核內(nèi)RNA對真核細胞基因組三維高級結(jié)構(gòu)的影響
2021-01-05 08:40:18金加沈文龍李平張彥陳昭烈趙志虎
生物技術通訊 2020年5期
關鍵詞:結(jié)構(gòu)
金加,沈文龍,李平,張彥,陳昭烈,趙志虎
軍事醫(yī)學研究院 生物工程研究所,北京 100071
細胞核內(nèi)染色質(zhì)在三維空間上的精確折疊包裝和選擇性表達決定了細胞的命運,也為各種染色體活動(如轉(zhuǎn)錄和復制[1-3])的正常進行提供了保障。一維線性染色質(zhì)通過螺旋纏繞和高度折疊最終形成更高級的拓撲相關結(jié)構(gòu)域(topologically associating domains,TAD)和A/B區(qū)室。A區(qū)室主要富集H3K4ac、H3K4me1等活躍的組蛋白修飾標志,存在高密度的基因分布和復制起始位點;相反B區(qū)室富含H3K27me3等沉默的組蛋白修飾標志,分布的基因較少,另外還有晚期復制區(qū)域[2-4]。然而這些結(jié)構(gòu)并不是一成不變的,在分化或疾病發(fā)生時也可能會發(fā)生動態(tài)變化[5-6]。
Isoda等研究發(fā)現(xiàn)核內(nèi)RNA參與了區(qū)室和環(huán)(loop)的形成[7-8],并且Erdel發(fā)現(xiàn)核內(nèi)RNA可以通過“液相分離”[9]來創(chuàng)建局部細胞核環(huán)境,從而促進細胞核內(nèi)無膜細胞器的組裝,進而形成更高度有序的核結(jié)構(gòu),比如核糖體RNA介導的核結(jié)構(gòu)的形成[10-11]。據(jù)報道,核內(nèi)RNA不僅可以參與形成完整的區(qū)室[12-15],還參與形成其他核結(jié)構(gòu)域,如剪接因子富集斑點、核仁等[16-19]。如果將核內(nèi)RNA去除,通過電子顯微鏡進行顯色觀察發(fā)現(xiàn)高度有序的核結(jié)構(gòu)發(fā)生混亂[14,20],所以核內(nèi)RNA在核結(jié)構(gòu)的形成中扮演重要的角色。
為了探究核內(nèi)RNA對染色體三維高級結(jié)構(gòu)的影響,我們用全基因組染色質(zhì)構(gòu)象捕獲(highthroughput/resolution chromosome conformation capture,Hi-C)技術分析了野生型GM12878細胞和經(jīng)RNase處理的GM12878細胞染色質(zhì)的相互作用,以及A/B區(qū)室和拓撲相關結(jié)構(gòu)域的變化。
1 材料與方法
1.1 材料
女性B淋巴細胞GM12878由本室保存。GM12878細胞存在特殊的X染色體失活現(xiàn)象,是研究核內(nèi)RNA對染色體三維結(jié)構(gòu)影響的很好的實驗模型。GM12878細胞已經(jīng)參與了許多組學研究,可為后續(xù)實驗提供參考且便于培養(yǎng)。
RPMI培養(yǎng)基(Hyclone公司);多聚甲醛(國藥集團化學試劑有限公司);RNaseA(50 U/mL)、RNaseT1(20 000 U/mL)、Pro Flex PCR System(Life Technologies公司);RNaseH(5000 U/mL)、HindⅢ-HF(100 U/μL)(NEB公司);T4DNA連接酶(5 Weiss U/μL)(Thermo Scientific公司);超聲破碎儀(VCX-750 SONICS)。
1.2 GM12878細胞的Hi-C實驗
采用Hi-C技術,分別對野生型GM12878細胞和經(jīng)RNase處理的GM12878細胞進行實驗。首先將約6×107GM12878細胞于4℃、500 r/min離心5 min,用2 mL 1×PBS洗滌1次,用4 mL 1×PBS重懸細胞沉淀,將細胞混勻后移入50 mL離心管,在其中加入40 μL 10%甲醛溶液,使其終濃度為1%,在搖床上交聯(lián)10 min,加入終濃度為0.125 mol/L的甘氨酸,混勻后在搖床上終止交聯(lián)5 min,4℃、500 r/min離心5 min,棄上清,用裂解液重懸細胞沉淀后在冰上靜置5 min,再將裂解液移入Dounce管,輕輕上下轉(zhuǎn)動管內(nèi)的磨砂玻璃棒20次,然后將野生GM12878細胞Dounce管中的裂解液直接用5 μLHindⅢ酶切,RNase處理組Dounce管中的裂解液在5 μLHindⅢ酶切的同時加入 5.2 μL RNaseH、13 μL RNaseA/RNaseT1,4℃、500 r/min離心5 min,棄上清,用2 μL KLenow、2.4 μL 10mmol/L dA/T/GTP、2.4 μL 5 mmol/L生物素標記的dCTP、5 μL緩沖液2和39 μL ddH2O補平粘性末端,吹打混勻,在旋轉(zhuǎn)混勻儀上于23℃混勻4 h,補平體系,500 r/min離心3 min,棄上清,加入215 μL ddH2O、25 μL T4DNA連接酶緩沖液洗滌1次,用10 μL T4DNA連接酶于16℃連接8 h,然后用蛋白酶K解交聯(lián),超聲打斷后,進行建庫二代測序。
1.3 數(shù)據(jù)處理與分析
首先將得到的Hi-C數(shù)據(jù)進行質(zhì)控,分別進行文庫過濾統(tǒng)計、文庫比對數(shù)據(jù)統(tǒng)計和文庫有效數(shù)據(jù)過濾統(tǒng)計,在此基礎上進一步處理實驗數(shù)據(jù),分別繪制全基因組相互作用熱圖、全基因組相互作用衰減指數(shù)(interaction decay exponents,IDE)比較分析、差異熱圖比較分析、統(tǒng)計A、B區(qū)室差異并采用GO注釋進行功能分析。此外,對染色體之間的相互作用(秩和檢驗P<0.05)和TAD邊界強度進行比較分析(Vilcox檢驗P<0.05)。
2 結(jié)果
2.1 Hi-C數(shù)據(jù)的質(zhì)量控制
在文庫過濾統(tǒng)計中(圖1A),野生型GM12878末端完全配對片段占比97.92%,經(jīng)RNase處理的GM12878末端完全配對片段占比98.43%,2組文庫的完全配對片段占比較高。文庫比對數(shù)據(jù)統(tǒng)計(圖1B),與參考文庫相比,野生型GM12878文庫惟一配對片段占比74.65%,經(jīng)RNase處理的GM12878文庫惟一配對片段占比73.52%,沒有配對的其他類型片段占比較低,2組文庫樣本質(zhì)量較好。2組文庫有效數(shù)據(jù)過濾統(tǒng)計(圖1C),野生型GM12878文庫相互作用片段占比95.45%,其中有效相互作用片段占比82.09%,在RNase處理的GM12878文庫中,相互作用占比95.09%,其中有效相互作用74.1%,表明Hi-C實驗質(zhì)量較高。
2.2 核內(nèi)RNA對染色體相互作用的影響
野生型GM12878染色體內(nèi)相互作用熱圖(圖2A)和RNase處理后的GM12878染色體相互作用熱圖(圖2B)中的橫坐標和縱坐標代表同一染色體的不同堿基位置,圖中的紅點代表了染色體兩位置之間存在較強的相互作用,紅色越強表示相互作用的強度越大。野生型GM12878和RNase處理的GM12878染色體內(nèi)部的相互作用呈現(xiàn)出一樣的相互作用狀態(tài),都是區(qū)室內(nèi)部的相互作用大于區(qū)室之間的相互作用。
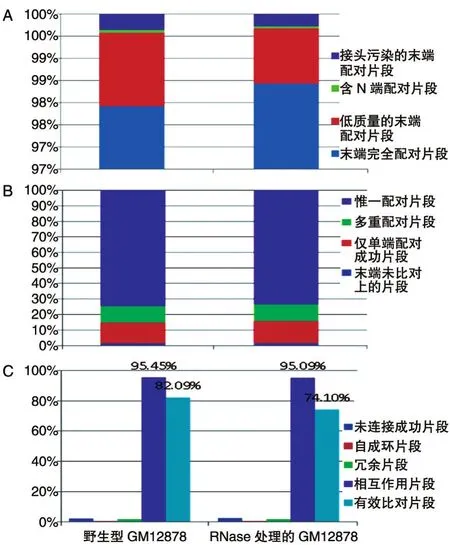
圖1 Hi-C數(shù)據(jù)的質(zhì)量控制
為了比較野生型GM12878和RNase處理的GM12878染色體內(nèi)部的相互作用差異,將野生型GM12878染色體內(nèi)相互作用減扣RNase處理的GM12878染色體內(nèi)相互作用。圖3橫縱坐標分別代表同一染色體上的不同位置,圖內(nèi)紅點代表野生型GM12878相互作用更強,藍點代表RNase處理的GM12878相互作用更強。相比于近距離的染色質(zhì)內(nèi)相互作用,染色體內(nèi)遠距離的相互作用有更多的改變。這也表明核內(nèi)RNA在遠距離的相互作用上起到了更重要的作用。
我們繪制了全基因組IDE曲線(圖4),隨著基因距離的不斷增大,野生型GM12878和RNase處理的GM12878細胞的相互作用頻率由10-6下降到10-8,其中RNase處理組在基因距離108堿基對處的相互作用頻率略高于野生型GM12878,說明相比于野生型GM12878細胞,經(jīng)RNase處理后的GM12878細胞的染色體遠距離相互作用有所增強,染色體的結(jié)構(gòu)變得松散。
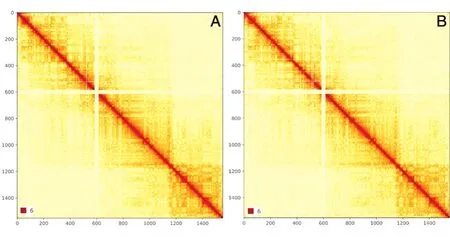
圖2 X染色質(zhì)相互作用熱圖

圖3 染色質(zhì)內(nèi)相互作用的減扣
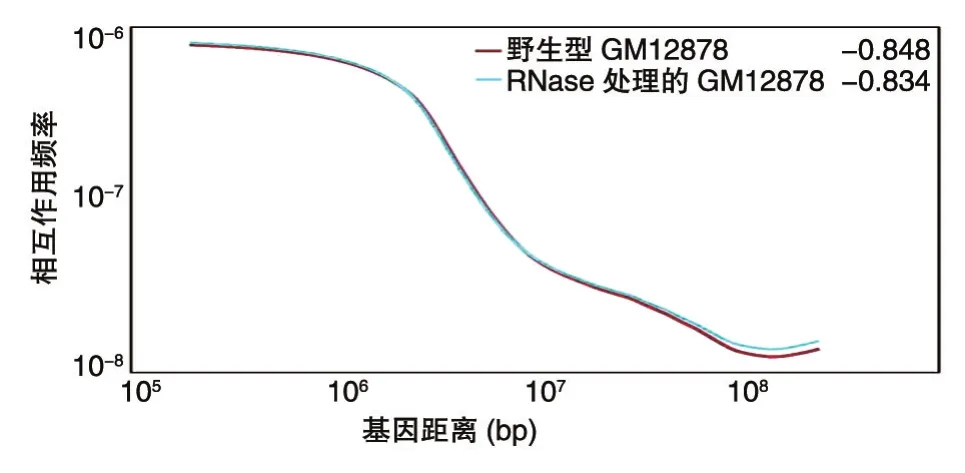
圖4 染色質(zhì)相互作用衰減指數(shù)
除此之外,我們還比較了2組細胞的染色體之間的相互作用(圖5),RNase處理的GM12878細胞染色體間相互作用分數(shù)高于野生型GM12878細胞(秩和檢驗P<0.05)。這也說明經(jīng)過RNase處理的GM12878細胞的染色質(zhì)結(jié)構(gòu)變得松散。
2.3 核內(nèi)RNA對A/B區(qū)室的影響
為了進一步分析核內(nèi)RNA對區(qū)室的影響,我們統(tǒng)計了2組GM1287細胞在A/B區(qū)室上的變化。所有染色體A/B區(qū)室轉(zhuǎn)變比例餅圖(圖6A)中,A區(qū)室轉(zhuǎn)變?yōu)锽區(qū)室占比0.51%,B區(qū)室轉(zhuǎn)變?yōu)锳區(qū)室占比0.62%,未發(fā)生變化的A區(qū)室占比48.38%,未發(fā)生改變的B區(qū)室占比50.48%。經(jīng)比較發(fā)現(xiàn),B區(qū)室轉(zhuǎn)變?yōu)锳區(qū)室所占比例高于A區(qū)室轉(zhuǎn)變?yōu)锽區(qū)室。將發(fā)生轉(zhuǎn)變區(qū)室上的基因進行GO注釋功能分析,發(fā)現(xiàn)在轉(zhuǎn)變區(qū)塊中,A區(qū)室轉(zhuǎn)變成B區(qū)室上的基因在功能上主要和脫氫酶相關,B區(qū)室轉(zhuǎn)變成A區(qū)室上的基因在功能上主要和細胞外基質(zhì)結(jié)構(gòu)成分相關。在X染色體A/B區(qū)室轉(zhuǎn)變比例餅圖(圖6B)中,A區(qū)室轉(zhuǎn)變?yōu)锽區(qū)室占比1.45%,B區(qū)室轉(zhuǎn)變?yōu)锳區(qū)室占比1.45%,未發(fā)生變化的A區(qū)室占比47.98%,未發(fā)生改變的B區(qū)室占比49.12%,所以相比于所有染色體的變化,X染色體上發(fā)生變化的A/B區(qū)室的比例更高。由于GM12878是女性淋巴細胞,含有2條X染色體,其中一條由于核內(nèi)非編碼RNA Xist的緊密纏繞而處于壓縮狀態(tài),我們認為RNase的處理導致緊密的X染色體變得松散,所以相比于所有染色體A/B區(qū)室的變化,X染色體的A/B區(qū)室變化更大。對編碼Xist基因的74M左右區(qū)域進行染色體相互作用分析(圖7),野生型GM12878細胞Xist基因位置的相互作用減扣RNase處理的GM12878細胞Xist位置的相互作用顯示藍色增多,說明相比于野生型GM12878細胞,RNase處理后GM12878細胞的相互作用更弱。
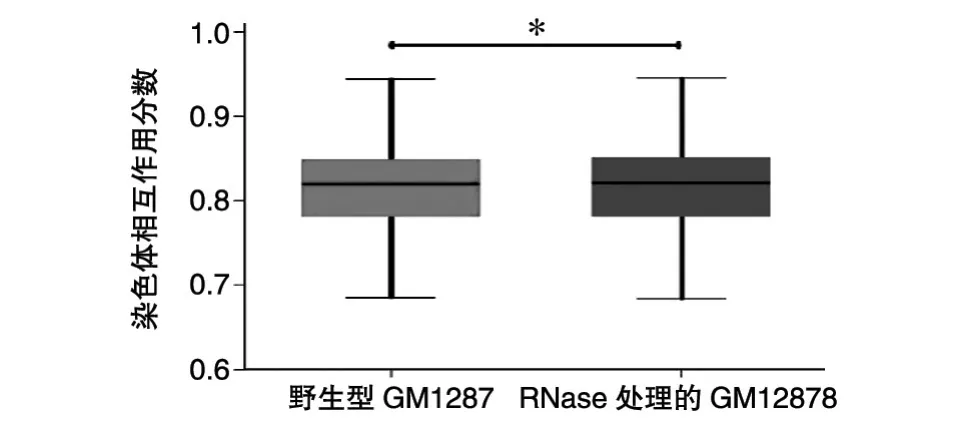
圖5 染色質(zhì)之間的相互作用比較(*P<0.05)
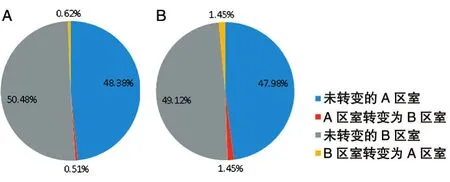
圖6 A/B區(qū)室轉(zhuǎn)變比例
2.4 核內(nèi)RNA對TAD邊界強度的影響
為了觀察RNase處理后TAD邊界強弱的變化,將野生型GM12878和RNase處理的GM12878細胞的TAD邊界的強弱進行比較(圖8),發(fā)現(xiàn)相比于野生型GM12878細胞,經(jīng)RNase處理的GM12878細胞的TAD邊界強度較小(Vilcox校驗P<0.05)。這是因為由于核內(nèi)RNA參與了loop形成,經(jīng)RNase處理,導致環(huán)結(jié)構(gòu)改變,進而影響TAD結(jié)構(gòu)變得松散,從而減弱了TAD邊界強度。
3 結(jié)論
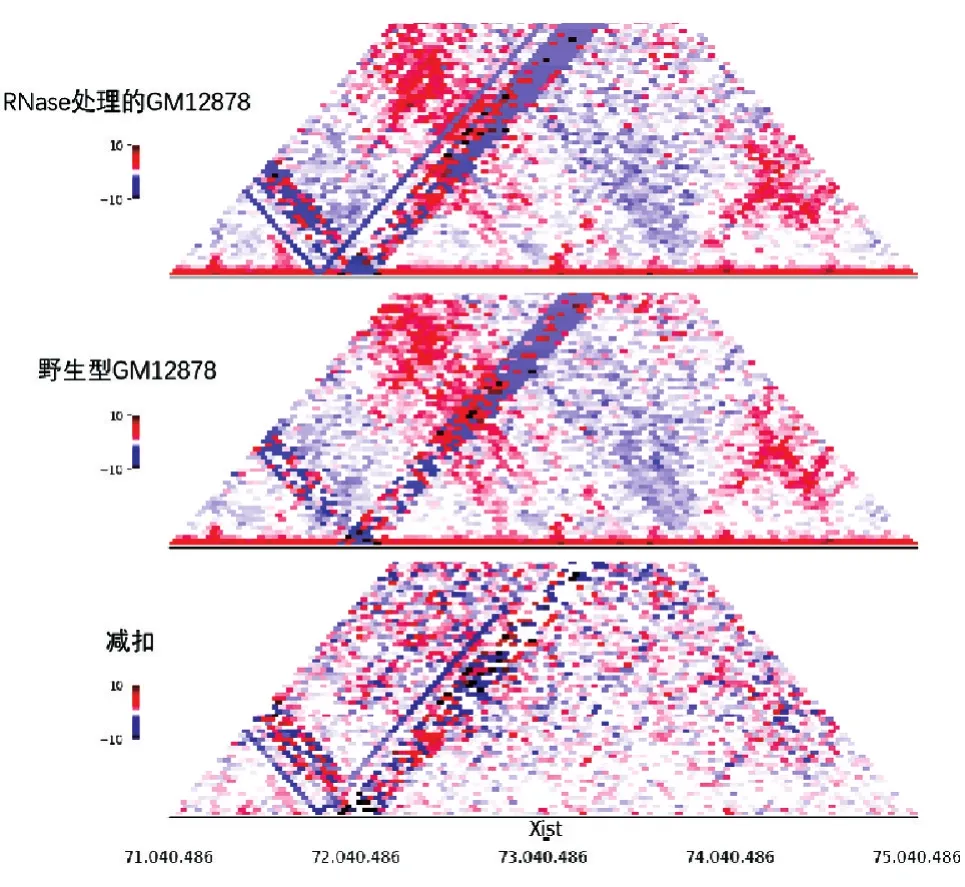
圖7 X染色體74M左右區(qū)域相互作用熱圖
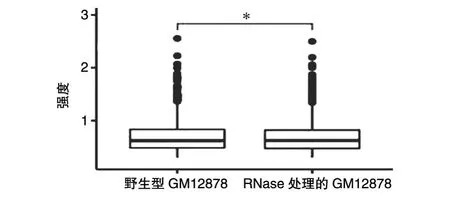
圖8 TAD邊界強弱變化
我們證實了核內(nèi)RNA對真核細胞染色體三維高級結(jié)構(gòu)是有影響的,RNA的去除導致真核細胞染色體三維高級結(jié)構(gòu)在整體上變得松散,A/B區(qū)室發(fā)生改變,目前已經(jīng)發(fā)現(xiàn)RNA可以通過結(jié)合RNA結(jié)合蛋白對染色質(zhì)進行修飾,進而調(diào)控染色質(zhì)環(huán)的形成,核內(nèi)RNA還參與了TAD邊界的形成和建立,而RNase處理導致了環(huán)結(jié)構(gòu)的破壞,TAD邊界強度減弱。基因組染色體三維結(jié)構(gòu)的穩(wěn)定性是表觀遺傳的關鍵,受到許多物質(zhì)的調(diào)控,如核內(nèi)RNA、蛋白質(zhì)、激素等。如果基因組的三維結(jié)構(gòu)或者調(diào)控出現(xiàn)了偏差,有可能導致疾病。我們認為在未來會研發(fā)出一些更好的分子生物實驗,特別是研究核內(nèi)RNA對染色質(zhì)三維高級結(jié)構(gòu)和染色質(zhì)修飾物影響的實驗,這對于揭示基因組三維高級結(jié)構(gòu)形成的潛在原理至關重要。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數(shù)理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
