蟲害誘導植物信息化合物介導的植物間交流及機制
2021-01-04 01:03:39姚誠誠杜立嘯李云河
植物保護 2021年6期
姚誠誠 杜立嘯 李云河
摘要 :當遭受植食性昆蟲取食時,植物會釋放復雜和多樣的揮發(fā)性有機化合物。蟲害誘導的揮發(fā)物在調(diào)控植物與不同營養(yǎng)層昆蟲群落相互關(guān)系中發(fā)揮重要作用。同時,鄰近植物也能感知這種蟲害誘導揮發(fā)物,對可能即將到來的蟲害威脅做出防御準備。獲得防御準備的植物在受到蟲害損傷時,會立即啟動相應的防御反應。為了應對植物的這種害蟲防御策略,某些昆蟲可通過調(diào)控植物揮發(fā)物信號來傳遞“假信息”,從而抑制相鄰植物的防御反應。這種信息交流既可發(fā)生在同種植物之間,也可發(fā)生在不同科屬的植物之間。近年,植物間的信息交流現(xiàn)象已得到廣泛研究,但相關(guān)的生化和分子機制及生態(tài)學意義尚不清楚。本文主要探討了蟲害誘導信息化合物介導的植物間交流類型、機制和生態(tài)學意義,以及影響植物間交流的生物或非生物因子,分析了利用植物間信息交流開展害蟲綠色防控的前景。
關(guān)鍵詞 :蟲害誘導的植物揮發(fā)物; 植物與植物交流; 植物與昆蟲互作
中圖分類號:
Q948.1, S433
文獻標識碼: A
DOI: 10.16688/j.zwbh.2020427
Plant-to-plant communications medicated by herbivore-induced plant volatiles and the mechanisms
YAO Chengcheng, DU Lixiao*, LI Yunhe*
(State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection,
Chinese Academy of Agricultural Sciences, Beijing 100193, China)
Abstract
Plants, when being attacked by herbivores, will constitutively emit a wide variety of volatile organic compounds, called herbivore-induced plant volatiles (HIPVs), which plays a key role in mediating the interactions between plants and diverse trophic levels of insects.In addition, neighboring plants can perceive these distress signals from herbivore-attacked plants and prepare well for incoming threats.Plants that have primed their defenses can immediately activate corresponding defensive responses once they were damaged by herbivorous insects.Conversely, some insects may manipulate their host plants to emit volatiles that disseminate the deceptive information to neighboring plants, which will suppress their defenses.The phenomenon of HIPVs-medicated plant-plant communication occurs among the plants of the same or different families and genera.Although the concept of this communication has been widely accepted, its biochemical and molecular mechanisms and ecological significances remain largely obscure.In this overview, we reviewed the types, mechanisms and significances of plant-plant communications mediated by HIPVs, and we focused on both ecological factors affecting plant-plant communications and the prospects for using volatiles to control pests based on plant-plant communications.
Key words
herbivore-induced plant volatiles; plant-plant communication; plant-insect interaction
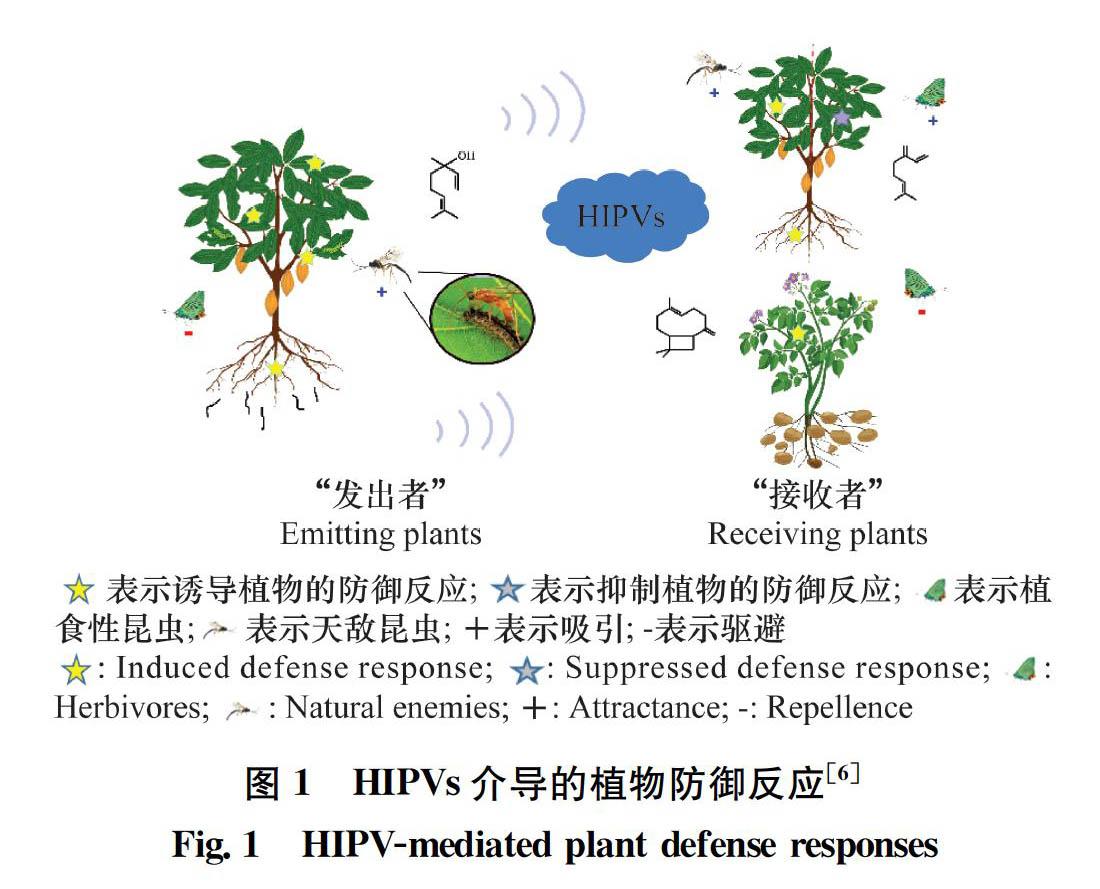
當遭受植食性昆蟲取食時,植物會產(chǎn)生一系列防御反應。一方面,植物會產(chǎn)生有毒化合物、抗營養(yǎng)酶類和抗消化酶類等物質(zhì)直接毒殺或抑制植食性昆蟲生長發(fā)育;另一方面,植物會釋放揮發(fā)性化合物吸引捕食性或寄生性天敵來防御害蟲[12]。此外,相鄰植株也能感知這種蟲害誘導植物揮發(fā)物(herbivore-induced plant volatiles, HIPVs),相鄰植株接收HIPVs后做出的反應主要有兩類:一類是直接激活自身的防御反應[35];另一類是不啟動防御反應,而是“武裝”自己,在受到植食性昆蟲為害時,可啟動更強或更快的防御反應[2, 58]。
蟲害誘導植物揮發(fā)物介導的植物間交流現(xiàn)象最早發(fā)現(xiàn)于柳樹中[9]。天幕毛蟲Malacosoma californicum為害的柳樹Salix sitchensis誘使其鄰近的健康柳樹植株對天幕毛蟲的抗性增強。這一報道引起了研究者對植物間交流現(xiàn)象的廣泛關(guān)注,相關(guān)報道日益增多[1014]。此外,也有研究發(fā)現(xiàn),某些植食性昆蟲會利用蟲害誘導植物揮發(fā)物抑制相鄰植物的防御反應,從而為種內(nèi)其他昆蟲創(chuàng)造更加有利的生存環(huán)境[1516]。本文通過探討蟲害誘導植物揮發(fā)物介導的植物間交流類型、機制和生態(tài)學意義,以及影響植物間交流的生物和非生物因子,分析利用植物間信息交流現(xiàn)象開展害蟲綠色防控的前景,以期為該領(lǐng)域的研究者提供一定的啟發(fā)和參考。
1 HIPVs介導的植物間交流
在自然界中,植物間交流的現(xiàn)象普遍存在[7, 1721]。當遭到植食性昆蟲為害時,植物會釋放HIPVs吸引害蟲天敵或驅(qū)避成蟲產(chǎn)卵,減輕危害;同時,鄰近同種或異種植物能夠感知這種HIPVs,對可能即將到來的蟲害威脅進行預判,做出防御準備,提高對害蟲的防御能力。然而,某些昆蟲會利用植物的這種特性,調(diào)控植物向鄰近植物傳遞“假信息”,抑制鄰近植物的防御反應(圖1)。下文對HIPVs調(diào)控鄰近植物防御反應的兩種類型分別進行闡述。
1.1 HIPVs誘導植物的防御反應
1.1.1 HIPVs誘導同種植物間的防御反應
目前,關(guān)于HIPVs誘導植物間防御反應的研究主要集中在同種植物之間[19]。研究發(fā)現(xiàn),二斑葉螨Tetranychus urticae為害的棉花Gossypium hirsutum釋放的植物揮發(fā)物會降低鄰近棉花上二斑葉螨的落卵量[22];大菜粉蝶Pieris brassicae為害的甘藍Brassica oleracea var.capitata釋放的植物揮發(fā)物增強了鄰近甘藍對大菜粉蝶的抗性,同時還吸引菜粉蝶寄生蜂粉蝶盤絨繭蜂Cotesia glomerata[10];黏蟲Mythimna separata為害的玉米Zea mays L.cv.Royal Dent釋放的植物揮發(fā)物會導致鄰近玉米上的黏蟲生長發(fā)育受阻[11]。由于相同的遺傳背景,同種植物在應對蟲害時的防御反應一致,識別危險信號分子上也存在保守性,因此,在同種植物間利用HIPVs進行交流的效率更高[45, 23]。被植食性昆蟲為害的植物釋放HIPVs來調(diào)整鄰近植物的狀態(tài),一方面干擾害蟲的行為選擇,來減輕對自身的為害;另一方面利用植物群體防御來提高物種自身的穩(wěn)定性[45]。但是同種物種之間也存在著激烈的生存競爭,植物受到蟲害后,釋放HIPVs有自身防御的需要,但這種信號被鄰近植物收到,是有意為之,還是鄰近植物“竊聽”的結(jié)果,目前尚無定論。
1.1.2 HIPVs誘導異種植物間的防御反應
與同種植物間的交流相比,HIPVs誘導異種植物防御反應的研究很少。研究顯示,海灰翅夜蛾Spodoptera littoralis為害的棉花釋放的植物揮發(fā)物可提高鄰近紫苜蓿Medicago sativa的防御水平,導致海灰翅夜蛾在苜蓿上的產(chǎn)卵量下降[24]。這種跨越物種的交流現(xiàn)象,究竟是不同物種抵御相同害蟲的需要還是自然進化過程中群體選擇的需要,目前還存在爭議[5]。
1.2 HIPVs抑制植物的防御反應
雖然多數(shù)研究表明,HIPVs可誘導鄰近植物對植食性昆蟲產(chǎn)生抗性,但最近的研究發(fā)現(xiàn),昆蟲也可通過調(diào)控植物揮發(fā)物,誘使鄰近植物更適合昆蟲的發(fā)育。譬如,煙粉虱Bemisia tabaci為害的番茄Lycopersicon esculentum cv.Moneymaker釋放的植物揮發(fā)物不但沒有誘導鄰近番茄對煙粉虱的抗性,反而使其更適合煙粉虱取食[16]。小菜蛾P(guān)lutella xylostella為害的甘藍釋放的植物揮發(fā)物同樣未能增強鄰近甘藍的抗蟲性,反而增加了小菜蛾在鄰近甘藍上的落卵量[15]。
我們推測在長期的進化過程中,昆蟲適應了寄主植物的防御反應,利用植物間交流的“漏洞”,通過調(diào)控受害植物的HIPVs向鄰近植物傳遞“假”信號抑制鄰近植物的防御反應,為其后代提供更有利的生存環(huán)境。有關(guān)HIPVs抑制鄰近植物防御反應的現(xiàn)象還知之甚少,而HIPVs是否會抑制鄰近異種植物防御反應目前也未見報道。
2 HIPVs調(diào)控植物防御反應的機制
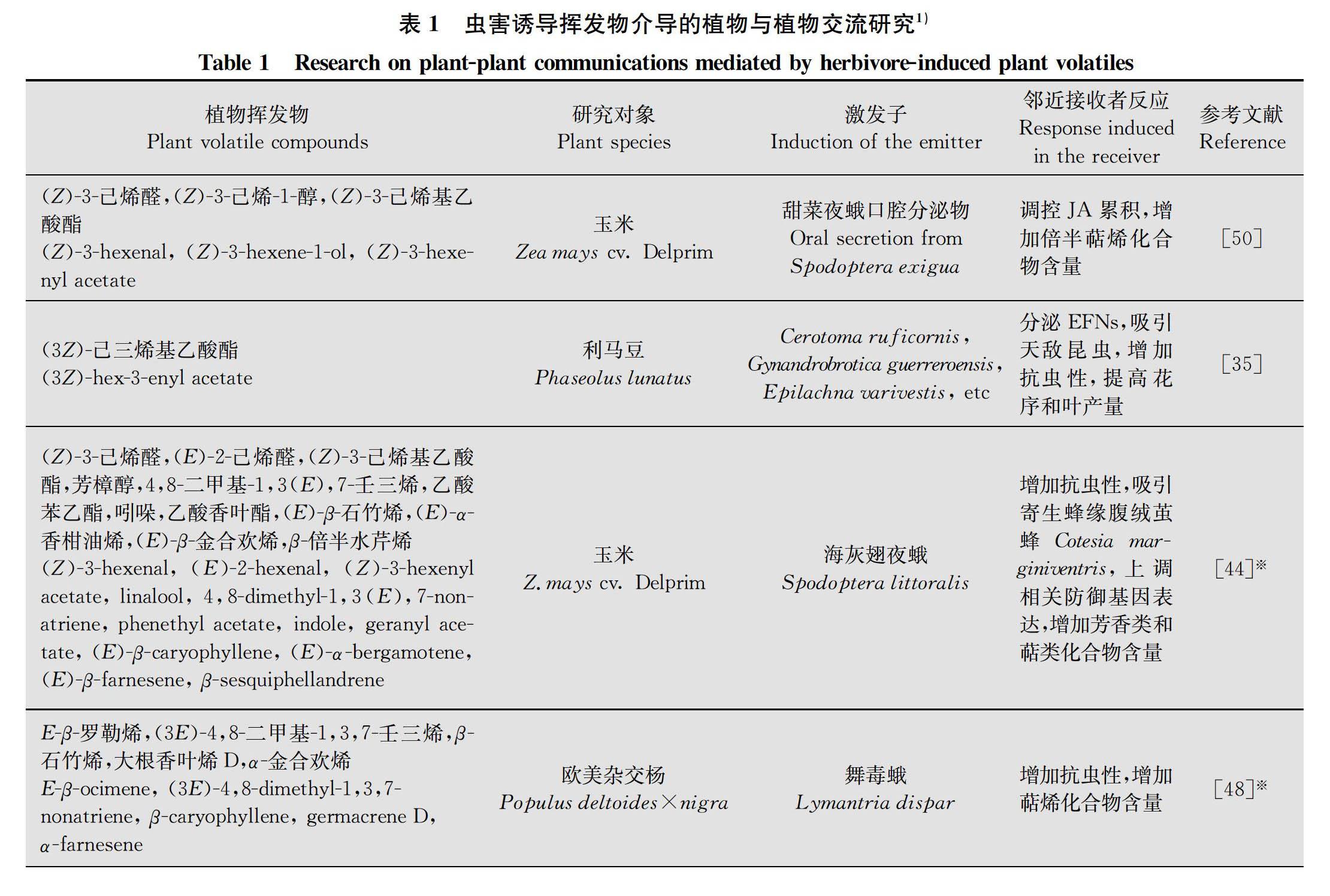
研究者通過對“發(fā)出者(emitting plants)”釋放的揮發(fā)物進行實時檢測,以及對“接收者(receiving plants)”接收到化學信號后所做出的反應進行了研究,發(fā)現(xiàn)蟲害誘導植物揮發(fā)物在植物間交流中發(fā)揮著媒介作用(表1),因此,這種蟲害誘導揮發(fā)物常常被稱為“植物語言(plant language)”[2530]。
2.1 HIPVs誘導鄰近植物或同一植物不同組織防御反應的機制
HIPVs誘導鄰近植物的防御反應包括直接防御和間接防御。直接防御是鄰近植物接收到HIPVs信號后,通過誘導防御基因表達,調(diào)控植物激素和產(chǎn)生防御相關(guān)次生代謝物質(zhì)等一系列過程,使植物對植食性昆蟲產(chǎn)生直接的生理抗性[34, 3133];間接防御是被HIPVs誘導的鄰近植物通過分泌花外蜜露(extrafloral nectars, EFNs)或釋放揮發(fā)性化合物吸引捕食性或寄生性天敵昆蟲,間接地降低植食性昆蟲的危害[3438]。
2.1.1 HIPVs誘導鄰近植物的直接防御機制
甘薯Ipomoea batatas遭到海灰翅夜蛾取食后迅速釋放出(E)-4,8-dimethyl-1,3,7-nonatriene (DMNT),DMNT誘導鄰近甘薯中蛋白酶抑制劑(sporamin protease inhibitor, SPI)基因的表達上調(diào),使葉片中胰蛋白酶抑制劑的含量提高,進而增強了鄰近甘薯對海灰翅夜蛾的抗性[33]。歐洲榿木Alnus glutinosa受到藍毛臀螢葉甲Agelastica alni為害后釋放的DMNT、β-ocimene等會誘導鄰近榿木蛋白酶抑制劑和酚類物質(zhì)的活性提高[39];β-羅勒烯(β-ocimene)能夠誘導鄰近健康大白菜Brassica rapa var.glabra水楊酸和茉莉酸相關(guān)防御基因上調(diào)表達,提高芥子油苷的含量,從而抑制桃蚜Myzus persicae的生長發(fā)育,改變其取食行為[40]。這說明HIPVs作為植物間交流的信號分子可以激活植物防御反應,提高植物抗蟲性。此外,鄰近植物還可以直接利用HIPVs合成具有防御功能的化合物,抑制植食性昆蟲生長發(fā)育。例如,番茄被斜紋夜蛾Spodoptera litura為害后釋放出大量的(Z)-3-己烯醇[(Z)-3-hexenol],鄰近番茄吸收(Z)-3-hexenol后會將其轉(zhuǎn)化成(Z)-3-hexenylvicianoside,導致斜紋夜蛾生長發(fā)育受阻,成活率下降[41]。
研究表明,HIPVs誘導鄰近植物防御反應主要受植物激素的調(diào)控,其中茉莉酸(jasmonic acid, JA)、茉莉酸異亮氨酸(jasmonoyl-isoleucine, JA-Ile)、水楊酸(salicylic acid, SA)和脫落酸(abscisic acid, ABA)等被證實在植物防御過程中發(fā)揮著重要調(diào)控作用[42]。例如,吲哚(indole)能夠誘導鄰近水稻Oryza sativa中JA途徑相關(guān)防御基因的表達,以及12-oxophytodienoic acid (OPDA)和JA的累積量增加,增強水稻對草地貪夜蛾Spodoptera frugiperda幼蟲的抗性[32];同時,吲哚還能夠誘導鄰近玉米中JA-Ile和ABA的累積量上升,進而增加綠葉揮發(fā)物和萜烯類化合物的釋放量,增強玉米對海灰翅夜蛾幼蟲的防御[12]。此外,無論是專食性害蟲煙草天蛾Manduca sexta還是多食性害蟲煙芽夜蛾Heliothis virescens為害本生煙Nicotiana benthamiana后植物釋放的HIPVs均能夠誘導鄰近植物的防御準備;對鄰近植物采用機械的方式損傷并涂抹任意一種害蟲的口腔分泌物均會顯著增加本生煙中JA含量,調(diào)控本生煙防御性揮發(fā)物的代謝過程,增加HIPVs的釋放[43]。但也有研究表明,HIPVs誘導鄰近植物的抗性反應獨立于植物激素代謝通路。比如,海灰翅夜蛾幼蟲為害的甘薯會誘導鄰近甘薯葉片中胰蛋白酶抑制劑含量升高,增強對海灰翅夜蛾幼蟲的抗性,但并未引起鄰近葉片中JA的累積[33]。同時Sugimoto等[41]證實番茄吸收鄰近蟲害植株釋放的(Z)-3-hexenol,將其轉(zhuǎn)化成對斜紋夜蛾生長發(fā)育不利的(Z)-3-hexenylvicianoside,但植株中JA的含量并沒有顯著升高。
2.1.2 HIPVs誘導鄰近植物的間接防御機制
HIPVs除了誘導鄰近植物產(chǎn)生直接防御反應,也會誘導鄰近植物釋放揮發(fā)性有機化合物(volatile organic compounds,VOCs)吸引天敵而間接地防御植食性昆蟲。番茄被海灰翅夜蛾為害后釋放的植物揮發(fā)物誘導鄰近番茄釋放β-ocimene等物質(zhì)吸引阿爾蚜繭蜂Aphidius ervi,降低海灰翅夜蛾的為害[14]。玉米受到斑禾草螟Chilo partellus (Swinhoe)為害后釋放的植物揮發(fā)物會誘導鄰近玉米釋放DMNT吸引卵寄生蜂布氏赤眼蜂Trichogramma bournieri和幼蟲寄生蜂大螟盤絨繭蜂Cotesia sesamiae [13]。海灰翅夜蛾為害的玉米釋放的植物揮發(fā)物能夠誘導鄰近玉米釋放芳香類和萜類化合物提高對緣腹絨繭蜂Cotesia marginiventris的引誘作用[44]。
此外,研究還發(fā)現(xiàn),HIPVs會促進鄰近植物分泌花外蜜露吸引天敵。例如,蟲害誘導利馬豆Phaseolus lunatus釋放的(3Z)-hex-3-enyl acetate能夠誘導鄰近利馬豆分泌花外蜜露吸引捕食性和寄生性天敵,降低甲蟲和象甲等植食性昆蟲的為害[35]。
2.1.3 HIPVs誘導同一植物鄰近健康組織的防御機制
HIPVs能夠誘導植物損傷部位的防御,同時還可以警告鄰近組織做好應對蟲害的準備。當遭到植食性昆蟲取食時,已經(jīng)接收到HIPVs的健康組織會立即啟動防御反應降低害蟲為害[3, 4547]。例如,被蟲侵害的歐美雜交楊Populus deltoides × nigra葉片釋放的萜烯化合物E-β-ocimene、DMNT、β-caryophyllene、germacrene D、α-farnesene均能夠顯著提高鄰近葉片中萜烯化合物含量,增強楊樹對舞毒蛾Lymantria dispar幼蟲的抗性[48]。被蟲侵害的玉米葉片釋放的吲哚可以顯著提高鄰近葉片中萜烯類化合物含量[12]。對于植物自身而言,當某些部位不可避免要受到植食性昆蟲為害時,它們就可能會釋放HIPVs信號,克服長距離和維管束的限制,快速到達鄰近組織警告威脅的存在,以便快速啟動全身的防御系統(tǒng);對于受害組織來說,釋放HIPVs也是誘導受害組織啟動防御反應抵御蟲害的需要[45]。
2.2 HIPVs抑制鄰近植物防御反應的機制
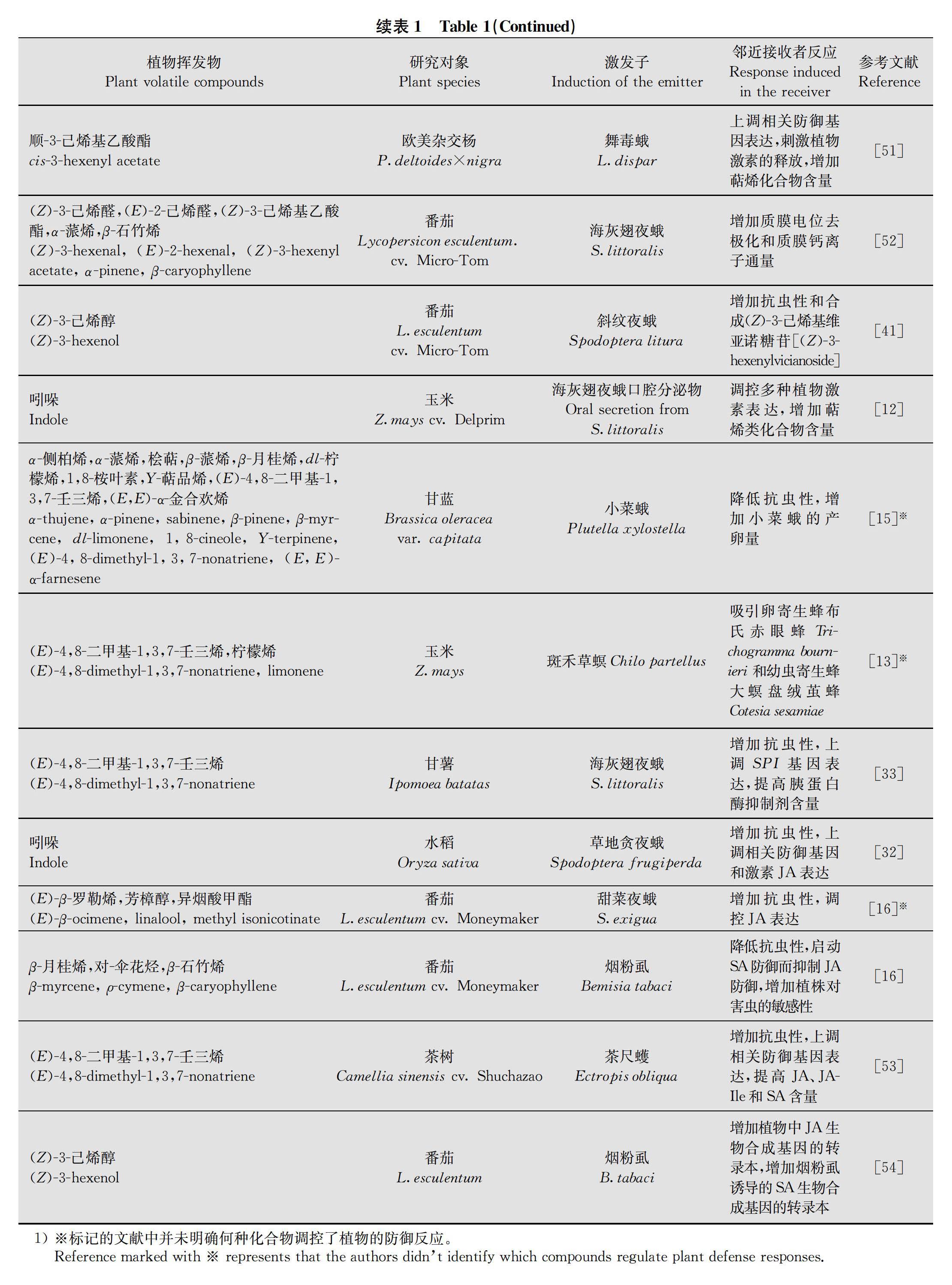
除了誘導鄰近植物對植食性昆蟲的防御,某些HIPVs還會充當植物防御抑制劑,抑制植物對害蟲的防御反應[42]。比如,煙粉虱為害番茄釋放的β-myrcene、ρ-cymene、β-caryophyllene能夠顯著提高鄰近番茄中SA含量,誘導抗性基因PR-1a和PR-1b的上調(diào)表達,而抑制JA通路相關(guān)的抗性基因PⅠ-Ⅰ和PⅠ-Ⅱ的表達,進而使鄰近番茄更容易受到煙粉虱的侵害[16]。田間試驗也發(fā)現(xiàn),田間釋放綠葉揮發(fā)物(green leaf volatiles; GLVs)、(Z)-3-hexenal、(Z)-3-hexenol、(E)-2-hexenal和(Z)-3-hexenyl acetate,盡管可以提高玉米植株萜烯類化合物的釋放,但并未降低植食性昆蟲對玉米的為害,反而造成玉米受蟲害加重[49]。
3 影響HIPVs介導植物間交流的因子
HIPVs介導的植物間交流受到各種生物或非生物因素的影響,例如植物間的距離、空氣流通程度、物種間親緣關(guān)系、物種特異性等都會影響它們之間交流的效率。
3.1 距離
植物間交流的效率會隨著植物間距離的增加而降低[51]。在同一濃度同一劑量下,野生煙草Nicotiana attenuata距離修剪的三齒蒿Artemisia tridentata 10 cm或15 cm之內(nèi)時,受到害蟲為害的程度會明顯減低,然而隨著距離增加,這種效應會逐漸降低[5556]。同種三齒蒿植株之間也能進行交流[57],這種交流能達到60 cm[58],利馬豆間能達到50 cm[47],然而超過該閾值,植物間交流的效率會逐漸降低直至消失。研究表明植物接收HIPVs信號分子與細胞膜去極化和細胞質(zhì)Ca2+濃度變化有關(guān)。而通過不同濃度的α-pinene 和β-caryophyllene處理番茄發(fā)現(xiàn),高濃度化合物處理下番茄細胞膜去極化程度更高,這說明植物間交流存在濃度效應[52]。因此,隨著植物間距離的增加,HIPVs由蟲害植物傳遞到鄰近植物的量必然會大大降低,它們交流的效率也會下降甚至消失。
3.2 空氣流通
對于植物的地上部分來說,空氣流通是植物間信號交流的必要因素[55, 5860]。機械損傷玉米葉片并涂抹海灰翅夜蛾口腔分泌物一段時間后,損傷鄰近葉片并收集揮發(fā)物發(fā)現(xiàn),萜烯類化合物含量顯著增加;而當用聚四氟乙烯袋封閉蟲害葉片,阻斷蟲害葉片和鄰近葉片的空氣流通時,同樣時間段內(nèi)在相同處理的鄰近葉片中未發(fā)現(xiàn)萜烯類化合物含量的改變[12]。
3.3 親緣關(guān)系
親緣關(guān)系近的植物在信息化合物交流中可能會有額外的優(yōu)勢,這一優(yōu)勢被稱為親屬選擇假說(Kin-Selection Hypothesis)[61]。Karban等[23]認為來自親屬的揮發(fā)物信號可能更容易被同類所察覺。通過對兩種不同品系玉米的研究發(fā)現(xiàn),健康玉米暴露在同一品系玉米釋放的經(jīng)蟲害誘導的揮發(fā)物中,對寄生蜂的吸引性更強[13]。這一現(xiàn)象在發(fā)草Deschampsia cespitosa[62]、擬南芥Arabidopsis thaliana[63]和三齒蒿[23, 59, 64]等植物中都有報道。
研究發(fā)現(xiàn),某些植物只能和部分鄰近植物進行交流,而對其他植物無反應,植物間的交流存在著物種特異性。譬如,海灰翅夜蛾為害的棉花植株釋放出的揮發(fā)物增加了鄰近棉花和苜蓿植物的抗蟲性,降低了海灰翅夜蛾的產(chǎn)卵量;然而海灰翅夜蛾為害的苜蓿和埃及車軸草Trifolium alexandrinum植株釋放的揮發(fā)物并沒有增加鄰近棉花的抗蟲性[24]。玉米遭到海灰翅夜蛾為害后可以迅速地釋放吲哚,顯著增強鄰近玉米的抗蟲性,輕微地誘導鄰近棉花的抗蟲性,而沒有誘導鄰近豇豆Vigna unguiculata的抗蟲性[12]。由此可見,植物能夠特異性識別鄰近植物釋放的揮發(fā)物。因此,根據(jù)植物間交流的物種特異性,我們可以利用特定植物的揮發(fā)物防御害蟲,實現(xiàn)綠色防控。
3.4 植食性昆蟲
植食性昆蟲也是驅(qū)動HIPVs調(diào)控植物間信息傳遞的重要因子。北美一枝黃花Solidago altissima被黃花葉甲蟲Trirhabda virgate為害后釋放的HIPVs能夠激發(fā)鄰近相同甚至不同基因型的植物提高對甲蟲的防御水平;而采用未經(jīng)甲蟲誘導的揮發(fā)物處理植物,只有鄰近相同基因型的植物會啟動同等防御,不同基因型的植物沒有表現(xiàn)同樣的反應[17]。這一結(jié)果說明植食性昆蟲會驅(qū)動植物進化出更開放的交流機制。
此外,植物個體的發(fā)育程度、植物暴露揮發(fā)物的時間和濃度、植食性昆蟲為害的時間和強度以及氣候變化等生物或非生物因素都影響著植物與植物交流的效率[15, 32, 6566]。
4 展望
在HIPVs介導的植物間交流中,HIPVs的釋放和傳遞通常對“接收者”是有益的,但是對于“發(fā)出者”產(chǎn)生了一種耗能代價[6768]。提高鄰近植物的抗性可能改變了“發(fā)出者”和“接收者”的競爭性平衡,這種平衡對“發(fā)出者”表現(xiàn)不利,但幫助“接收者”成功地降低了害蟲的攻擊[69]。植物之間互相競爭營養(yǎng)等資源,而植物間的交流為“接收者”提供了可靠的信息,但對“發(fā)出者”沒有表現(xiàn)出明顯的優(yōu)勢,為什么植物會浪費能量幫助自己的競爭對手去應對即將來臨的威脅[70]?目前普遍認為,HIPVs的“發(fā)出者”可能并非是對“接收者”有意圖的警告,更可能是“接收者”的竊聽行為[19, 69, 7172]。因此,在未來的研究中,我們既要評估“發(fā)出者”和“接收者”的適應性,也要比較二者對資源的需求度,充分理解二者之間的關(guān)系,才能更好地利用HIPVs介導植物間的交流。
目前對植物間交流的探究多集中在實驗室研究階段,且多數(shù)研究使用人工合成的揮發(fā)物組分模擬HIPVs的釋放,導致?lián)]發(fā)物處理濃度遠遠高于HIPVs的真實釋放量,由此得出的結(jié)果是否能夠代表自然界中存在的真實情況還有待進一步的驗證[45]。這也是目前限制HIPVs調(diào)控植物間信息傳遞研究繼續(xù)深入的關(guān)鍵。因此,研究者應該更加關(guān)注自然界中植物間的交流,特別是自然界中HIPVs的濃度,這對未來利用HIPVs進行害蟲的綠色防控至關(guān)重要。此外,鄰近植物識別HIPVs的過程對于理解植物間交流至關(guān)重要,在植物體內(nèi)是否存在類似昆蟲嗅覺識別系統(tǒng)的神經(jīng)調(diào)控網(wǎng)絡,還不得而知[4];在接收到HIPVs后,植物如何利用這些化學信號來調(diào)控自身的防御系統(tǒng),究竟是直接利用化合物進行調(diào)控還是這些化合物僅作為調(diào)控信號仍缺乏深入研究[45]。
近年來,對植物間交流的研究逐漸增多,利用植物間信息交流開展害蟲綠色防控具有非常廣闊的前景。比如,基于相鄰植物間交流的理論,種植哨兵植物預先察覺即將到來的威脅,警告鄰近植物做好防御準備[2, 73];或利用植物間信息交流中的生物活性揮發(fā)物組分,通過基因修飾(轉(zhuǎn)基因植物)增加植物對昆蟲的敏感性,快速釋放關(guān)鍵活性揮發(fā)物警告鄰近植物[74]。目前農(nóng)業(yè)生產(chǎn)上采用的相鄰植物間“推拉”調(diào)控策略效果顯著,通過選擇一種或多種驅(qū)避(推)害蟲的植物以及吸引(拉)它們?nèi)∈车闹参铮贡槐Wo作物免受侵擾[2, 73, 75]。
除HIPVs介導植物間的交流外,病原菌誘導的植物揮發(fā)物(pathogen-induced plant volatiles; PIPVs)[7679]及機械損傷誘導的植物揮發(fā)物(wound-induced plant volatiles; WIPVs)[5658, 8083]也會介導植物間的交流,誘導鄰近植物產(chǎn)生抗病性或抗蟲性。因此,我們還可以采取一些機械方法在害蟲為害之前預防并降低其為害,比如,對植株采取機械性修剪[59]、去頂[84]以及人為的干擾觸動[85],一方面降低害蟲對植物的為害,另一方面也誘導了其鄰近植物的抗蟲性。
此外,未受脅迫的植物釋放的組成型揮發(fā)物也能夠介導植物間的交流。健康洋蔥Allium cepa釋放的揮發(fā)物會改變鄰近馬鈴薯Solanum tuberosum揮發(fā)物的釋放模式,驅(qū)避桃蚜Myzus persicae[86]。我們推測,植物之間會相互競爭資源,“接收者”通過獲取未受脅迫的“發(fā)出者”揮發(fā)物中的重要組分,從而提高自身的適應性。
參考文獻
[1] POELMAN E H, DICKE M. Plant-mediated interactions among insects within a community ecological perspective [M]∥Annual plant reviews 47: insect-plant interactions. New York: Wiley, 2014: 309338.
[2] TURLINGS T C J, ERB M. Tritrophic interactions mediated by herbivore-induced plant volatiles: mechanisms, ecological relevance, and application potential [J]. Annual Review of Entomology, 2018, 63(1): 433452.
[3] HEIL M, BUENO J C S. Within-plant signaling by volatiles leads to induction and priming of an indirect plant defense in nature [J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(13): 54675472.
[4] HEIL M. Herbivore-induced plant volatiles: targets, perception and unanswered questions [J]. New Phytologist, 2014, 204(2): 297306.
[5] KARBAN R, YANG L H, ESWARDS K F. Volatile communication between plants that affects herbivory: a meta-analysis [J]. Ecology Letters, 2014, 17(1): 4452.
[6] DAS A, LEE S H, HYUN T K, et al. Plant volatiles as method of communication [J]. Plant Biotechnology Reports, 2013, 7(1): 926.
[7] DELORY B M, DELAPLACE P, FAUCONNIER M L, et al. Root-emitted volatile organic compounds: can they mediate belowground plant-plant interactions? [J]. Plant Soil, 2016, 402(1/2): 126.
[8] 王杰, 宋圓圓, 胡林, 等. 植物抗蟲“防御警備”:概念、機理與應用[J]. 應用生態(tài)學報, 2018, 29(6): 20682078.
[9] RHOADES D F. Responses of alder and willow to attack by tent caterpillars and webworms: evidence for pheromonal sensitivity of willows [J]. American Chemical Society Symposium Series, 1983, 208: 5568.
[10]PENG J, VAN LOON J J A, ZHENG S, et al. Herbivore-induced volatiles of cabbage (Brassica oleracea) prime defence responses in neighbouring intact plants [J]. Plant Biology, 2011, 13(2): 276284.
[11]ALI M, SUGIMOTO K, RAMADAN A, et al. Memory of plant communications for priming anti-herbivore responses [J/OL]. Scientific Reports, 2013, 3: 1872. DOI: 10. 1038/srep01872.
[12]ERB M, VEYRAT N, ROBERT C A M, et al. Indole is an essential herbivore-induced volatile priming signal in maize [J/OL]. Nature Communications, 2015, 6: 6273. DOI: 10. 1038/ncomms7273.
[13]MUTYAMBAI D M, BRUCE T J A, BERG J V D, et al. An indirect defence trait mediated through egg-induced maize volatiles from neighbouring plants [J/OL]. PLoS ONE, 2016, 11(7): e0158744. DOI: 10. 1371/journal. pone. 0158744.
[14]COPPOLA M, CASCONE P, MADONNA V, et al. Plant-to-plant communication triggered by systemin primes anti-herbivore resistance in tomato [J/OL]. Scientific Reports, 2017, 7(1): 15522. DOI: 10. 1038/s41598-017-15481-8.
[15]LI Tao, BLANDE J D. Associational susceptibility in broccoli: mediated by plant volatiles, impeded by ozone [J]. Global Change Biology, 2015, 21(5): 19932004.
[16]ZHANG Pengjun, WEI Jianing, ZHAO Chan, et al. Airborne host-plant manipulation by whiteflies via an inducible blend of plant volatiles [J]. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(15): 73877396.
[17]KALSKE A, SHIOJIRI K, UESUGI A, et al. Insect herbivory selects for volatile-mediated plant-plant communication [J]. Current Biology, 2019, 29(18): 31283133.
[18]BALDWIN I T, HALITSCHKE R, PASCHOLD A, et al. Volatile signaling in plant-plant interactions: "talking trees" in the genomics era [J]. Science, 2006, 311(5762): 812815.
[19]KARBAN R. Plant behaviour and communication [J]. Ecology Letters, 2008, 11(7): 727739.
[20]MOREIRA X, ABDALA-ROBERTS L. Specificity and context-dependency of plant-plant communication in response to insect herbivory [J]. Current Opinion in Insect Science, 2019, 32: 1521.
[21]張?zhí)K芳, 張真, 王鴻斌, 等. 植物防御的新發(fā)現(xiàn):植物植物相互交流[J]. 植物生態(tài)學報, 2013, 36(10): 11201124.
[22]BRUIN J, DICKE M, SABELIS M W. Plants are better protected against spider-mites after exposure to volatiles from infested conspecifics [J]. Experientia, 1992, 48(5): 525529.
[23]KARBAN A, SHIOJIRI K, ISHIZAKI S, et al. Kin recognition affects plant communication and defence [J/OL]. Proceedings of the Royal Society B: Biological Sciences, 2013, 280(1756): 20123062. DOI: 10.1098/rspb.2012.3062.
[24]ZAKIR A, SADEK M M, BENGTSSON M, et al. Herbivore-induced plant volatiles provide associational resistance against an ovipositing herbivore [J]. Journal of Ecology, 2013, 101(2): 410417.
[25]DICKE M, VAN LOON J J A. Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context [J]. Entomologia Experimentalis et Applicata, 2000, 97(3): 237249.
[26]DICKE M, BALDWIN I T. The evolutionary context for herbivore-induced plant volatiles: beyond the ‘cry for help’ [J]. Trends Plant Science, 2010, 15(3): 167175.
[27]HOLOPAINEN J K, BLANDE J D. Molecular plant volatile communication [J]. Advances in Experimental Medicine and Biology, 2012, 739: 1731.
[28]MESCHER M C, DE MORAES C M. Role of plant sensory perception in plant-animal interactions [J]. Journal of Experimental Botany, 2015, 66(2): 425433.
[29]SIMPRAGA M, TAKABAYASHI J, HOLOPAINEN J K. Language of plants: Where is the word? [J]. Journal of Integrative Plant Biology, 2016, 58(4): 343349.
[30]孫仲享, 宋圓圓, 曾任森. 植物揮發(fā)物介導的種內(nèi)與種間關(guān)系研究進展[J]. 華南農(nóng)業(yè)大學學報, 2019, 40(5): 166174.
[31]NAGASHIMA A, HIGAKI T, KOEDUKA T, et al. Transcriptional regulators involved in responses to volatile organic compounds in plants [J]. Journal of Biological Chemistry, 2019, 294(7): 22562266.
[32]YE Meng, GLAUSER G, LOU Yonggen, et al. Molecular dissection of early defense signaling underlying volatile-mediated defense regulation and herbivore resistance in rice [J]. Plant Cell, 2019, 31(3): 687698.
[33]MEENTS A K, CHEN S P, REICHELT M, et al. Volatile DMNT systemically induces jasmonate-independent direct anti-herbivore defense in leaves of sweet potato (Ipomoea batatas) plants [J/OL]. Scientific Reports, 2019, 9(1): 17431. DOI: 10.1038/s41598019539460.
[34]KESSLER A, BALDWIN I T. Defensive function of herbivore-induced plant volatile emissions in nature [J]. Science, 2001, 291(5511): 21412144.
[35]KOST C, HEIL M. Herbivore-induced plant volatiles induce an indirect defence in neighbouring plants [J]. Journal of Ecology, 2006, 94(3): 619628.
[36]HEIL M. Indirect defence via tritrophic interactions [J]. New Phytologist, 2008, 178(1): 4161.
[37]HARE J D. Ecological role of volatiles produced by plants in response to damage by herbivorous insects [J]. Annual Review of Entomology, 2011, 56(1): 161180.
[38]LI Tao, HOLOPAINEN J K, KOKKO H, et al. Herbivore-induced aspen volatiles temporally regulate two different indirect defences in neighbouring plants [J]. Functional Ecology, 2012, 26(5): 11761185.
[39]TSCHARNTKE T, THIESSEN S, DOLCH R, et al. Herbivory, induced resistance, and interplant signal transfer in Alnus glutinosa [J]. Biochemical Systematics & Ecology, 2001, 29(10): 10251047.
[40]KANG Zhiwei, LIU Fanghua, ZHANG Zhanfeng, et al. Volatile β-ocimene can regulate developmental performance of peach aphid Myzus persicae through activation of defense responses in Chinese cabbage Brassica pekinensis [J/OL]. Frontiers in Plant Science, 2018, 9: 708. DOI: 10. 3389/fpls. 2018. 00708.
[41]SUGIMOTO K, MATSUI K, IIJIMA Y, et al. Intake and transformation to a glycoside of (Z)-3-hexenol from infested neighbors reveals a mode of plant odor reception and defense [J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(19): 71447149.
[42]ERB M. Volatiles as inducers and suppressors of plant defense and immunity-origins, specificity, perception and signaling [J]. Current Opinion in Plant Biology, 2018, 44: 117121.
[43]TIMILSENA B P, SEIDL-ADAMS I, TUMLINSON J H. Herbivore-specific plant volatiles prime neighboring plants for nonspecific defense responses [J]. Plant, Cell & Environment, 2019, 43(3): 787800.
[44]TON J, D’ALESSANDRO M, JOURDIE V, et al. Priming by airborne signals boosts direct and indirect resistance in maize [J]. Plant Journal, 2006, 49(1): 1626.
[45]GERSHENZON J. Plant volatiles carry both public and private messages [J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(13): 52575258.
[46]HEIL M, TON J. Long-distance signalling in plant defence [J]. Trends in Plant Science, 2008, 13(6): 264272.
[47]HEIL M, ADAME-ALVAREZ R M. Short signalling distances make plant communication a soliloquy [J]. Biology Letters, 2010, 6: 843845.
[48]FROST C J, APPEL H M, CARLSON J E, et al. Within-plant signalling via volatiles overcomes vascular constraints on systemic signalling and primes responses against herbivores [J]. Ecology Letters, 2007, 10(6): 490498.
[49]VON MEREY G, VEYRAT N, MAHUKU G, et al. Dispensing synthetic green leaf volatiles in maize fields increases the release of sesquiterpenes by the plants, but has little effect on the attraction of pest and beneficial insects [J]. Phytochemistry, 2011, 72(14/15): 18381847.
[50]ENGELBERTH J, ALBORN H T, SCHMELZ E A, et al. Airborne signals prime plants against insect herbivore attack [J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(6): 17811785.
[51]FROST C J, MESCHER M C, DERVINIS C, et al. Priming defense genes and metabolites in hybrid poplar by the green leaf volatile cis-3-hexenyl acetate [J]. New Phytologist, 2008, 180(3): 722734.
[52]ZEBELO S A, MATSUI K, OZAWA R, et al. Plasma membrane potential depolarization and cytosolic calcium flux are early events involved in tomato (Solanum lycopersicon) plant-to-plant communication [J]. Plant Science, 2012, 196: 93100.
[53]JING Tingting, DU Wenkai, GAO Ting, et al. Herbivore-induced DMNT catalyzed by CYP82D47 plays an important role in the induction of JA-dependent herbivore resistance of neighboring tea plants [J]. Plant, Cell & Environment, 2021,44(4):11781191.
[54]SU Qi, YANG Fengbo, ZHANG Qinghe, et al. Defence priming in tomato by the green leaf volatile (Z)-3-hexenol reduces whitefly transmission of a plant virus [J]. Plant, Cell & Environment, 2020, 43(11): 27972811.
[55]KARBAN R, BALDWIN I T, BAXTER K J, et al. Communication between plants: induced resistance in wild tobacco plants following clipping of neighboring sagebrush [J]. Oecologia, 2000, 125(1): 6671.
[56]KARBAN R, MARON J, FELTON G W, et al. Herbivore damage to sagebrush induces resistance in wild tobacco: evidence for eavesdropping between plants [J]. OIKOS, 2003, 100(2): 325332.
[57]KARBAN R, HUNTZINGER M, MCCALL A C. The specificity of eavesdropping on sagebrush by other plants [J]. Ecology, 2004, 85(7): 18461852.
[58]KARBAN R, SHIOJIRI K, MCCALL H A C. Damage-induced resistance in sagebrush: volatiles are key to intra- and interplant communication [J]. Ecology, 2006, 87(4): 922930.
[59]KARBAN R, SHIOJIRI K, ISHIZAKI S. An air transfer experiment confirms the role of volatile cues in communication between plants [J]. American Naturalist, 2010, 176(3): 381384.
[60]PEARSE I S, HUGHES K, SHIOJIRI K, et al. Interplant volatile signaling in willows: revisiting the original talking trees [J]. Oecologia, 2013, 172(3): 869875.
[61]KOBAYASHI Y, YAMAMURA N. Evolution of signal emission by uninfested plants to help nearby infested relatives [J]. Evolutionary Ecology, 2007, 21(3): 281294.
[62]SEMCHENKO M, SAAR S, LEPIK A. Plant root exudates mediate neighbour recognition and trigger complex behavioural changes [J]. New Phytologist, 2014, 204(3): 631637.
[63]BIEDRZYCKI M L, JILANY T A, DUDLEY SA, et al. Root exudates mediate kin recognition in plants [J]. Communicative & Integrative Biology, 2010, 3(1): 2835.
[64]KARBAN R, SHIOJIRI K. Self-recognition affects plant communication and defense [J]. Ecology Letters, 2009, 12(6): 502506.
[65]SHIOJIRI K, KARBAN R. Plant age, communication, and resistance to herbivores: young sagebrush plants are better emitters and receivers [J]. Oecologia, 2006, 149(2): 214220.
[66]GIRON-CALVA P S, MOLINA-TORRES J, HEIL M, et al. Volatile dose and exposure time impact perception in neighboring plants [J]. Journal of Chemical Ecology, 2012, 38(2): 226228.
[67]STRAUSS S Y, RUDGERS J A, LAU J A, et al. Direct and ecological costs of resistance to herbivory [J]. Trends in Ecology & Evolution, 2002, 17(6): 278285.
[68]HOBALLAH M E, KLLNER T G, DEGENHARDT J, et al. Costs of induced volatile production in maize [J]. OIKOS, 2004, 105(1): 168180.
[69]HEIL M, KARBAN R. Explaining evolution of plant communication by airborne signals [J]. Trends in Ecology & Evolution, 2010, 25(3): 137144.
[70]NINKOVIC V, MARKOVIC D, DAHLIN I. Decoding neighbour volatiles in preparation for future competition and implications for tritrophic interactions [J]. Perspectives in Plant Ecology Evolution and Systematics, 2016, 23: 1117.
[71]KARBAN R, MARON J. The fitness consequences of interspecific eavesdropping between plants [J]. Ecology, 2002, 83(5): 12091213.
[72]BALDWIN I T, KESSLER A, HALITSCHKE R. Volatile signaling in plant-plant-herbivore interactions: what is real? [J]. Current Opinion in Plant Biology, 2002, 5(4): 351354.
[73]PICKETT J A, KHAN Z R. Plant volatile-mediated signalling and its application in agriculture: successes and challenges [J]. New Phytologist, 2016, 212(4): 856870.
[74]ARIMURA G I, MUROI A, NISHIHARA M. Plant-plant-plant communications, mediated by (E)-β-ocimene emitted from transgenic tobacco plants, prime indirect defense responses of lima beans [J]. Journal of Plant Interactions, 2012, 7(3): 193196.
[75]PICKETT J A, WOODCOCK C M, MIDEGA C A, et al. Push-pull farming systems [J]. Current Opinion in Biotechnology, 2014, 26: 125132.
[76]SHULAEV V, SILVERMAN P, RASKIN I, et al. Airborne signalling by methyl salicylate in plant pathogen resistance [J]. Nature, 1997, 385(6618): 718721.
[77]KYUTARO K, KENJI M, RIKA O, et al. Volatile C6-aldehydes and allo-ocimene activate defense genes and induce resistance against Botrytis cinerea in Arabidopsis thaliana [J]. Plant & Cell Physiology, 2005, 46(7): 10931102.
[78]YI H S, HEIL M, ADAME-ALVAREZ R M, et al. Airborne induction and priming of plant defenses against a bacterial pathogen [J]. Plant Physiology, 2009, 151(4): 21522161.
[79]PIESIK D, LEMNCZYK G, SKOCZEK A, et al. Fusarium infection in maize: volatile induction of infected and neighboring uninfected plants has the potential to attract a pest cereal leaf beetle, Oulema melanopus [J]. Journal of Plant Physiology, 2011, 168(13): 15341542.
[80]FARMER E E, RYAN C A. Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves [J]. Proceedings of the National Academy of Sciences of the United States of America, 1990, 87(19): 77137716.
[81]DOLCH R, TSCHARNTKE T. Defoliation of alders (Alnus glutinosa) affects herbivory by leaf beetles on undamaged neighbours [J]. Oecologia, 2000, 125(4): 504511.
[82]KARBAN R. Communication between sagebrush and wild tobacco in the field [J]. Biochemical Systematics & Ecology, 2001, 29(10): 9951005.
[83]KIKUTA Y, UEDA H, NAKAYAMA K, et al. Specific regulation of pyrethrin biosynthesis in Chrysanthemum cinerariaefolium by a blend of volatiles emitted from artificially damaged conspecific plants [J]. Plant & Cell Physiology, 2011, 52(3): 588596.
[84]RENOU A, TRTA I, TOGOLA M. Manual topping decreases bollworm infestations in cotton cultivation in Mali [J]. Crop Protection, 2011, 30(10): 13701375.
[85]MARKOVIC D, COLZI I, TAITI C, et al. Airborne signals synchronize the defenses of neighboring plants in response to touch [J]. Journal of Experimental Botany, 2019, 70(2): 691700.
[86]NINKOVIC V, DAHLIN I, VUCETIC A, et al. Volatile exchange between undamaged plants-a new mechanism affecting insect orientation in intercropping [J/OL]. PLoS ONE, 2013, 8(7): e69431. DOI: 10. 1371/journal. pone. 0069431.
(責任編輯:楊明麗)
