生物-化學法合成維生素D的研究進展
2021-01-20 10:34:02曲麗莎于文文呂雪芹李江華堵國成劉龍
食品與發酵工業 2021年1期
曲麗莎,于文文,呂雪芹*,李江華,堵國成,劉龍*
1(江南大學 未來食品中心,江蘇 無錫,214122) 2(糖化學與生物技術教育部重點實驗室(江南大學),江蘇 無錫,214122)
維生素D(Vitamin D, VD),被稱為“陽光維生素”,是維持機體健康所必需的一種脂溶性維生素。麥角鈣化醇(ergocalciferol, VD2)和膽鈣化醇(Cholecalciferol, VD3)是VD系列化合物中主要的兩種存在形式[1]。人體皮膚組織內含有大量的7-脫氫膽固醇(7-dehydrocholesterol, 7-DHC),經紫外光照射后會轉變為VD3,該合成途徑也是人體獲得VD最主要的來源。與VD3可在體內自主代謝合成不同,VD2在人體內無法代謝合成,必須依靠外界的飲食攝取。
VD本身沒有生理活性,只有經過羥基化修飾后轉變為活性形式才能在人體內發揮生理作用。如圖1所示,飲食攝取或人體自身合成的VD3由結合蛋白(vitamin D-binding protein, DBP)從血液轉運至肝臟,在肝臟中經細胞色素P450酶羥基化修飾形成25-羥基維生素D3(25-hydroxyvitamin D3, 25(OH)D3),25(OH)D3是VD在人體內的主要循環方式,其濃度是衡量體內VD狀況的標準之一。之后25(OH)D3將由DBP再次運輸至腎臟并修飾轉變為1,25-二羥基維生素D3(calcitriol,1,25(OH)2D3)、24,25-二羥基維生素D3(24,25-hydroxyvitamin D3,24,25(OH)2D3)等活性形式[2]。
VD是骨骼正常生長發育所需的重要營養物質,人體缺乏VD時會導致佝僂病、骨質軟化等疾病[3]。VD不僅能夠促進鈣磷吸收[4],還能作用于大腦神經,預防精神分裂癥、自閉、認知障礙和神經退化等疾病[5-8],同時適當補充VD可有效治療和預防糖尿病[9-10]。此外越來越多的實驗證明,VD在治療免疫性系統疾病、癌癥以及心血管疾病等方面也具有良好的生理作用[11-13]。
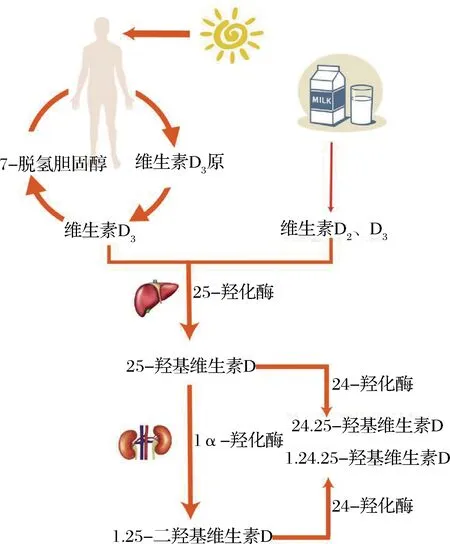
圖1 VD活性形式轉變示意圖Fig.1 Schematic diagram of VD active form transformation
隨著VD的各種生理活性不斷被發現,VD的全球市場年需求量也在日益增長。目前工業化生產VD的主要方法為半化學合成法,即以膽固醇、麥角固醇、豆甾醇、巖藻甾醇以及孕烯醇酮等為底物,經過簡單的化學反應生成[14]。反應底物是半化學合成的重要因素之一,如何大量廉價地獲取天然甾醇也是目前工業化生產VD過程中面臨的挑戰之一。麥角固醇、7-DHC分別是半化學合成VD2及VD3的理想前體,但這些天然甾醇在宿主中的單體含量較少,已漸漸無法滿足當下工業化合成VD的生產需求。
合成生物學是以系統生物學和代謝工程學的研究為基礎,使用工程化設計思路,將已知或未知的天然生物模塊進行構建串聯,從而建立全新的人工生物系統。隨著合成生物學的迅猛發展,越來越多的代謝機理途徑得到闡明,這使得人們可以借助合成生物學在微生物細胞內對宿主固有途徑進行調控,協調外源基因表達模塊與內源代謝路徑模塊,平衡各模塊之間的代謝流通量,從而在工業規模上實現其高效合成。這種新技術不僅可以滿足市場的增長需求,而且綠色環保,是解決資源可持續化問題、提高目標代謝物產量的最佳途徑之一。
利用合成生物學技術在微生物體內重構代謝途徑是提高產物產量的有效手段。由于提高VD前體甾醇的合成是增加VD得率的關鍵步驟,因此本篇文章著重介紹當前VD前體合成的新興技術與手段,主要包括動態調控、亞細胞工程、無細胞系統、定向進化等。
1 VD前體的生物合成進展
1.1 VD前體生物合成途徑
如圖2所示,甾醇在細胞內的生物合成途徑可以分為2個部分,即法尼基二焦磷酸(farnesyl diphosphate,FPP)合成途徑與后角鯊烯合成途徑[15]。
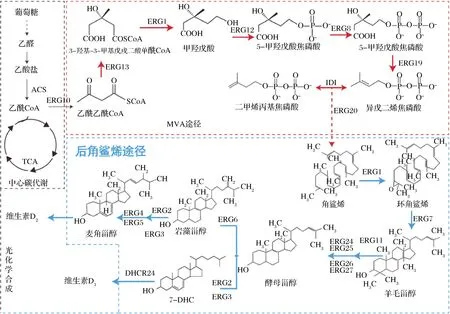
ERG10:乙酰乙酰輔酶A 硫解酶(acetyl-CoA C-acetyltransferase);ERG13:3-羥基-3-甲基戊二酰-CoA合酶(3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) synthase);HMG1:3-羥基-3-甲基戊二酰輔酶A還原酶 (3-hydroxy-3-methylglutaryl coenzyme A reductase);ERG12:甲羥戊酸激酶(mevalonate kinase);ERG8:磷酸甲羥戊酸激酶(phosphomevalonate kinase);ERG19:甲羥戊酸焦磷酸脫羧酶(mevalonate pyrophosphate decarboxylase);IDI1:異戊烯基焦磷酸異構酶(isopentenyl diphosphate isomerase);ERG20:法尼基焦磷酸合酶(farnesyl pyrophosphate synthetase);ERG1:鯊烯環氧酶(squalene epoxidase);ERG7:羊毛甾醇合酶(lanosterol synthase);ERG11:甾醇14α-去甲基化酶(lanosterol 14-alpha-demethylase);ERG24:C-14甾醇還原酶(C-14 sterol reductase);ERG25:C-4固醇甲基氧化酶(C-4 methyl sterol oxidase);ERG26:C-3甾醇脫氫酶(C-3 sterol dehydrogenase);ERG27:C3-酮基還原酶(3-keto sterol reductase);ERG2:C-8 甾醇異構酶(C-8 sterol isomerase);ERG3:甾醇-C5-去飽和酶(sterol C5-desaturase); ERG4:固醇C-24(28)還原酶(C-24(28) sterol reductase);ERG5:固醇C-22-去飽和酶(C-22 sterol desaturase);ERG6:甾醇C-24 甲基轉移酶 (Delta(24)-sterol C-methyltransferase)圖2 甾醇代謝合成途徑Fig.2 Sterol metabolic pathway
原核細胞內的FPP主要通過2-C-甲基-D-赤藻糖醇-4-磷酸(2-methyl-D-erythritol-4-phosphate, MEP)途徑代謝合成,而真核生物則是通過甲羥戊酸(mevalonate,MVA)途徑合成。真核生物中含有線粒體、內質網等細胞器,有助于關鍵酶(細胞色素P450酶、異源蛋白酶等)的表達,因此選擇真核生物如釀酒酵母(Saccharomycescerevisiae)等為底盤細胞進行甾醇產量的優化調控更為高效[16]。MVA途徑以乙酰輔酶A(acetyl coenzyme A,acetyl-CoA)為催化底物,經多步酶作用合成FPP[17]。MVA途徑是麥角固醇與7-DHC合成的共同代謝途徑,調控MVA代謝途徑,提高FPP代謝通量是最終實現甾醇的高效合成的首要方法。HMGR (3-羥基-3-甲基戊二酰輔酶A還原酶,encoding the catalytic domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase)和IDI1都是MVA途徑代謝的主要限速酶,增加HMGR與IDI1的表達量能夠顯著提高FPP的產量[18]。KEASLING等在S.cerevisiae中過表達截短的HMGR(tHMGR),減少了HMG-CoA的積累,進而提高了甲羥戊酸的產量[19]。JIANG等在S.cerevisiae中過表達tHMGR和IDI1,異戊二烯焦磷酸(isopentenyl diphosphate,IPP)和二甲烯丙基焦磷酸(dimethylallyl pyrophosphate,DMAPP)的合成產量明顯提高[20]。本課題組前期在枯草芽孢桿菌中通過異源表達kdpG基因,引入外源ED途徑,同時替換組成型強啟動子的方式強化MEP途徑中的關鍵酶基因dxs、dxr、fni的表達,增強維生素K2前體異戊二烯焦磷酸IPP的含量,最終使維生素K2的產量提高到32 mg/L[21]。
FPP在角鯊烯合成酶的催化下合成角鯊烯,角鯊烯在酶催化作用下代謝合成麥角固醇。麥角固醇是細胞膜的重要組成成分,決定著結構膜的流動性和滲透性。強化ERG1、ERG4、ERG11等基因的表達最終都會提高麥角固醇的發酵產量[22]。POLAKOWSKI等[23]提高細胞的酯化能力,進而增加了酵母胞內的甾醇濃度。Are2基因編碼甾醇-酰基轉移酶,HE等在S.cerevisiae內同時強化表達了ERG4和Are2p基因,麥角固醇的產量最終達到1 707 mg/L[24]。
麥角固醇的合成途徑天然存在于S.cerevisiae內[25],而7-DHC的合成途徑需引入外源基因3β-羥基甾醇-D24還原酶(24-dehydrocholesterol reductase, DHCR24)[26]。麥角固醇代謝途徑是合成7-DHC的最大競爭途徑,抑制或敲除ERG5、ERG6基因可以阻斷內源性麥角固醇合成途徑[27],促進酵母甾醇的積累,從而有利于7-DHC的高效合成。GUO等在家雞中分離鑒定出Gg_DHCR24蛋白,異源表達后7-DHC的產量達到64.1 mg/L,之后組合工程技術優化代謝途徑7-DHC的產量達到了1.07 g/L[28]。
1.2 代謝工程調控甾醇合成途徑
1.2.1 “推-拉-抑制”技術
Acetyl-CoA是碳代謝的關鍵中間體,其供給水平是決定MVA途徑代謝通量的首要因素。增加acetyl-CoA在MVA途徑中的供給效率,是近年來代謝工程的研究熱點。胞質內的acetyl-CoA是由丙酮酸脫羧化酶(pyruvate decarboxylase,PDC)、乙醛脫氫酶(acetaldehyde dehydrogenase, ALD)、乙醇脫氫酶(alcohol dehydrogenase,ADH)和acetyl-CoA合成酶(Acetyl-CoA synthase, ACS)等多種酶共同催化作用[29]。乙酰CoA推拉抑制策略是通過代謝工程改造天然MVA途徑和乙酰CoA途徑的常用手段。該策略包括將代謝通量“推”向乙酰CoA的有效產生,即過表達ACS、乙酰乙酰輔酶A硫解酶(acetoacetyl-CoA thiolase, AACT)或者重構非天然代謝途徑[30],使得碳代謝通量更多地流向乙酰CoA合成途徑;“拉”動乙酰CoA的有效消耗,即過表達tHMGR、IDI1以及異戊二烯合酶(isoprene synthase, ispS)[31-32]。此外,抑制丙酮酸脫氫酶旁路(pyruvate dehydrogenase,PDH)以及乙醇降解途徑同樣可以顯著提高胞質內acetyl-CoA含量[33]。LIAN等[34]在S.cerevisiae中過表達ADH2、ALD6、ACS等基因,使得碳代謝通量最大化地流向acetyl-CoA合成途徑,并在此基礎上過表達ACL基因,催化檸檬酸生成acetyl-CoA,最終胞內acetyl-CoA的濃度提高了3倍。SU等[35]基于模塊化途徑工程技術在S.cerevisiae內調控優化acetyl-CoA 的供給水平,最終7-DHC的產量增加了85.44%。
1.2.2 動態調控策略技術
傳統的靜態調控雖然可以提高目標代謝物的產量,但蛋白表達量的急劇增加會導致部分蛋白無法正確表達其功能,且會引起代謝失衡、有毒中間代謝物積累及細胞生長受損等問題。動態調控是近年來新興的一種代謝工程改造策略。相比于靜態調控,動態調控能夠響應細胞內外環境的變化,可以在提高產物合成能力的同時動態協調相關代謝網絡的流量分布,從而避免靜態調控帶來的問題,因此其在近年來受到了人們的廣泛關注。
生物傳感器是構建動態調控系統的關鍵。轉錄因子是最具代表性的生物傳感器之一,轉錄因子是能夠結合DNA上游特異核苷酸序列并且調節轉錄水平的一類蛋白質。轉錄因子由DNA結合結構域、鋅指結構域等多個功能域構成,每個區域在轉錄調控中都發揮著關鍵作用。細胞內的基因轉錄受到大量轉錄因子的調控,通過調控轉錄因子對代謝途徑優化是動態調控策略的經典思路。轉錄因子Upc2包含一個N末端鋅指結構域和一個C末端配體結合結構域,該結構域可以直接結合麥角固醇以響應胞內的麥角固醇濃度,進而調控麥角固醇的生物合成。當胞內的麥角固醇濃度較高時,Upc2與其結合從而停留在細胞核外;當麥角固醇濃度降低時,Upc2與麥角固醇上解離,并轉移到細胞核中以激活麥角固醇相關基因的轉錄[36]。
天然的生物傳感器由于其內源性的特點,在進行表達時會有更好的兼容性。但天然的生物傳感器在后續應用中存在著明顯的局限性,許多代謝物尚未發現與之對應的天然生物傳感器,這極大地限制了動態調控策略的普適性。基于此,研究者們通過結合蛋白質工程和生物傳感器技術,創造了一個全新可編程的化學誘導二聚體系統(chemically induced dimerization systems, CIDs)。不同于在已知結構位點的改造,CIDs在蛋白質異二聚體復合物的連接處構建小分子結合位點,并在該聚合體上連接3~4個殘基用于與目標配體進行相互作用;之后借助于計算機算法篩選優化相容的蛋白質支架。FPP是MVA代謝合成途徑中的重要關鍵體, 篩選構建FPP生物傳感器將具有良好應用性。ANUM等[37]選擇FPP作為目標配體,借助于CIDs系統對已有的配體-蛋白結合進行篩選改造,最終得到了響應FPP的生物傳感器。
群體感應系統(quorum-sensing, QS)是動態調控策略中另一經典思路,該系統能夠較好地平衡細胞生長與代謝產物之間的關系[38]。不同于傳統的人工誘導調節,QS能夠通過調控代謝產物來響應細胞濃度的變化,即細胞濃度達到一定的閥值范圍時,控制代謝途徑的開關才會響應表達[39]。圖3形象展示了QS系統在釀酒酵母細胞內的應用。THOMAS等[40]在S.cerevisiae內首先以響應芳香氨基酸的啟動子ARO9控制表達α信息素基因,使得細胞濃度與α信息素濃度相關聯;啟動子FUS1可以響應α信息素,因此外源基因的表達由啟動子FUS1控制,最終構建出適用于S.cerevisiae的QS系統。WILLIAMS等[41]基于QS系統的基礎上,結合RNA干擾(RNA interference, RNAi)技術開發了雙向動態調控系統。在釀酒酵母細胞中導入AGO1和DCR1基因,當目標代謝產物途徑響應α信息素進行高效表達時,雙鏈發夾結構的RNA(dsRNA)可以靶向任何mRNA,并對其進行降解,使目標基因沉默。因此通過RNAi干擾對競爭代謝支路進行抑制作用,進而更加高效地實現目標代謝物的合成。基于群體響應具有不依賴代謝途徑,不需額外添加誘導劑等優勢,本課題組前期在枯草芽孢桿菌中利用群體響應系統Phr60-Rap60-Spo0A動態調控脂溶性維生素K2的合成,使細胞的生長、有毒代謝物的積累與維生素K2的高效合成形成相對平衡的狀態,使維生素K2的產量相較于野生菌株產量提高40倍,在15 L生物反應器中,第3天的產量達到200 mg/L[21]。
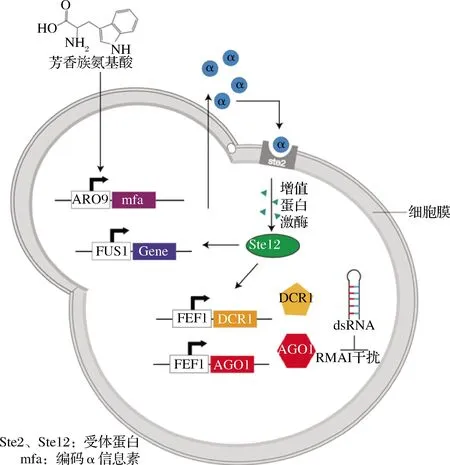
圖3 動態調控Fig.3 Dynamic regulation
1.2.3 亞細胞工程
與原核生物相比,真核生物胞內具有內質網、線粒體等細胞器。這些細胞器在微生物生長代謝過程中發揮著重要作用。亞細胞工程就是對某些特定細胞器進行理性改造,使其能夠更加有利于目的產物的代謝合成。代謝區室化可以為生物合成途徑提供合適的物理化學環境、充足的前體物質和相應的酶。正如圖4所示的乙酰CoA,其不僅可以在胞質內生成,還可以在線粒體、過氧化物酶體等多個細胞器中合成[29]。這類細胞器中并沒有天然的MVA途徑存在,因此在其中重構MVA途徑可避免其他支路途徑的競爭,提高代謝合成效率,增加FPP合成來源。線粒體基質中氧化還原電位較高,ATP含量豐富,有利于各種生化反應。此外線粒體內的A乙酰CoA濃度是胞質濃度的20~30倍,可以極大地提高MVA途徑的代謝通量[42]。DANIELLE等[43]在MVA途徑基因N端添加特定的定位氨基酸序列,成功在線粒體內重構完整的MVA途徑。LV等[32]以S.cerevisiae為出發菌株,同時調控優化胞質產乙酰CoA途徑和線粒體產乙酰CoA途徑,促進了細胞生長并且提高了發酵產率。LIU等[44]在S.cerevisiae中發現過氧化物酶體是儲存角鯊烯的動態倉庫。利用過氧酶體作為亞細胞間室進行角鯊烯合成,使角鯊烯的發酵產量(1 312.82 mg/L)相對于親本菌株提高了138倍。通過細胞質和過氧化物酶體工程的雙重調控,角鯊烯發酵產量進一步提高到1 698.02 mg/L。這表明過氧化物酶體同樣具有成為甾醇類生物合成亞細胞工廠的潛能。

圖4 代謝區室化Fig.4 Metabolic compartment
內質網(endoplasmic reticulum,ER)是蛋白質合成和折疊的場所,當蛋白表達量急劇增加時,ER無法平衡其蛋白合成與折疊修飾能力,蛋白質的合成效率降低,部分蛋白合成后無法折疊修飾,此時需擴張ER的尺寸進而來恢復ER的正常生理功能。ER的尺寸調節是通過誘導磷脂合成實現的[45],ER的擴張不僅為細胞提供了額外的空間來容納結合蛋白,而且還提高了蛋白質合成的能力。細胞色素P450是典型定位于內質網的蛋白酶,強化基因INO2的表達時,內質網尺寸會明顯擴張,該類蛋白酶的活性也會因此顯著提高。因此對于含有細胞色素P450酶的代謝途徑,內質網是理想的改造目標。
1.2.4 無細胞系統
通過微生物代謝工程做到生物制品的可持續、低成本的生產是實現工業化的必然趨勢。而當前技術存在的常見問題包括有毒中間體或產物的積累、關鍵前體在競爭途徑中的損失、最終產品的回收昂貴等[46]。為了克服這些局限性,無細胞代謝工程應運而生,其無需使用完整的細胞即可進行精確的復雜生物分子合成。通過使用體外純化的酶或粗裂解物制備的催化蛋白集合體來擴展傳統生物工程模型的范圍,以生產目標產物[47]。圖5簡單介紹了傳統代謝工程和無細脆代謝工程之間的差別。由于已知每種酶組分的濃度和活性,此系統可以精確控制反應條件和途徑通量。研究表明,通過無細胞系統可提高MVA途徑中間體和目的產物的合成。RODRIGUEZ等[48]利用無細胞代謝工程對MVA途徑進行改造,同時對酶底物和產物進行及時回收,能夠生產40 g/L的DMAPP和IPP。
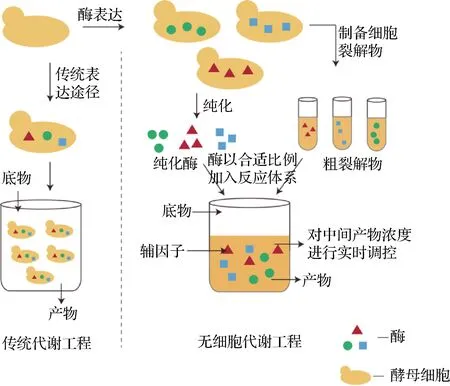
圖5 無細胞系統Fig.5 Cell-free system
從生物制造的角度來看,無細胞系統將細胞生長與代謝產物生產分開,并將所有碳通量轉移到產物上,可以實現更高的產率和回收率,并且更容易控制反應條件。但是,對于一個需要大量酶參與反應的途徑來說,利用無細胞系統成本較高,輔因子和酶的再生也是一個不可忽視的挑戰。另外,目前體外系統的成本遠超體內方法的成本,限制了規模的擴大,因此大多數無細胞系統尚未應用在工業化生產上。
1.2.5 定向進化
定向進化是改善微生物性能的有效方法之一,旨在篩選符合預期的正向突變體。定向進化首先需建立基因突變庫,之后結合與高通量篩選方法快速提升蛋白的特定性質,是目前最為常用的蛋白質設計改造策略[49]。基因突變庫的構建手段主要包含隨機突變、半理性設計和DNA改組等。易錯PCR是目前最常用的隨機突變方法之一,其通過改變PCR反應條件,降低DNA聚合酶的保真性,從而提高PCR反應的突變率,使錯誤的堿基以一定的頻率隨機地摻入到擴增的基因產物中,得到DNA突變文庫;隨后結合高通量篩選方法篩選出符合預期目標的突變基因[50]。IDI是甾醇代謝途徑中的關鍵限速酶之一,天然的IDI酶活性低,半衰期短,底物親和力弱。CHEN等[51]通過易錯PCR以及組合位點特異性飽和誘變,篩選得到正向突變體,突變體IDI的Km比親本IDI的Km降低了10%,Kcat比原IDI提高了28%。
相比于易錯PCR的體外建庫,ICE(Invivocontinuous evolution)是一種應用于真核生物體內的定向進化策略。該策略利用酵母細胞中天然存在的Ty1轉座子,擴展酵母體內的誘變系統。將需進化的目標基因克隆至可誘導的Ty1反轉錄轉座子的基因組中,之后逆轉錄酶將Ty1基因組以非高保真形式逆轉錄為cDNA,然后重新整合至基因組中。CROOK等[52]以S.cerevisiaeBY4741(Δrrm3)為出發菌株,基于ICE技術以木糖代謝途徑作為進化目標,發現全局代謝途徑進化的最適突變體并非該途徑中單個酶最佳突變體的簡單疊加。
隨機突變的局限性在于所需的突變文庫較大,并且產生的絕大多數突變體不符合預期性能或性能不佳。而半理性設計可以在人工選擇的特定位置或區域內進行隨機突變,可以大大縮小突變文庫的容量,更易于篩選。FISCHER等[53]對ERG20進行同源模建,發現第197位的賴氨酸(Lys)是關鍵活性位點,之后以代謝物香葉醇產量作為酶活的間接表征方法,對該位點進行了一系列的定點突變,最終香葉醇的發酵產量提高約10~20倍,間接證明了突變后的ERG20酶活的顯著提高。
與ICE策略類似,EvolvR系統可以實現胞內的定點突變[54]。該系統包含一個含有導向序列(gRNA)的質粒,改造后的enCas9蛋白(改造后其與非特異性DNA的親和力降低),以及與enCas9蛋白融合表達的非高保真DNA聚合酶。當gRNA引導enCas9-DNA聚合酶融合蛋白至基因組上的靶向位點時,enCas9蛋白與靶基因結合使DNA雙鏈斷裂,然后非高保真DNA聚合酶對切口進行靶向突變。EvolvR可以同時對多個靶向基因進行定點突變,且在酵母細胞中具有傳代時間短、細胞生長繁殖快速等優勢。EvolvR技術為高等真核生物體內定向突變提供了一個高效的平臺,但該技術目前仍存在一定的缺陷性,即胞內會存在游離的非高保真DNA聚合酶對整個基因組進行低概率的突變,因此距離實現完全定向的突變仍具有一定的距離。
定向進化是當前備受矚目的研究熱點領域。通過對蛋白質進行多輪突變、表達和篩選,引導蛋白質的性能朝著預期方向進化,進而大幅縮短蛋白質進化的過程[55]。但定向進化存在的最大挑戰即是如何建立適宜高效的篩選方法,目前較為普遍的篩選方法是選擇適合的生物傳感器, 并以熒光基因表征突變結果。但如前文所述,許多代謝物目前仍未有與之響應的生物傳感器,無法結合高通量篩選技術,這也成為定向進化技術提高應用普適性的難題之一。
2 光化學合成VD
VD是由關鍵前體甾醇經過光化學開環反應產生的。VD的光合作用是一個由多條異構化反應組成的復雜分支網絡。
麥角固醇轉化為VD2的過程中,影響VD2的轉化率關鍵在于光源的選擇和光轉化器構造。目前國內光源的選擇一般選用低壓和高壓汞燈兩種。在VD2的光化學反應中波長的選擇很重要。275~300 nm波長范圍內,280~284 nm的紫外光有利于生成VD2,短于280 nm的光易于生成速甾醇,而長于284 nm的光容易生成光甾醇,VD2過度照射,過度輻射還會生成毒甾醇等物質[56],因此VD2的光轉化中熒光燈的波長范圍要求極其嚴格。圖6展示了VD在光照條件下的像話轉化。研究發現,波長為283 nm的低壓汞蒸氣紫外熒光燈照射麥角固醇,轉化效果最佳,麥角固醇轉化率50%以上,VD2收率在70%以上[57]。

圖6 VD的光化學反應Fig.6 Photochemical reaction of VD
7-DHC則會被290~315 nm波長的光子異構化為VD3,這與紫外線B (UVB)的電磁波譜范圍相對應。FUSEO等提出利用連續微流系統合成VD3[58],與傳統的兩步法合成VD3不同,該方法在微反應器中可以同時進行光反應和熱反應。7-DHC在反應器中光異構化后轉化為維生素原D3,維生素原D3繼續熱異構化轉化為VD3。相比于傳統光合成法,該方法收率高(HPLC-UV:60%,分離率:32%),操作簡便且無需純化中間產物維生素原D3,具有良好的工業應用價值。
3 展望
隨著人們逐漸意識到VD對于維持機體健康的重要意義,其全球年市場需求量也因此持續增加,但關鍵前體甾醇的供給不足卻成為限制VD產能擴大的主要因素之一[59-60]。隨著合成生物學的發展,在細胞內調控重構甾醇代謝途徑,利用微生物發酵的方式制備甾醇成為解決甾醇可持續平穩供給的有效技術之一。
本文著重從推-拉-抑制、動態調控、亞細胞工程、無細胞系統、定向進化等方面闡述了代謝工程技術在甾醇代謝調控領域的研究應用。然而這些方法雖能夠顯著地提高甾醇代謝途徑的催化效率與產量,但仍有些許方面值得進一步研究與探索:
(1)如何高效實現酶的催化活性?
酶的豐度和活性共同決定了酶的催化效率,過表達基因增加酶的豐度雖然在一定程度上可以提高酶的催化效率,但當豐度達到一定閾值后,酶的催化活性卻無法繼續提高,其主要因素之一就是忽視了輔因子在代謝途徑中的關鍵作用[61]。MVA途徑代謝過程中需要NADPH、ATP等輔因子的參與[62],因此在提高酶豐度的同時需增加NADPH、ATP等輔因子的供給。調控NADH激酶、NADP+依賴型脫氫酶增強NADPH合成途徑,重構、替換消耗NADPH的支路代謝途徑,使得NADPH等輔因子最大化地流向MVA代謝途徑。輔因子的相對平衡是維持細胞內環境穩態的重要條件之一,而胞內全局的輔因子調控則需要對輔因子的合成、分布、運輸和調節機制完全了解。
(2)如何實現甾醇代謝途徑的整體定向進化?
定向進化可以改善細胞魯棒性,優化細胞性能。然而目前定向進化主要集中于 “點”的進化,即對某一特定酶的改造。但代謝途徑的催化效率并非是各種酶的簡單疊加,每個“點”在整個代謝途徑中需要的是最適的催化效率而非最高的催化效率。甾醇合成途徑包含MVA途徑、后角鯊烯途徑等,涉及到多種蛋白酶,且許多中間代謝物沒有與之特異響應的生物傳感器,無法建立合適的高通量篩選方法。ICE與CIDs策略的交叉結合將成為解決這一難題的突破口,ICE策略有望實現甾醇代謝途徑的全局性進化,而CIDs策略則可以計算、模擬、構建出非天然的生物傳感器,建立高效、簡便的高通量篩選方案。
(3)如何真正實現從頭合成VD?
目前工業化制備VD的方法是以甾醇為前體,再經過化學合成得到VD。如何在細胞內真正實現從頭合成VD將會是未來研究的熱點與難點,其關鍵在于是否能夠鑒定或人工設計出特定的酶來實現光化學合成這一步驟的功能[63]。酶工程是一種具有針對性的策略,借助蛋白質晶體結構和計算機模擬,酶工程可以提高酶的特異性和效率,甚至為特定的反應創造非天然的酶[64]。隨著結構生物學的發展,越來越多的催化酶晶體結構得到了解析,這為鑒定未知酶、構建非天然酶提供了可行性基礎。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛生(2016年3期)2016-11-12 13:23:26
