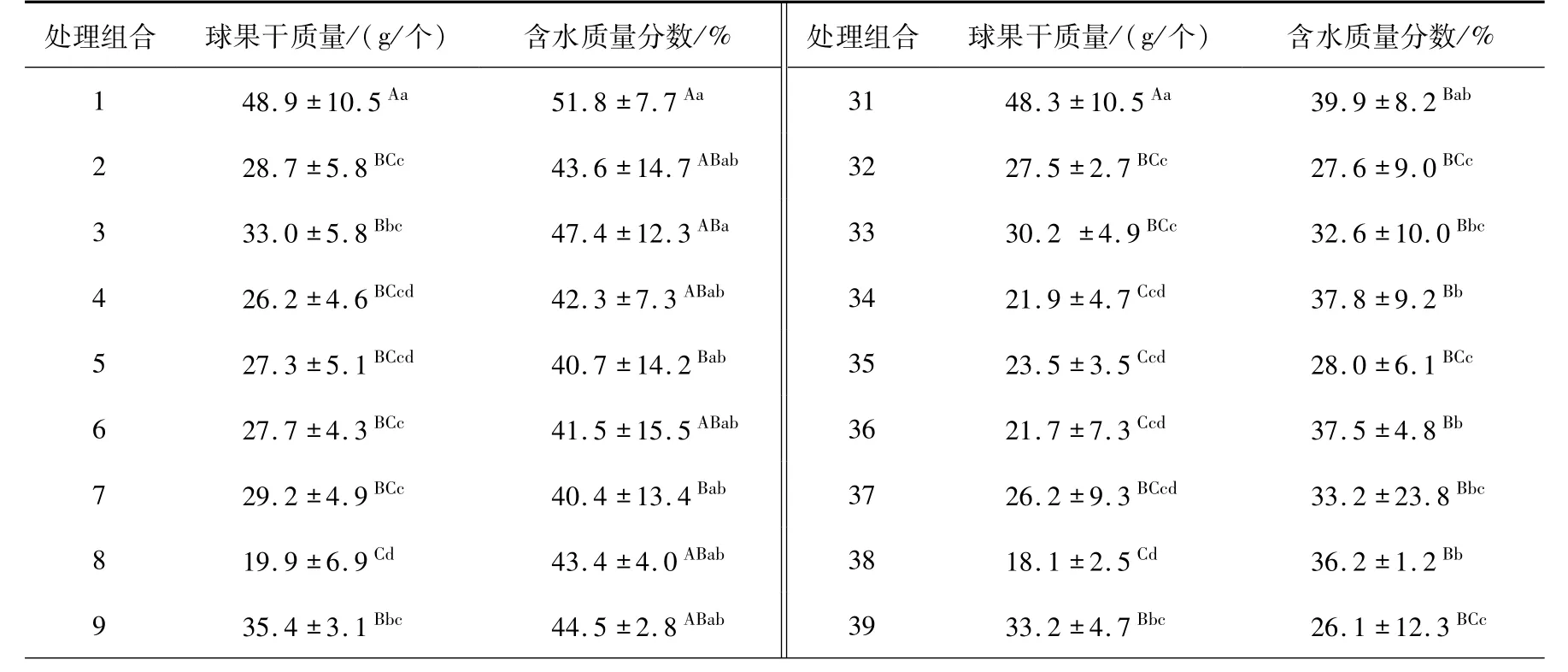
表4 處理組合的球果干質量和含水質量分數Tab. 4 Mean dry weights and moisture contents of a cone in the TCs

表 4(續)
影響球果干質量的主導因子仍為母樹,母樹間的球果干質量具有極顯著的差異(P≈0. 000 <0.01),M1的單個球果干質量(47.7g)極顯著地大于其他采種母樹(表5),即云南松球果較大的單株較少;不同采種時間的球果干質量也具有極顯著差異(P≈0.000 <0.01),第6 次采種時的球果干質量極顯著地大于除第一和第二次的其余采種時期(表5).
母樹與采種時期的交互作用對球果干質量具有極顯著的差異影響(P=0.007 <0.01),表明球果質量量受母樹和采種時間的共同作用,采種期內母樹的球果質量仍發生變化,可能是不同母樹球果的成熟期不一致引起的.
影響球果含水質量分數的主導因子為采種時期(表5),其對球果含水質量分數具有極顯著的差異影響(P≈0.000 <0.01).采種期內,含水質量分數從高到低變化隨時間呈慢(10 月27 日-翌年1月5 日)-快(1 月6 日-2 月26 日)-慢(2 月27日-3 月16 日)的趨勢,即1 月6 日至2 月26 日期間球果的含水質量分數急劇下降,該階段是球果成熟的高峰期.同樣,不同母樹的球果含水質量分數具有極顯著的差異(P≈0.000 <0.01),即不同母樹其整個成熟期球果平均含水量也不同,結合采種時間含水質量分數的差異,進一步表明母樹間球果成熟期參差不齊的現象.
在種子園中,可根據母樹成熟時間差異分單株于不同時間采種,為種子采集獲得充足的時間,合理安排時間的同時,可獲得品質優良的種子.

表5 球果干質量和含水質量分數的極差分析Tab. 5 Range analysis of the dry weights and moisture contents
3.3 不同采種時間和母樹對球果出種率和種子千粒質量的影響處理組合間球果(鮮果)的出種率和種子千粒質量分別為1.7% ~4.1%和10.4 ~20.6 g,均具有極顯著的差異(P≈0.000 <0.01).
其中,處理組合56(M6的T5)的出種率極顯著地高于處理組合36、45、46、54 和55(M6的T4和T5;M5的T4和T6;M4的T6),以上處理組合均極顯著地高于其他處理組合,即球果出種率較大的集中于第5 和6 批次采種,并集中于部分母樹,表明出種率同時受采種時間和母樹的共同作用.種子千粒質量最大的處理組合40(M10的T4,20.1 g)極顯著地高于其他處理組合,同時,處理組合10、20、30、50 和60(M10的T1~T3和T5~T6)僅低于處理組合40,表明選擇家系采種可獲得千粒質量較大的種子(表6).

表6 處理組合的出種率和千粒質量Tab. 6 Mean seed yield and 1 000-seed weights of the TCs

表 6(續)
極差分析結果表明(表7),母樹仍然是影響單個球果出種率和種子千粒質量的主導因子,其對此2 指標具有極顯著的差異影響(P≈0.000 <0.01).母樹M5 和M6 的單果出種率極顯著地大于其他母樹,M6 和M10 的出種率和種子千粒質量分別極顯著地高于其余母樹,選擇不同家系采種對提高種子收獲量具有重要作用.采種時間對以上2 指標也具有極顯著的差異影響(P≈0.000 <0.01),隨采種時間延后,球果的出種率呈上升趨勢,其主要受采種期球果含水質量分數和干質量的影響.

表7 出種率和千粒質量的極差分析Tab. 7 Range analysis of the seed yield and 1 000 -seed weights of cone
3.4 球果和種子指標間的相關分析Person相關分析結果(表8),球果橫徑和縱徑與干質量、含水質量分數和種子千粒質量間均呈極顯著正相關(R=0.161 ~0.873,P≈0.000 <0.01),表明球果尺寸大小可指示其質量、含水質量分數和種子質量狀況,采種時選擇尺寸較大的球果可獲得相對較優的種子,與前述結果相一致.球果出種率則與球果橫徑、干質量和含水質量分數呈極顯著的負相關(R=-0.558 ~-0.189,P≈0.000 <0.01),表明球果尺寸和含水質量分數越大,其出種率則越低,符合球果出種率變化規律.

表8 相關分析Tab. 8 The correlation analysis
4 結論和討論
4.1 結論以滇中地區11 a 生云南松人工林為對象,分6 個時期對10 株母樹進行采種,采用2 因子完全隨機設計分析球果和種子性狀的變化.處理組合的球果橫徑、縱徑、干質量、含水質量分數、出種率和種子千粒質量分別為29. 6 ~46. 6 mm、51.21 ~85. 7 mm、15. 4 ~50. 4 g/個、8. 9% ~51.8%、1.7% ~4.1%和10.4 ~20.6 g,不同處理組合的以上指標均具有極顯著的差異(P≈0.000 <0.01).其中,影響球果橫徑、縱徑、干質量和種子千粒質量的主要因子為母樹,影響球果含水質量分數的則為采種時間;相關性分析結果,橫徑和縱徑與球果干質量、含水質量分數及種子千粒質量間均呈極顯著正相關(R=0. 161 ~0. 873,P≈0. 000 <0.01),即球果尺寸可指示種實的質量.
4.2 討論云南松人工林主要采用種子繁殖,適宜的時間采種可獲得優質種子,有益于壯苗培育[15].據資料記載,云南松球果的成熟期為每年的12 月至次年3 月,成熟球果的種鱗于2—3 月開裂釋放種子,部分球果則宿存于枝條上逐漸開裂,宿存時間可長達9 a[9,14].李濱等[10]于12 月至3 月對西昌地區的云南松采種研究指出,球果干質量、含水質量分數和千粒質量隨采種時間的推遲逐漸下降,出種率則不斷提高,其中球果含水質量分數于12月份穩定在40%左右,1 月份后迅速下降.本研究采種的初始時間提前48 d,結束時間則相對一致;含水質量分數、出種率和種子千粒質量隨采種時間的變化呈相同的變化規律.本研究與已有研究具有共性,也有差異,不同之處受云南松采種地不同決定.球果尺寸隨采種期延后,呈現縮小的趨勢,是由于采種時球果已發育成熟,隨時間變化,球果尺寸隨體內的含水量下降而種鱗縮小.
針葉樹種的球果和種子性狀受環境和遺傳因子的雙重調控,已有研究指出,遺傳因素是控制球果和種子性狀變異的主要因子[11,15-17].徐亮等[16]指出,岷山柏(Cupressus chengiana)種群間球果和種子的大小具有顯著的遺傳變異;王慶華等[17]研究禿杉(Taiwania flousiana)8 個種源52 個家系球果和種子的形態變異指出,群體內的分化差異大于群體間,表明同一群體內存在豐富的遺傳多樣性.在本研究中,同一林分中不同母樹的橫徑、縱徑、干質量、含水質量分數、出種率和種子千粒質量均具有極顯著的差異,與已有研究結果相似,即單株間球果和種子現狀呈現極顯著變異,進一步為云南松遺傳改良的單株選擇奠定理論基礎.
致謝西南林業大學徐婷婷、凌麗芳、李阿花、顏杰和夏云杉等同學參與了指標的測定和數據的收集整理,在此謹致謝意.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國生殖健康(2019年2期)2019-08-23 08:12:08
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
產品可靠性報告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
