北部灣大風江與南流江河口紅樹林空間分布格局研究
2021-01-15 08:17:00王日明戴志軍黃鵠梁喜幸黎樹式胡寶清周曉妍吳天亮
海洋學報 2020年12期
王日明,戴志軍,黃鵠,梁喜幸,黎樹式,胡寶清,周曉妍,吳天亮
( 1. 北部灣大學 廣西北部灣海岸科學與工程實驗室/欽州市環境生態修復重點實驗室,廣西 欽州 535011;2. 華東師范大學 河口海岸學國家重點實驗室,上海 200062;3. 南寧師范大學 北部灣環境演變與資源利用教育部重點實驗室,廣西南寧 510310)
1 引言
紅樹是生長在熱帶亞熱帶河口或海岸潮間帶的耐鹽性木本植物,在漁業、海岸保護、營養鹽循環及捕獲碳通量等方面具有極其重要的生態服務價值[1–2]。因全球海平面上升和高強度人類活動的作用,沿海紅樹林海岸遭受侵蝕,紅樹林面臨大規模消失的風險[2]。紅樹林變化、紅樹林潮灘沖淤以及紅樹林生境及生態服務等已成為眾多學者和相關政府關注的重大理論與社會問題[1,3–5]。較多研究針對前述問題開展了相關工作[1–6],對推動紅樹林生態、生理及生境和響應全球變化研究具有重要價值。與此同時,作為影響紅樹能否發育與成活的關鍵因子,鹽度不僅脅迫紅樹組織結構,影響抗氧化酶活性,而且直接控制紅樹林種屬地域分布特征[7–12]。故在影響紅樹生理與形態方面的環境因子研究中,有不少是鹽度對紅樹林的影響研究[8–9]。譬如,受潮汐淹水造成的鹽度在潮灘分布不一,紅樹植物在潮間帶分布呈現出低灘帶、中潮帶和高潮帶帶狀分布特征[3]。河口水體鹽度的規律性變化成為影響紅樹植物沿入海河段分布的限制因子[7,10–11]。此外,一些研究指出桐花樹(Aegiceras corniculatum(L)Blanco)作為先鋒紅樹植物的一種,鹽度會影響其幼苗萌發和生長的進程[12],其最適宜海水鹽度為8~15[13],桐花樹在低鹽度下生長量最大,在中等鹽度海灘可以種植桐花樹[14]。然而,到目前為止一直較少有研究涉及入海河口段紅樹林自海向陸是如何分布以及不同因素在其間扮演的角色,這在很大程度上影響了紅樹林宜林地選擇與國家“南紅北柳”—“南紅”重大生態工程的開展。基于此,本文以北部灣大風江和南流江為代表,研究北部灣入海河口紅樹林沿程空間分布格局,探討外部因素對紅樹林的影響過程,旨在揭示區域入海河口紅樹林分布狀態及驅動機制,在理論上補充河口段紅樹林發育與演化體系,在應用上為“南紅”工程與生態堤防建設提供技術支撐。
2 研究區域概述
大風江和南流江同屬于北部灣山溪型入海河流(圖1)。該區域位于北熱帶季風氣候區,夏季高溫多雨,冬季溫和少雨,雨熱同期。年降水量為1 693 mm,降雨主要集中在6?9 月[15],10 月至翌年5 月為枯水期。潮汐屬于不規則全日潮[16],受潮汐影響的南流江入海河口發育面積約550 km2的三角洲,主入海河段16 km,河床落差達3.34‰。河口段河岸多處修建有人工河堤,自海向陸河口段形成的潮灘由發育相對平緩到坡度較陡,呈狹長的斷續帶狀分布。受控于周邊地形,大風江河口段形成深入內陸的溺谷海灣環境。河口上游東場構建有擋潮閘,導致徑流較少過閘入海,僅沿線匯流區內降水進入該區域。河岸坡度較陡,河口潮灘自海向陸由發育稀疏過渡到狹長而呈斷續帶狀分布特征。
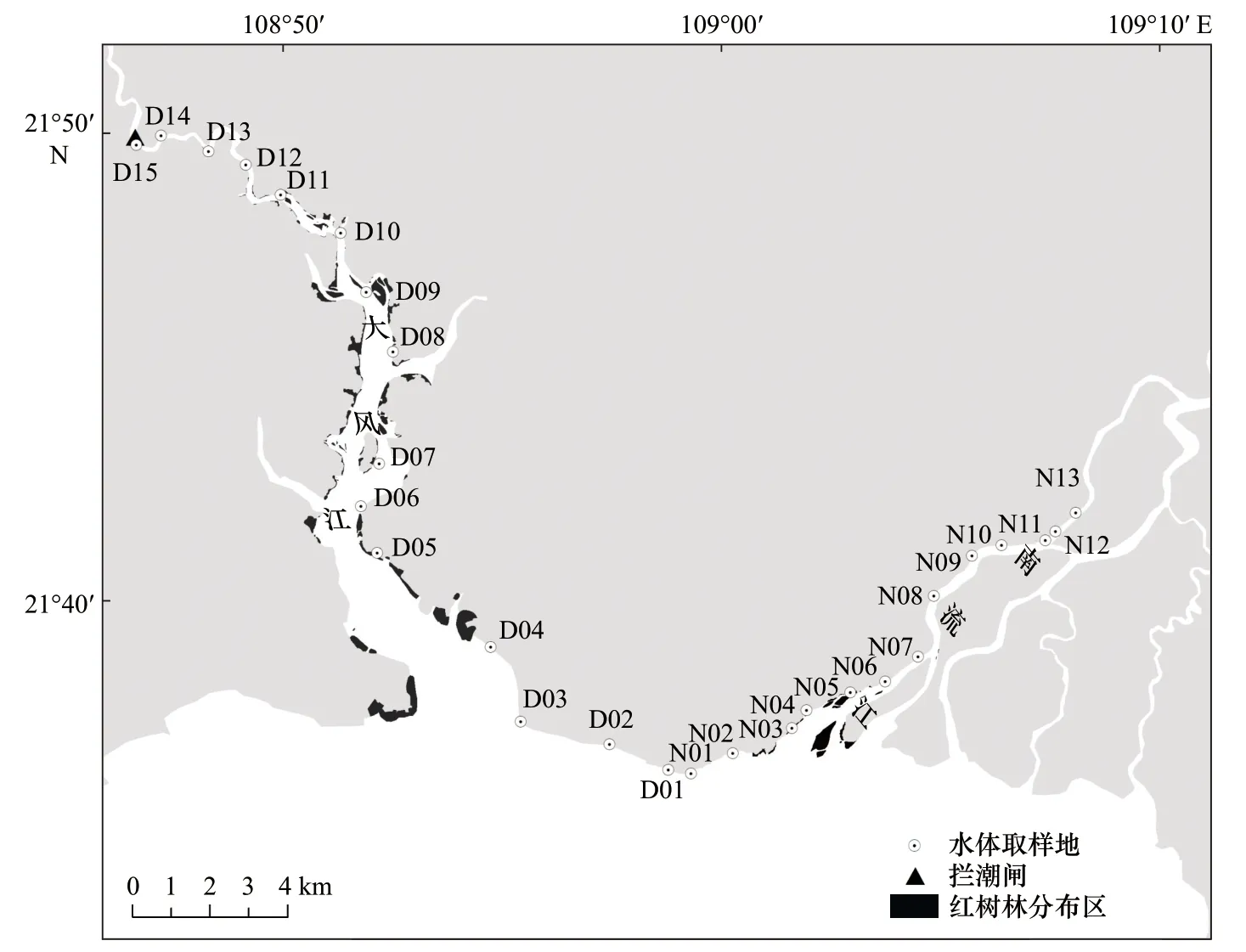
圖1 研究區域紅樹林分布與采樣站點Fig. 1 Mangrove distribution and survey stations of the research area
3 實驗設計與研究方法
為研究兩個河口段自海向陸紅樹林的空間分布特征及其耐鹽性,實驗分為室內和野外兩個階段。其中室內實驗選擇本地河口區優勢種桐花樹胚胎進行不同鹽度環境的沉水測試。野外實驗則選擇在冬季大潮高潮期間沿河口向陸地測定不同位置水體表層鹽度,同時在低潮期間調查所測位置河口潮灘紅樹林分布及灘地坡度等。
3.1 桐花樹胚胎沉水實驗
3.1.1 實驗材料
(1)實驗所用桐花樹胚胎來自南流江河口七星島島尾潮灘桐花樹種群。這些胚胎全部是當地生長健壯的成年樹樹冠外圍中上部向陽面的成熟胚胎。胚胎采集時間為2019 年9 月22 日。
(2)實驗所用不同鹽度配比水體:不同鹽度水體包括0、20、30、40、50,其中鹽度為0 實驗用水為靜放24 h 的自來水,其他鹽度用水采用從實驗區內所取鹽度為10.9 的天然海水加海鹽與經靜放24 h 的自來水調配。不同鹽度水體均用Pocket PAL-ES3 鹽度計標定鹽度,用PZ-D-5 型液體比重天平標定比重。
3.1.2 實驗設計
于 2019 年9 月23 日08:00 把從實驗區采回的供試新鮮胚胎進行浸泡下沉實驗。以鹽度為0 的自來水浸泡為對照,分別設鹽度為20、30、40、50 共4 個處理,每個鹽度處理設5 個重復,每組樣品為10 顆桐花樹胚胎。為減少水分蒸發導致的水體鹽度升高,保證桐花樹胚胎所處水體鹽度恒定在所設計鹽度,每間隔2 d 對各鹽度水體進行標定、換水。每天8:00、10:00、12:00、14:00、16:00、18:00、20:00、22:00 定 時觀測和記錄紅樹林胚胎吸水下沉數量與下沉時間。
3.2 河口實地實驗
為較準確地反映河口紅樹林的空間分布及生境特性,本文選擇在枯季徑流最弱期間、大潮高潮漲平階段進行水體采樣,在大潮低平期對沿岸潮灘紅樹進行調查。枯季大潮高潮漲平階段河口鹽水上溯抵達位置最遠,即枯季河口潮區界所在。水樣采集站分別沿南流江主入海河口和大風江河口河岸自口外海濱到潮區界布設,調查取樣點分別為13 個及15 個(圖1)。同時利用RTK 對各個站點之間的坐標進行量測,隨后基于坐標計算得到各站點之間的距離。
3.2.1 河口表層水體鹽度采樣設計
(1)南流江入海口河段鹽度調查取樣:為了取得枯水季節大潮高潮位期間漲潮水流對南流江河口段水體鹽度的影響,本實驗于2019 年12 月15 日05:58?10:17 測定各站點(圖1)表層水體鹽度。為較完整地記錄高潮位鹽度變化特征,于06:00 在各站點水面與灘地交界處插設標桿,標桿上每隔5 cm 貼標記,以記錄站點水位上升、下降情況。漲潮時水面每上升5 cm時取一次水樣,直到達到水位最高點;隨后落潮時每下降5 cm 時取一次水樣,連取3~5 個水樣。把所取水樣帶回實驗室經過充分攪拌后用Pocket PAL-ES3鹽度計測定各水樣鹽度。
(2)大風江河口河段鹽度調查取樣:實驗分別在2019 年12 月29 日和30 日漲潮至最高潮位期間駕船往返于1~15 號點,每隔5 min 取一次水樣帶回實驗室測定河水鹽度。
3.2.2 河口紅樹林植物分布調查取樣
紅樹林植物沿河口向上游分布具有明顯差異,為便于研究和比較,本文將河口潮間帶植被定性歸納為5 類。
Ⅰ類:紅樹植物純林(含多種紅樹植物混交林)區段,紅樹植物在河口段沿岸潮灘地大范圍分布,極少有非紅樹植物混雜。
Ⅱ類:紅樹植物大斑塊區段,紅樹植物在河口段沿岸潮灘地聚集組團分布形成紅樹植物大斑塊,紅樹植物大斑塊間有半紅樹植物、非紅樹斑塊,但以紅樹植物斑塊為主。
Ⅲ類:局部紅樹植物混交林區段,有成年紅樹植物小斑塊鑲嵌于其他非紅樹混交林中,以非紅樹植物為主。
Ⅳ類:有一二年生紅樹植物小苗區段,但未發現成年紅樹植物。
Ⅴ類:有紅樹植物胚胎區段,潮灘地發現有紅樹植物胚胎,但未發現紅樹植物小苗與成年樹。
基于上述標準,實驗在2019 年12 月29?30 日對南流江和大風江入海河口河段沿岸紅樹植物分布狀況采用按站點分區段普查方法進行現場調查。其中,按上述13 個站點把南流江主入海河口21.3 km 河段從口外海濱上溯,分成12 區段,按上述15 個站點把大風江河口38.1 km 河段從海向陸上溯,分成14 個區段(圖1)。由此分別統計低潮時兩個河口各區段潮灘出現的紅樹植物的分布狀況。
4 結果分析
4.1 桐花樹胚胎浸水下沉對鹽度脅迫的響應
所采回供實驗用桐花樹成熟新鮮胚胎比重為0.923,置于鹽度為0 的水中,漂浮于水面,有部分懸浮于水中。隨后用鹽度為20、30、40、50 的人工鹽水對桐花樹胚胎浸泡處理,用鹽度為0 的水作為對照,觀測胚胎在不同的鹽度水體中吸水下沉時間與數量,結果見圖2。
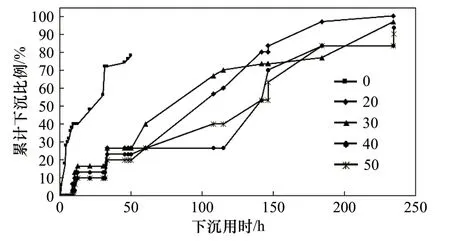
圖2 新鮮桐花樹胚胎對不同鹽度脅迫的響應Fig. 2 The stress response of the fresh Aegiceras corniculatum embryos to different salinity condition
圖2 表明,新鮮的桐花樹胚胎經不同鹽度的人工海水處理,隨著浸泡時間的延長出現不同的變化特征。其中,桐花樹胚胎在低鹽度水體中下沉速度明顯快于高鹽度水體(圖2,表1)。具體而言,在0~10 h時段內,對照組有約25%胚胎下沉,而50 鹽度處理組沒有下沉;20、30、40 鹽度處理組有不同程度下沉,但差異不顯著。10~30 h、30~50 h 時段內,各處理組中30 鹽度處理組下沉最快,50 鹽度處理組下沉最慢,分別為15%、25%與9%、18%。到第50 h 時,對照組已全部下沉,對照組與各處理組表現出顯著差異。到第230 h 時,20 鹽度處理組已全部下沉,但30、40、50 鹽度的處理組仍有約15%胚胎處于漂浮(懸浮)狀態,表明桐花樹胚胎在高鹽度水體中漂浮(懸浮)時間更長。
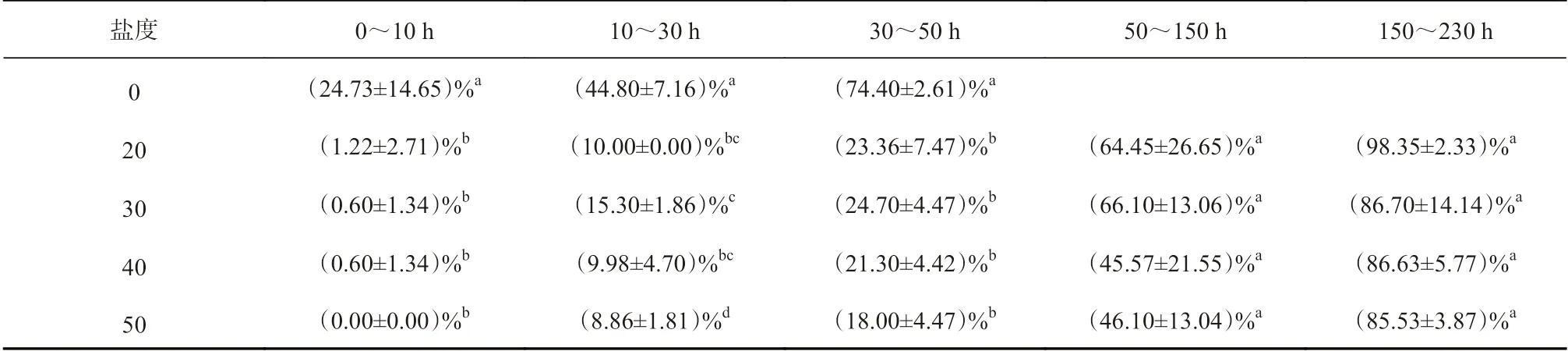
表1 不同鹽度水體中新鮮桐花樹胚胎在不同時段內吸水下沉差異性分析Table 1 Difference analysis for water absorption of the fresh Aegiceras corniculatum embryos in different salinity conditions during the different periods
4.2 入海河口段水體鹽度空間分布
4.2.1 南流江河口段鹽度空間分布特征
枯水期大潮漲平階段,潮汐向河段內推進約15 km,其中口外海濱N01?N03 最高鹽度在23 以上,鹽度最高達25.8,平均鹽度為23.2,潮區界左右的區段鹽度平均為3.6。向海距離越遠,鹽度值越大,向陸距離越遠,鹽度值越小(圖3)。
4.2.2 大風江河口段鹽度空間分布特征
類似地,大風江河口口外海濱D01?D04 最高鹽度在26 以上,向海距離越遠,鹽度值越大,向河內距離越遠,鹽度值越小。然而,與南流江河口有所差異,向陸到33.88 km 附近,其鹽度仍高達15.8,平均鹽度為10.5(D15)。有必要指出的是,D15 恰好位于大風江河流東場攔潮閘下,因閘的阻攔作用,進入閘下淡水徑流極少,導致潮汐作用最遠距離為33.9 km,且鹽度偏高(圖3)。
4.3 河口潮灘紅樹林植物空間分布特征
4.3.1 南流江河口潮灘地紅樹植物群落空間分布
南流江干流河口紅樹林面積為109.13 hm2(圖1)。南流江主入海河段潮灘紅樹植物分布最集中的河段為N03?N05,沿線3.5 km 的河岸潮灘與江心洲滿布紅樹植物秋茄(Kandelia candel(Linn.))與桐花樹,另有少量的無瓣海桑(Sonneratia apetala)、海漆(Excoecaria agallochaLinn.)分布;距N03 站點口外的N01?N03沿線約4.0 km 的海濱潮灘分布有大量的互花米草與秋茄片林;距N03 站點口內3.5 km 的N05?N06 沿線1.5 km 的潮灘,桐花樹生長良好,但有部分半紅樹植物(黃槿(Hibiscus tiliaceus)等)與非紅樹植物混雜;N06?N07 河段潮灘內偶布有桐花樹成年苗,再入上游幾乎沒有成年苗;N12 河段潮灘為桐花樹分布上限,只有少量的一二年生桐花樹小苗分布。總而言之,南流江口潮灘表現出離河口距離越遠,紅樹植物分布越少,長勢越差的趨勢(圖4A)。
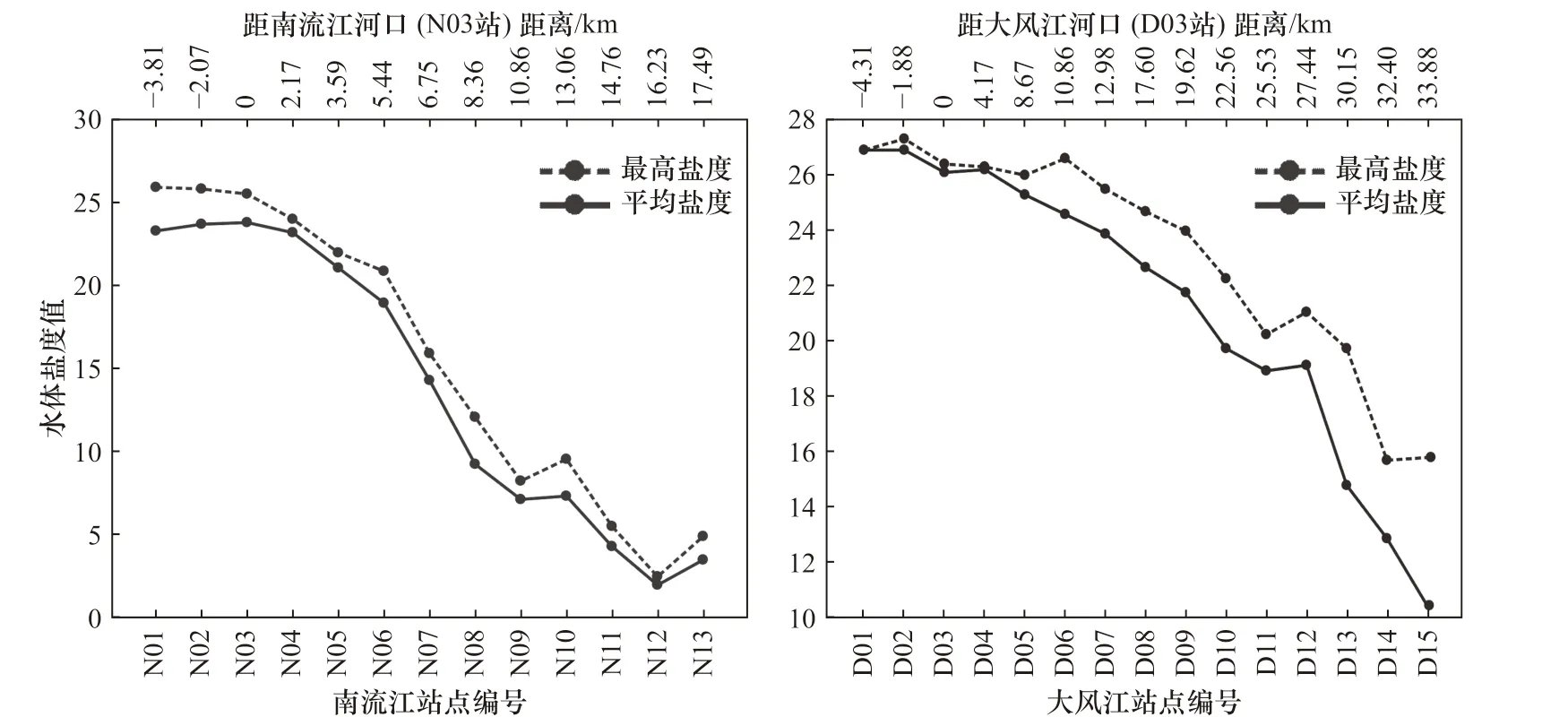
圖3 觀測站點鹽度Fig. 3 The salinity of the observed sites
4.3.2 大風江河口潮灘地紅樹植物群落空間分布
大風江河口紅樹林面積為620.62 hm2(圖1)。大風江入海河口段潮灘紅樹植物分布基本表現出與南流江河口相似的趨勢。D03?D10 河岸潮灘與江心洲為純紅樹林,其中秋茄、桐花樹為絕對優勢種,高潮位以上有黃槿、海漆與苦郎(Clerodendrum inerme (L.) )等半紅樹植物;距D03 站點口外的D01?D03 沿線約4.5 km 的海濱潮灘大多為光灘,有少量的海欖雌(Avicennia marina)與桐花樹一年生苗分布;距D03 站點口內22.0 km 的D10?D12 沿線5.0 km 潮灘與江心洲皆以桐花樹為優勢種群,間或有半紅樹植物(黃槿等)與非紅樹植物;距D03 站點口內27 km 以上的D12?D15 桐花樹成年苗三五成群鑲嵌于半紅樹植物、非紅樹植物之間形成混交林(圖4B)。然而,相比南流江入海口,大風江入海口紅樹林分布范圍更廣,沿河上溯距離遠達距海口33.88 km 的D15 站點攔潮閘(圖4B),且向陸界限主要以紅樹、紅樹幼苗及半紅樹混生為主,向陸界限不明顯。
5 討論
受潮汐強度、堤岸坡度、灘地高程等因素的影響及人為干擾作用,入海河口段沿岸不同潮灘地紅樹林植物的分布可能存在不同差異。
5.1 漲潮時間與胚胎吸水下沉時間
桐花樹胚胎浸水實驗表明,桐花樹胚胎在低鹽度的水中浸泡將吸水下沉,同時下沉的數量又和其浸泡時間相關(圖2)。在相對適宜鹽度的水體中,宜林潮灘桐花樹胚胎成熟脫落后,可懸浮于水體且隨漲潮水動力向河口內陸延伸。基于此,在非整個全日潮影響下,漲潮12 h 內,南流江河口的紅樹胚胎能被帶到受潮汐作用影響的河段最上游N13 段(圖4A)。基于先前研究關于河流受潮汐影響的上限是紅樹植物沿河分布的上限[17]表明,如果在該上限區有合適的潮灘,胚胎落在潮灘上可以生根發育。然而,當沒有適宜潮灘時,紅樹胚胎在河床底著地后,將隨著落潮又被河水沖刷返回下游,繼續隨波逐流。故南流江河口上限僅發現少量的一二年生桐花樹小苗分布。紅樹植物分布的最上限很可能由潮汐周期漲潮上溯時長與胚胎吸水下沉時間決定。此外,在鹽度為40、50 的水體中浸泡230 h,仍有部分胚胎懸浮于水體中表明,桐花樹胚胎在高鹽度水體中懸浮時間長,這為桐花樹胚胎在落潮時隨海水回流而遠距離傳播提供了可能。
5.2 潮灘地坡度
桐花樹胚胎培育實驗發現,胚胎在完全淹水條件下不進行生根發育過程。除受淹水影響,灘地也可能是限制性因子。基于調查中發現的南流江河口段N06?N08 較陡灘地極少出現桐花樹胚胎與一二年生幼苗,而在N09?N12 河段緩灘發現有一二年生幼苗,在N12?N13 低潮位出露的灘涂緩灘發現有胚胎。這就表明,當桐花樹胚胎被漲潮時的潮水推進到河段上游,如果胚胎停留在過陡且有潮灘的位置,落潮后胚胎并不能有效停留在灘地,而將在落潮水動力作用下帶回下游。從而胚胎不能在相對較陡的灘地有效停留。在大風江河口的D10?D15 河段亦有類似情況,外緣沒有適宜于胚胎停留的潮灘地,而僅在現有桐花樹林下有少量一二年生幼苗出現。因而,入海河口段適宜坡度的宜林潮灘地是紅樹植物正常生長發育的必要條件。
5.3 水體鹽度與潮水浸淹時間
桐花樹胚胎在鹽度為10 以下時,生根發育狀況最優,桐花樹對低鹽度表現出很強的適應性[18]。南流江河口的現場調查表明,紅樹植物長勢最好的河段為位于河口 的N03?N05 段,在N01?N02 紅 樹 長勢最差。產生這一現象的原因在于:N01?N02 區域,水體鹽度長期大于23,處于高鹽度水平,桐花樹胚胎萌發率極低[12],故紅樹植物極少,以光灘為主。N03?N05區域地處南流江河口徑潮流交匯頻繁作用的地帶,全年鹽度不僅在10 左右,而且發育相對較大的潮灘給紅樹提供了棲息場所。本研究所調查鹽度為南流江河口枯水季大潮漲潮期間的水體表層鹽度,故鹽度相對偏高,并不能代表該河段全年平均鹽度水平。
然而,大風江河口D03?D10 區段紅樹長勢最好,所測鹽度為20 以上,且由于沒有河水沖淡的作用,將常年維持這一鹽度水平,但紅樹植物依然長勢最好。其主要原因在于河岸潮灘與江心洲紅樹植物生長范圍處于中高潮位,受潮汐浸淹時間較短,而較少受到高鹽度水體影響。其中低潮位與低低潮位沒有紅樹植物生長正解釋了這一現象,即長期被高鹽度水體浸泡,胚胎不能萌根,從而沒有小苗發育。同時,大風江河口上段在東場建有擋潮閘,正常時段閘外水位高于閘內水位,常年沒有淡水補充,該河段相當于一個較深入內陸的狹長海灣,致使河段內鹽度嚴重偏高。經現場實測,在距離海口33.88 km 的擋潮閘處D15 站點的最高鹽度為15.8,平均鹽度為10.5,這就導致大風江口紅樹林分布上限比南流江口遠,分布范圍比南流江口廣(圖1)。這也是紅樹植物分布沿兩條河河口段分布有明顯不同的主要原因。故水體鹽度與潮水浸淹時間是決定河段紅樹種類的關鍵因子。
6 結論
受控于河口復雜的水文與地貌條件,河口紅樹林空間分布較海岸復雜。本文結合室內鹽度實驗,通過對北部灣大風江和南流江入海河口紅樹林及棲息環境的分析,主要結論包括:
(1)南流江和大風江河口紅樹林自海向陸都展現出距離河口越遠,紅樹群落趨于簡單和長勢較差的格局:紅樹林純林(或桐花樹、秋茄、無瓣海桑種類混生)→紅樹植物與半紅樹植物(黃槿、苦朗等)混生→紅樹植物、半紅樹植物與非紅樹植物混生→紅樹植物鑲嵌→紅樹林小苗。
(2)兩個河口水體鹽度自海向陸遞減直接控制紅樹的分布格局,潮水浸淹時間是影響紅樹胚胎發育的關鍵因素,漲潮上溯時長決定了紅樹向陸生長的極限位置,河口段適宜坡度的宜林潮灘地為紅樹植物正常生長發育的必要條件。
(3)大風江口向陸構建攔潮閘,導致閘下鹽度常年高于南流江,從而導致大風江口相較南流江河口的紅樹沒有明顯向陸界限,且主要分布紅樹、紅樹幼苗與半紅樹等。
猜你喜歡
歌海(2024年6期)2024-03-18 00:00:00
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
幼兒園(2021年4期)2021-07-28 07:38:04
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
歌海(2018年4期)2018-05-14 12:46:15
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
