均腐土不同亞綱典型土系土壤細菌群落多樣性研究
2021-01-18 04:20:50呂淑敏張東萌王光華
中國土壤與肥料 2020年6期
關鍵詞:研究
焦 峰,呂淑敏,唐 雷,張東萌,王光華
(1.黑龍江八一農墾大學,黑龍江 大慶 163319;2.中國科學院東北地理與農業生態研究所, 黑土農業生態重點實驗室,黑龍江 哈爾濱 150081)
均腐土(Isohumisols)是中國土壤系統分類(CST)體系中提出并使用的土綱級分類單元,指具暗沃表層、均腐殖質特性和土體鹽基飽和度≥50%的一類土壤,其中均腐殖質特性(Isohumic property,Ihp)是最早由法國發生土壤學及土壤制圖委員會提出并用于土綱級的劃分,CST引入并在高級分類研究階段采用了這個指標,并限定Ihp土表至20 cm與土表至100 cm土層腐殖質儲量比值≤0.4。Isohumosols檢索應首先具有暗沃表層,因此可以理解Isohumosols與美國的Mollisols有大致相似的特點,但因診斷特性Ihp的引入,使Mollisols中大部分不具備Ihp的Albolls很難實現均腐土土綱級的檢索,致使均腐土與Mollisols以及世界土壤資源參比基礎的Phaeozems、俄羅斯的Chernozems、中國土壤發生分類(GSGC)的Black soil有很大的不同,同時,均腐土又是我國重要的土壤類型[1],在東北平原多有分布。CST將均腐土分為干潤均腐土、濕潤均腐土和巖性均腐土3個亞綱[2-3],其中巖性均腐土分布在中國南海諸島[1],而東北平原因其獨特的氣候環境和地理位置主要發育干潤均腐土和濕潤均腐土兩個亞綱,并成為均腐土的主要類型。在均腐殖質特性的環境中生存的細菌形成了特有的結構和機能,而均腐土不同亞綱典型土系的土壤組分可能成為驅動土壤細菌差異的重要因素。近年來,我國學者對于CST體系下均腐土的研究較少且集中于理化指標、土壤質量評價等方面[4-5],對于均腐土生物學性質的變化情況還未涉及。
土壤微生物是生態系統的重要組成部分,在土壤有機質的分解、腐殖質的形成及土壤養分的轉化和循環等生物化學過程中發揮著重要作用[6-9]。土壤微生物對土壤條件變化非常敏感,能在較短時間內發生大幅度變化,因而土壤微生物量的變化通常作為土壤質量演變的重要依據[10-12]。土壤微生物是否存在與植物和動物等大型生物相似或不同的地理分布格局,以及哪些因素驅動著這種格局的存在是當前學者們研究的交叉熱點課題之一。GSGC體系下土壤微生物群落結構地理分布規律與化學性質關系及其驅動因素已有研究而結果不盡相同[13-16],同時,以往的研究均以固定的土壤深度為對比單元,忽略了成土過程和成土條件對土壤性質的影響,很難與土壤本質屬性結合分析。土壤細菌占土壤微生物總量的70%~90%,是土壤最活躍的因素[17]。盡管已有土壤細菌群落結構與化學性質關系及驅動因素的研究,但對于Isohumisols典型土系的研究卻尚未報道,特別是對發生層內部土壤微生物群落結構與理化性質間的關聯性了解極為有限。本研究采用高通量測序技術,研究CST體系下的均腐土不同亞綱典型土系腐殖質層(Ah層)化學性質及土壤細菌群落結構的變化,分析細菌群落結構與環境因子的關系,進一步闡明均腐土中驅動Ah層細菌群落結構的主要環境因子,為均腐土的科學保護利用提供理論基礎和 依據。
1 材料與方法
1.1 樣品采集
于2018年10月確定8個土系的樣點位置[主要分布在46.619 57°N~48.967 35oN及124.608 10oE~ 131.860 50oE(表1)],并采集土樣。土壤樣品采集時按1 m寬、2 m長、2 m深挖掘土壤剖面,確定土系及發生層后,按發生層厚度采集土壤,本研究只針對8個典型土系的Ah層土樣進行分析,每個樣點3次重復(n=24),樣點具體概況見表1所示。將采集的土樣去除植物根系和雜草,混勻,分裝兩份,一份風干研磨后過2 mm篩,用于土壤化學性質測定,一份直接過2 mm篩,放-80℃冰箱保存,用于土壤細菌群落多樣性的 測定。

表1 均腐土8個典型土系基本特征
1.2 土壤化學性質的測定
土壤化學性質的測定采用常規方法[18]:有機碳(SOC)采用重鉻酸鉀容量法測定;全氮(TN)采用凱式定氮法測定;全磷(TP)采用HF—硝酸混合消煮,鉬銻抗比色法測定;全鉀(TK)采用HF—硝酸混合消煮,AAS法測定;pH采用電位法測定(水土比2.5∶1);陽離子交換量(CEC)采用乙酸銨—EDTA交換法測定;交換性鉀(EXK)、交換性鈉(EX-Na)、交換性鈣(EX-Ga)、交換性鎂(EX-Mg)采用乙酸銨交換-AAS法測定。
1.3 DNA提取、PCR擴增和Illumina HiSeq測序
使用mobio土壤微生物DNA強力提取試劑盒(PowerSoil? DNA Isolation Kit)提取土壤微生物DNA,用NanoDrop 2000分光光度計檢測DNA 濃度。
根據文獻[19]的方法對細菌16S rRNA基因(V3+V4)區 域 的PCR進 行 擴 增。338F(5’- ACTCCTACGGGAGGCAGCA-3’)為 正 向 引 物,806R(5’-GGACTACHVGGGTWTCTAAT-3’)為反向引物。目標區域PCR擴增反應條件為95℃預變性5 min,15個循環(95℃,1 min,50℃,1 min,72℃,1 min),72℃,7 min,然后將樣品與磁珠按照1∶1.5混勻后進行磁珠篩選片段,純化完后進行SolexaPCR,反應條件為98℃,30 s,10個循環(98℃,10 s,65℃,30 s,72℃,30 s),72℃,5 min,然后進行磁珠純化,將純化后的產物進行Nanodrop2000定量后,按照質量比1∶1進行混樣,通過1.8%的瓊脂糖凝膠,120V,40 min電泳后,切目的片段,并回收。每個樣品做3次重復,送至北京百邁克生物科技有限公司,利用Illumina Hiseq 2500平臺進行測序。
1.4 測序數據處理
對原始數據進行拼接(FLASH[20],version 1.2.11),將拼接得到的序列進行質量過濾(Trimmomatic[21],version 0.33),并去除嵌合體(UCHIME[22], version 8.1),得到高質量的Tags序列。
在相似性97%的水平上對序列進行聚類(USEARCH[23],version 10.0),以測序所有序列數的 0.005%作為閾值過濾 OTU[24]。采用RDP Classifier(version 2.2)基于Sliva(細菌,Release 132)分類學數據庫對OTU進行分類學注釋,置信度閾值為0.8[25]。
1.5 數據分析
Alpha多樣性分析,包括群落豐富度指數(Chao1和ACE)、群落多樣性指數(香農Shannon和辛普森Simpson),使用Mothur(version 1.30)軟件,對樣品Alpha多樣性指數進行評估[26]。
數據采用Excel 2016和SPSS 21.0、SigmaPlot 12.5進行整理和分析,數據分析使用單因素方差分析,多重比較運用Duncan檢驗(P<0.05,n=24)。具體制圖運用北京百邁克生物科技有限公司的BMKCloud云平臺分析。
2 結果與分析
2.1 均腐土典型土系化學性質分析
均腐土不同亞綱8個典型土系pH值為5.85~ 8.25,由低到高依次為裴德系、衛星農場系、大西江系、富牧西系、新發北系、保國系、明水系、春雷南系,其中干潤均腐土高于濕潤均腐土。土壤SOC總體差異較大,大西江系的SOC含量最高,達46.24 g/kg,而富牧西系含量最低,僅12.11 g/kg。8種土系中TK、EX-Ga均為干潤均腐土高于濕潤均腐土。均腐土的土壤化學性質指標普遍較高,其中,大西江系土壤10種化學性質指標,除TP、CEC、EX-Na外均顯著(P<0.05)高于其他7種土系(表2)。
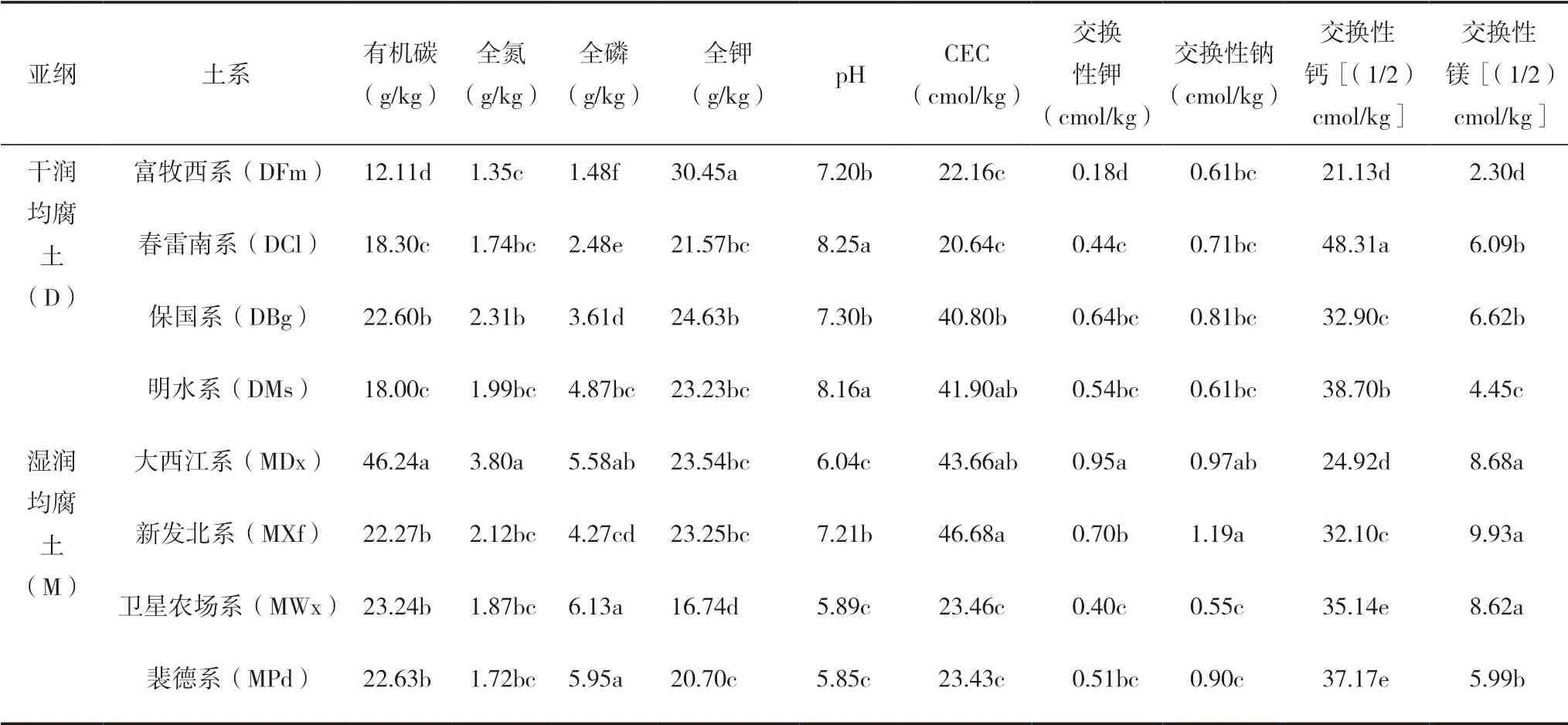
表2 均腐土土壤化學性質
2.2 均腐土典型土系細菌α-多樣性
表3顯示,均腐土的覆蓋率均為99%以上,說明該樣品測序已趨于飽和,表明Hiseq測序結果中含蓋了大多數微生物種群,可以真實反映均腐土微生物的群落結構組成。均腐土24個樣品8個土系測序共獲得1 674 634條Clean tags,各樣品的Clean tags在65 242~73 156之間(表3),有效序列在63 749~72 842之間。8個土系中有效序列的數量差異較小。干潤均腐土中春雷南系檢測出的OTU最少,保國系的OTU較高。春雷南系Clean tags檢測出的數量最高,但辛普森指數、Ace 指數、Chao1 指數均較小,表明春雷南系細菌豐度和多樣性比其他7種土系低。均腐土各土系的Ace 指數、Chao1 指數較為接近,表明均腐土8個土系的菌群豐度趨于相似。除春雷南系外,香農指數均表現為干潤均腐土>濕潤均腐土。辛普森指數表現為富牧西系與裴德系相同,春雷南系與明水系相同,保國系與大西江系、新發北系、衛星農場系相同,說明其群落多樣性相似。總體來看,在本研究中,均腐土8個土系中,干潤均腐土的細菌群落多樣性高于濕潤均腐土 (表4)。

表3 均腐土的Hiseq測序結果和多樣性指數

表4 均腐土不同亞綱的細菌多樣性指數
2.3 均腐土典型土系細菌類群
圖1為均腐土不同亞綱8個土系細菌OTU的數量和組成。在OTU水平上,8個土系共檢出 2 204個OTU,其中,干潤均腐土共有OTU為528個,占總OTU的23.96%,特有OTU分別為富牧西系13、春雷南系11、保國系43、明水系22個,分別占干潤均腐土共有OTU的2.5%、2.1%、8.1%、4.2%(圖1a);濕潤均腐土共有OTU為372個,占總OTU的16.88%,特有OTU分別為大西江系40、新發北系47、衛星農場系17、裴德系6個,分別占濕潤均腐土共有OTU的10.75%、12.63%、4.5%、1.6%(圖1b);將24個樣品的3個重復合并后發現,均腐土共有OTU為628個,占總OTU數量的28.5%。8個土系僅富牧西系、春雷南系、明水系、新發北系、衛星農場系檢出特有的OTU,分別為3、1、1、21、1個,其中新發北系特有OTU數量最多,為21個,保國系、大西江系、裴德系未檢出特有OTU(圖1c)。
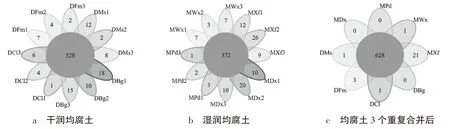
圖1 細菌OTU分布水平上的Venn圖
2.4 均腐土典型土系細菌群落組成
均腐土不同亞綱8個土系共檢測到25個細菌門,不同土系間細菌群落的差異較大。在門水平下,變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria)為富牧西系、春雷南系、保國系、大西江系、新發北系、衛星農場系、裴德系7種土系中均擁有較高的相對豐度,其總相對豐度超過了50%(圖2),這3種菌群為7種均腐土系的優勢菌群,但在組成比例上有一定的差異。與其他7種土系相比,明水系細菌群落中綠灣菌門(Chloroflexi)相對豐度達9.67%,與變形菌門、放線菌門為該土系的主要優勢菌群;而酸桿菌門在明水系中的群落相對豐度僅為5.48%,在其他7類均腐土系中均超過8%,最高達28.57%。變形菌門在春雷南系中的相對豐度最高,達45.78%。放線菌門在明水系中的相對豐度達24.58%,而在大西江系中只有12.48%。總體來看,均腐土8個土系細菌門主要優勢菌群僅變形菌門表現為干潤均腐土高于濕潤均腐土。

圖2 均腐土細菌群落在門水平上的組成和相對豐度
對于屬水平下細菌的分類組成和豐度分析能更好地為細菌門類提供完整信息,在屬水平下(圖3),共檢測到497個不同細菌屬,其中檢出豐度前10的細菌屬中,除未知菌屬外主要存在5種主要優勢菌屬:鞘氨醇單胞菌屬(Sphingomonas)、RB41、節桿菌屬(Arthrobacter)、芽單胞菌屬(Gemmatimonas)、念 珠 菌 屬(Candidatus_Udaeo-bacter)。均腐土8個土系中,富牧西系鞘氨醇單胞菌屬、RB41的相對豐度均大于5%,但其他細菌屬均較低。春雷南系鞘氨醇單胞菌屬的相對豐度達到12.25%。保國系鞘氨醇單胞菌屬、RB41的相對豐度分別達到11.13%和6.64%。明水系鞘氨醇單胞菌屬達10.44%。大西江系鞘氨醇單胞菌屬、節桿菌屬分別達到5.77%和7.96%。新發北系各細菌屬均較低。衛星農場系鞘氨醇單胞菌屬的豐度達7.64%,裴德系達14.27%。對比土系間細菌屬水平相對豐度關系發現,上述優勢菌群中各土系間均呈現顯著差異;總體來看,均腐土8個土系鞘氨醇單胞菌屬、RB41均表現為干潤均腐土高于濕潤均腐土。

圖3 均腐土細菌群落在屬水平上的組成和相對豐度
將24個樣品的3個重復合并后,基于bray- curtis距離均腐土不同亞綱8個土系細菌群落結構變化的樣品熱圖分析如圖4所示。均腐土不同亞綱分別聚為一類,其中干潤均腐土聚為一類,濕潤均腐土聚為一類。說明不同亞綱細菌群落結構差異顯著。

圖4 均腐土細菌樣品豐度熱圖
2.5 均腐土細菌群落與環境因子的關系
均腐土的細菌群落不僅受生物性質的影響,對于土壤環境因子的影響也至關重要。利用 Mental test分 析 發 現,pH、EX-Ca、EX-Mg、TP、CEC、SOC與均腐土細菌群落結構組成顯著相關(P<0.05),因此,為了進一步了解驅動均腐土細菌群落發生變化的主要影響因素,利用RDA分析發現,土壤pH值為細菌群落結構的主要影響因子,其中,EX-Ca、pH、CEC為驅動干潤均腐土細菌群落結構發生變化的主要影響因素,EX-Mg、SOC、TP為驅動濕潤均腐土細菌群落發生變化的主要影響因素。

表5 均腐土環境因子與細菌群落 組成間關系的Mental test結果

圖5 均腐土樣品細菌群落與化學性質的RDA分析
3 討論
道庫恰耶夫認為,土壤的成土過程總是由母質、氣候、時間、生物等因素相互作用而形成的,其中生物因素在土壤的成土過程中起著至關重要的作用。而威廉斯卻認為,土壤的五大成土因素中,土壤化學性質為主要影響因素[27]。本研究中,均腐土不同亞綱的化學性質有明顯差異,這可能與均腐土不同的母質與成土條件所處的環境有關,具體的研究還需深入進行。不同的土壤類型也會對土壤微生物造成影響。唐杰等[28]對不同若爾蓋濕地4種不同土壤類型細菌多樣性的研究表明,微生物數量總體上隨著土壤類型的變化而變化。張崇邦 等[29]對天臺山8種土壤類型的微生物區系研究也表明,微生物數量、細菌群落的組成和多樣性與土壤類型密切相關。Xu等[30]對不同土壤類型大豆根際細菌群落影響研究表明,大豆根際土壤細菌群落的多樣性由土壤類型決定。本研究中,均腐土不同亞綱8個土系的α-多樣性不同,細菌類群多樣性均表現為干潤均腐土高于濕潤均腐土,表明不同的土壤類型能夠影響土壤微生物群落。
本研究對均腐土不同亞綱8個土系的細菌群落組成在門、屬和OTU水平進行了分析,在8個土系中均存在較為一致的優勢菌群。而細菌菌群豐度相對較低的在均腐土中所占的比例和種類均表現出一定的差異。通過對門水平細菌群落組成研究發現,在不同亞綱8個土系的群落組成中豐度較高的有變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria)、綠灣菌門(Chloroflexi),這4類細菌為大多數均腐土綱中的優勢菌群[31-32]。劉紅梅等[32]研究發現變形菌門、放線菌門、酸桿菌門、綠灣菌門大量出現在暗栗鈣土(在中國土壤系統分類中也屬于均腐土綱)中,這也與本研究結果類似,說明類似的均腐土生境中細菌群落的分布特征也具有一定的相似性,同時表明此類細菌群落在均腐土土壤生境中至關重要,對保持土壤生態系統的穩定性有重要作用。此外,通過屬水平的群落組成研究發現,在不同亞綱8個土系的群落組成中具有降解土壤有毒物質的鞘氨醇單胞菌屬、RB41、節桿菌屬,具有土壤污染物降解功能的芽單胞菌屬、念珠菌屬相對豐度較高,說明均腐土中存在一些對土壤修復有特殊功能的菌屬,但這尚需進一步的研究。另外,對不同亞綱樣品熱圖分析發現,干潤均腐土和濕潤均腐土在聚類圖譜中被分別單獨聚為一類,表明即使是同一土綱下的土壤類型,不同亞綱細菌群落的組成也發生了明顯 分異。
近年來,關于土壤化學性質在細菌群落結構形成中的驅動作用得到了充分論證[33-34]。如張小青[35]對荒漠土壤細菌群落研究表明,pH、TN和TC是影響細菌群落結構的主導因子,侯建偉等[36]對貴州黃壤的細菌群落研究表明,土壤全氮、C/N、pH、有效磷和CEC對細菌群落結構變化貢獻較大,且TN與pH為主導環境因子,Shen等[37]和Zhang等[38]對不同海拔梯度土壤細菌群落研究表明,土壤pH為決定土壤細菌群落結構的主要影響因素。本研究發現,pH、EX-Ca、EX-Mg、TP、CEC、SOC是均腐土細菌群落結構發生變化的主要影響因子,且pH值為影響均腐土細菌群落結構的主導因子,這與前人的研究結果相同[38]。其中,pH、EX-Ca、CEC為驅動干潤均腐土細菌群落結構發生變化的主要影響因素,EX-Mg、SOC、TP為驅動濕潤均腐土細菌群落發生變化的主要影響因素,說明不同土壤類型土壤化學性質的變化是引起土壤細菌群落結構組成發生分異的主要驅動 因素。
4 結論
本研究的結果表明,干潤均腐土細菌多樣性均高于濕潤均腐土;同時,通過對均腐土不同亞綱8個土系細菌群落組成的分析發現,在門水平上變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria)為富牧西系、春雷南系、保國系、大西江系、新發北系、衛星農場系、裴德系7種均腐土系的優勢菌群。明水系以綠灣菌門、變形菌門、放線菌門為主要優勢菌群。屬水平上,均腐土各土系的群落差異較大。均腐土不同亞綱8個土系土壤細菌群落主要分為兩類,干潤均腐土為一類,濕潤均腐土為一類;對細菌群落與環境因子相關分析發現,pH、EX-Ca、EX-Mg、TP、CEC、SOC是均腐土細菌群落結構發生變化的主要影響因子;此外,RDA分析發現,pH值為影響均腐土細菌群落結構的主導因子。而pH、EX-Ca、CEC為驅動干潤均腐土細菌群落結構發生變化的主要影響因素,EX-Mg、SOC、TP為驅動濕潤均腐土細菌群落發生變化的主要影響因素。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
