3種天牛幼蟲蛀食振動小波包特征及其識別*
2021-01-26 00:48:34劉圣煌楊江天崔建新
林業科學 2020年12期
劉圣煌 楊江天 崔建新
(1.北京交通大學機械與電子控制工程學院 北京 100044; 2.河南科技學院害蟲天敵繁育研究中心 新鄉453003)
天牛是鞘翅目(Coleoptera)葉甲總科(Chrysomeloidea)天牛科(Cerambycidae)昆蟲的總稱,全世界已知超過2萬種,我國有4 000多種(王直誠, 2014),絕大多數為各類樹木的蛀干害蟲。天牛幼蟲在樹木內部蛀食樹干,通常會引起樹木生長受阻,產量減低,樹勢削弱,壽命縮短。受害嚴重時,可能導致樹木迅速枯萎與死亡。此外,被蛀食的樹木容易引起其他害蟲及病菌的侵入,易受大風吹折。林木受蛀害后,品相必然會降低,甚至失去觀賞性和商品價值。例如栗山天牛(Massicusraddei)在我國東北地區為害櫟樹每年造成數十億元經濟損失(唐艷龍等, 2011); 云斑天牛(Batocerahorsfieldi)嚴重為害白蠟(Fraxinuschinensis)、楊樹(Populus)、柳(Salix)、榆(Ulmus)、桑(Morus)、核桃(Juglansregia)等林木,對我國城市園林綠化景觀造成巨大危害(嚴敖金等, 1997; 路紀芳等, 2016); 滅字脊虎天牛(Xylotrechusquadrips)為害經濟作物咖啡(Coffea),在印度每年造成大約2億美元的損失(付興飛等, 2020)。同時天牛幼蟲會隨著木材的流動在世界范圍內遷移,造成生物入侵,對生態環境的穩定造成威脅。天牛不同種間的生物學習性差異很大,如栗山天牛在遼寧3年1代,云斑天牛在華北地區2年1代,滅字脊虎天牛在云南1年1代或2代(肖剛柔, 1991; 唐艷龍等, 2011; 楊文波等, 2017)。天牛幼蟲存活在樹干內部,傳統的農藥毒殺防治效果不佳而且污染環境,因而采用寄生蜂進行生物防治是常用的控制方法。利用腫腿蜂防治天牛幼蟲是我國最為重要的生防措施之一,在防治栗山天牛時,釋放白蠟吉丁腫腿蜂(Sclerodermuspupariae)防治的最佳時期是每年的8—9月和次年的5—6月,栗山天牛的幼蟲齡期分別為1齡和2齡,寄生率和致死率較高,防治效果最好(唐艷龍等, 2014)。而利用川硬皮腫腿蜂(Sclerodermussichuanensis)防治云斑天牛的最佳時間是6月中旬至7月中旬,此期云斑天牛的幼蟲齡期適宜(楊樺等, 2011; 梅增霞等, 2014; 肖剛柔, 1991)。由于利用寄生蜂只能針對某一種天牛或某些類天牛的特定發育階段寄生。因此,在不破壞樹干的前提下,確定天牛幼蟲的種類,了解其生物學習性,可以準確地指導適時防治,是開展天牛類隱蔽蛀干害蟲科學防治的關鍵技術。
目前常用的蛀木害蟲檢測方法仍為目視、剖檢、敲擊等人工檢測手段(潘杰等, 2013)。此種方法費時、費力、效率低。同時由于檢測方法沒有工程化,準確性受檢測人員的專業技術水平的限制。目前常用的工程化檢測方法有X光檢測法和聲音檢測法。X光檢測法存在以下問題: 1) 設備笨重且價格昂貴,不適合野外使用; 2) 難以確定天牛的種類; 3) X射線對人體有一定的傷害。利用害蟲發出的聲音識別害蟲種類已經有成功的報道,卜宇飛等(2016)利用Adobe Audition 3.0和MATLAB完成了對7種害蟲聲音信號的采集分析處理,有效提取了害蟲聲音特征。Mankin等(2008)在消音箱內采集害蟲發出的聲音信號,進行頻譜分析,成功識別出樹干內天牛幼蟲種類。但害蟲發出的聲音信號信噪比低,且易受到環境噪音干擾,實際應用受到很大限制。本文提出了通過采集分析幼蟲蛀食樹干產生的振動信號,檢測天牛種類方法,克服了聲音檢測方法的缺點,同時具有價格低廉、檢測時對樹木傷害小的突出優點,有很好的應用前景。
1 材料與方法
1.1 材料
試驗樹木及天牛幼蟲種類見表1。試驗地點為河南省新鄉市人民公園,測試時間為2017年7月18日,采樣時氣溫約為33 ℃,當天氣溫均值28 ℃。

表1 試驗樹木及天牛幼蟲種類Tab.1 Species of trees and longicorn larvae
1.2 方法
振動傳感器采用江蘇聯能電子有限公司生產的YD-189型壓電式加速度計,靈敏度5.015 V·g-1,頻率范圍0.2~5 000 Hz,最大橫向靈敏度<5%,量程±5 g。數據采集器為基于NImyDAQ開發的便攜式測振儀,適合野外環境下使用。
信號采集選擇無風(至少沒有強風)晴朗天氣條件下進行,這是因為大風會造成林木樹干晃動,對害蟲蛀木振動信號產生干擾。天牛幼蟲蛀木振動信號采集辦法如下:
1)為了保證試驗的準確性,應提前進行調查,選擇只有一種天牛幼蟲的樹木進行試驗。挑選糞孔附近的樹干表面,削去樹皮并刮出一個φ20 mm的平面,用502膠將傳感器黏接在樹干上。
2)用便攜式檢測儀采集天牛蛀木振動信號。設定采樣頻率10 000 Hz,每段采樣時間120 s。程控放大增益為小樹(周長小于80 cm)200倍,大樹(周長大于80 cm)400倍。這樣選擇程控增益放大倍數的目的是盡量將微弱的蛀木振動信號放大到數據采集器滿量程以下,半量程以上。每種樹采集20棵的樣本。
采集數據以后,解剖樹干,如果樹干內有1只以上天牛幼蟲或其他蛀干害蟲,將這組數據剔除,即保證分析的數據都只有1只天牛幼蟲。對各種天牛幼蟲初步鑒定后,分別罩籠飼養,直至飼養出成蟲完全確定種類為止。天牛成蟲的監測常年進行,種類監測結果與室內罩籠飼養結果相互確認。
2 結果與分析
2.1 數據篩選
卜宇飛等(2017)用紅外攝像機觀測天牛幼蟲在蛀道內的活動,同時采集幼蟲發出聲音信號,找到了天牛幼蟲活動的聲音信號特征(表2)。“振是聲之源”——振動擠壓周圍的空氣,產生聲音。天牛幼蟲活動聲音信號和振動信號有同樣的特征(劉圣煌, 2019)。根據這一原理進行振動信號篩選,天牛幼蟲在蛀道內活動引發的樹干振動信號由一系列沖擊振蕩組成,如圖1所示。從表2中可以看出,外界干擾與天牛幼蟲活動引起的振動在波形和相對幅值上存在比較大的差別,可以據此進行篩選,找出天牛幼蟲蛀食振動。
圖2為光肩星天牛幼蟲的蛀食樹干振動信號,圖3為由于風引發的樹干振動,圖4未定性的樹干振動。可以看出,圖2、3、4的波形明顯不同。通過此種方法可以篩選出由天牛幼蟲活動所產生的振動信號。
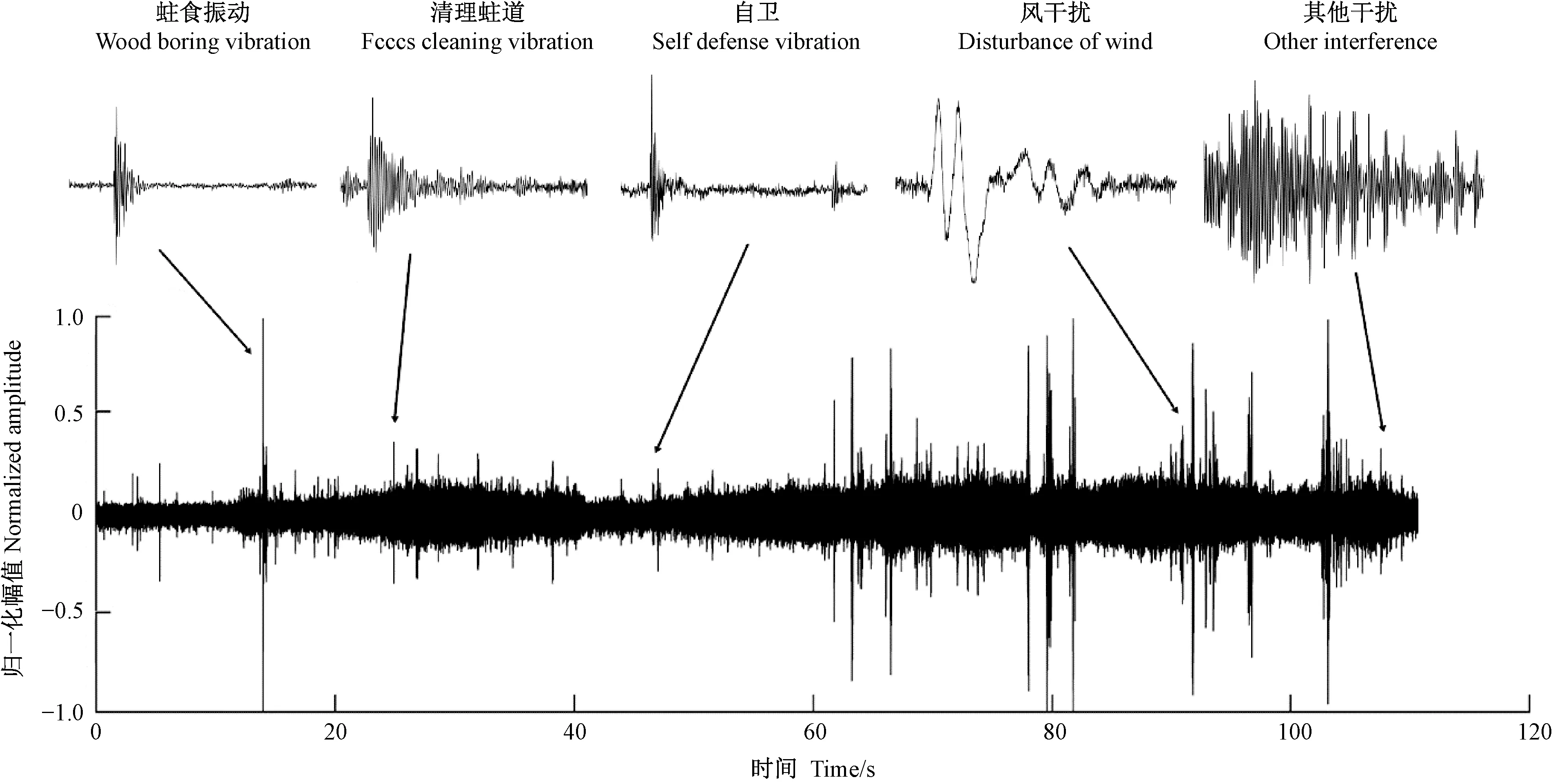
圖1 天牛幼蟲活動引起的樹干振動信號Fig.1 The vibration signal of tree trunk caused by longicorn larvae
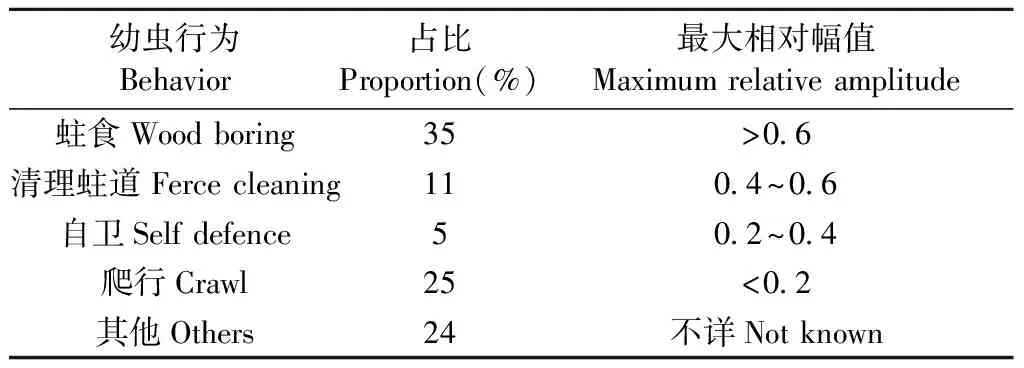
表2 天牛幼蟲行為占比及特征①Tab.2 Behavioral proportions and characteristics of longicorn larvae
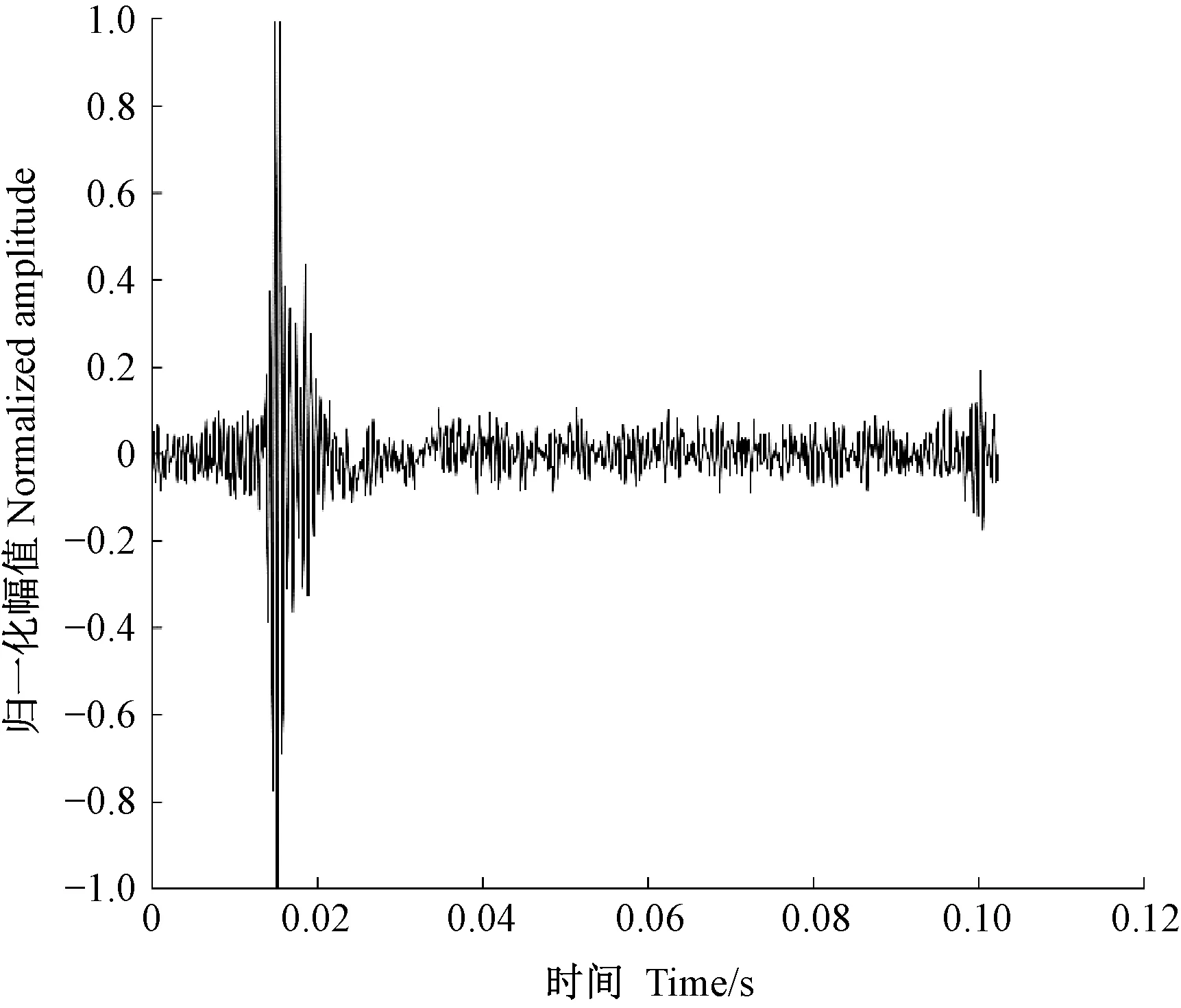
圖2 光肩星天牛蛀食二球懸鈴木樹干振動信號波形Fig.2 The vibration signal of Anoplophora glabripennis larva boring tree trunk of Platanus acerifolia
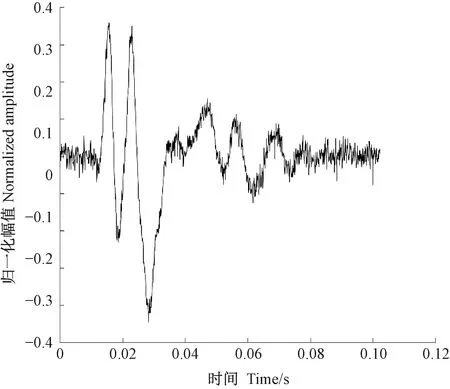
圖3 風吹引發的樹干振動信號波形Fig.3 The vibration signal caused by wind blows
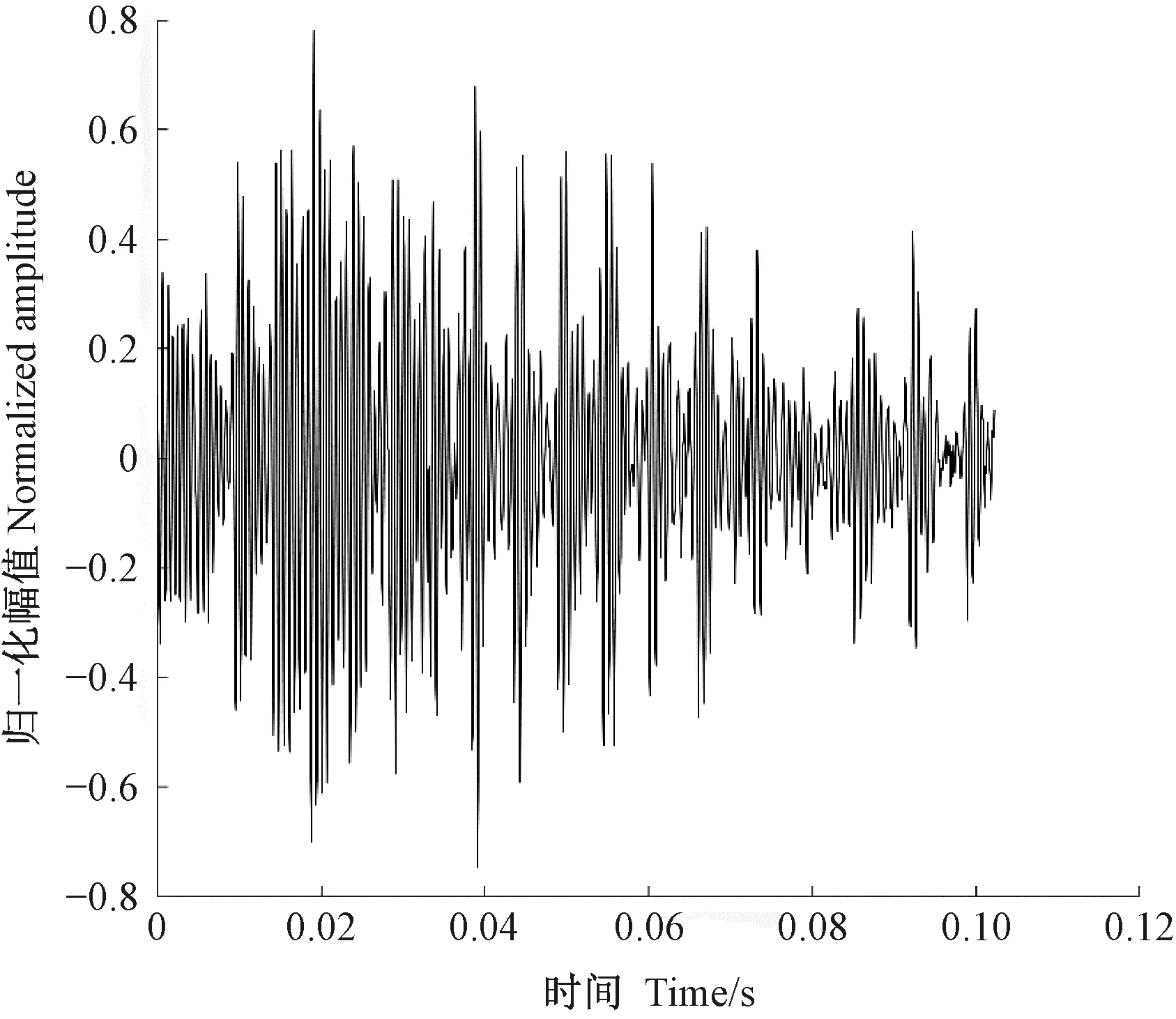
圖4 未定性樹干振動波形Fig.4 The undetermined trunk vibration waveform
天牛幼蟲在蛀道內除了蛀食行為外,還存在清理蛀道、自衛、爬行等其他的行為,這些行為都會引起樹干的振動。分析蛀食振動信號需要排除天牛幼蟲蛀食以外活動引起振動的干擾。相關研究(卜宇飛等, 2017)表明,天牛幼蟲在蛀道內不同活動引起樹干振動的相對幅值和波形存在差異。蛀食振動引起的沖擊最大,上升沿一個非常陡的脈沖,然后迅速衰減,持續時間短,最大的相對振幅可達0.8,明顯大于其他3類行為引起的振動; 天牛幼蟲在蛀道內爬行引起的振動持續時間較長且不規律,振幅有波動,相對振幅一般不超過0.2,在4類行為中最小; 天牛幼蟲清理蛀道引起的樹干振動存在一個較大的沖擊,相對振幅值可達0.5~0.6,僅次于蛀食振動; 天牛幼蟲自衛引起的振動波形間隔時間極短,一般小于50.0 ms,連續出現,振幅大于爬行引起的振動,相對振幅可達0.4。根據上述特點,篩選天牛幼蟲不同行為引起的樹干振動。圖5為天牛幼蟲蛀食樹干振動信號波形,圖6為幼蟲清理蛀道樹干振動信號波形,圖7為天牛幼蟲自衛引發的樹干振動波形。由于爬行引發的振動最大相對振幅值為0.2,未采集到振動波形。
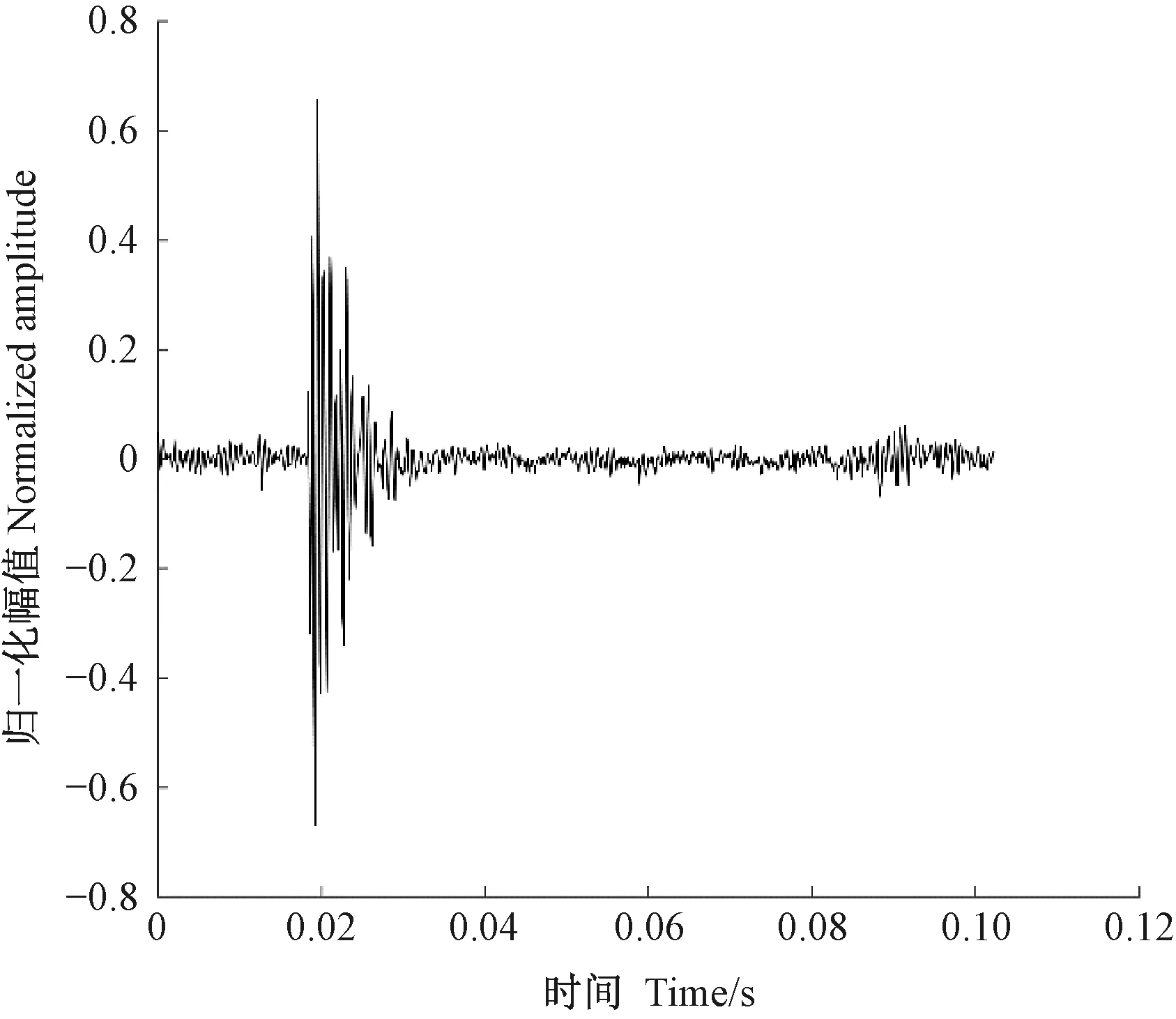
圖5 云斑天牛幼蟲蛀食白蠟樹樹干振動信號波形Fig.5 The vibration signal of Batocera horsfieldi larva boring tree trunk of Fraxinus chinensis
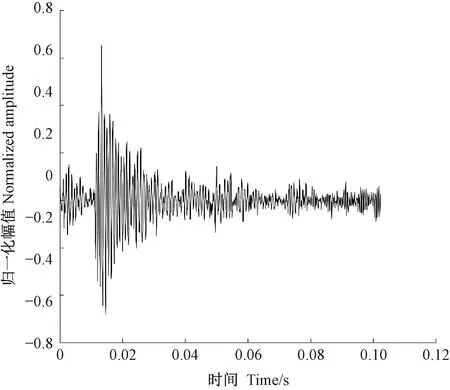
圖6 云斑天牛幼蟲清理白蠟樹樹干蛀道振動信號波形Fig.6 The vibration signal of Batocera horsfieldi larva cleaning feces in tree trunk of Fraxinus chinensis
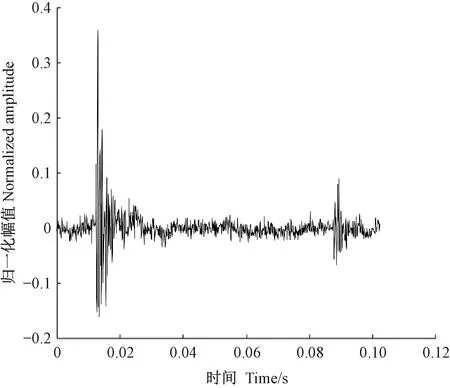
圖7 云斑天牛幼蟲在白蠟樹樹干內自衛活動的振動信號波形 Fig.7 The vibration signal of self-defense of Batocera horsfieldi larva in tree trunk of Fraxinus chinensis
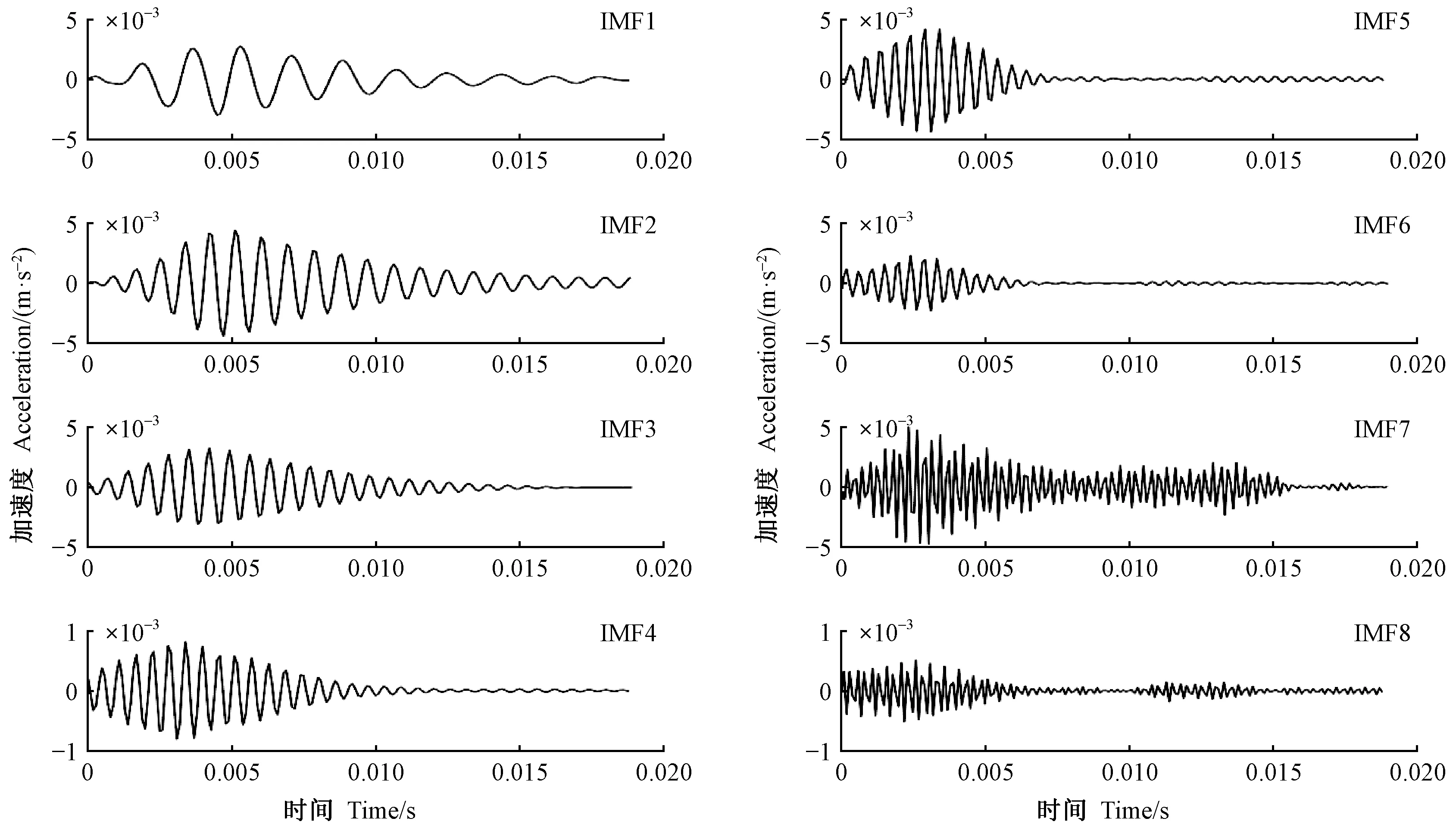
圖8 云斑天牛蛀食白蠟樹樹干振動信號變分模態分解Fig.8 The variational mode decomposition of the vibration signal of Batocera horsfieldi larva boring tree trunk of Fraxinus chinensis
2.2 振動信號降噪
使用變分模態分解(variational mode decomposition,VMD) (Dragomiretskiyetal., 2014) 對采集到的害蟲蛀木振動信號進行降噪處理。其原理是先將原始信號分解成一系列本征模態函數(intrinsic mode function,IMF),檢測各本征模態函數找出噪聲分量,濾除噪聲分量后重構原信號。使用中心判別法實驗確定變分模態分解最佳模態個數為8,懲罰因子為2 000。本文采用波形與時間熵判別法識別噪聲和有用信號。
截取天牛幼蟲蛀食振動信號前1 000個采樣點的數據進行分析,采樣頻率為10 000 Hz,因此實際振動信號的長度約為0.1 s(100 ms)。害蟲蛀食振動信號通常在短時間(30 ms)內迅速衰減,但是噪聲信號幅值不會很快衰減。根據這一性質,觀察變分模態分解得到的各本征模態函數的時域波形,如果幅值變化大就是有用信號,反之就是噪聲。
圖8為云斑天牛(為害白蠟)蛀干振動信號做變分模態分解得到8個本征模態函數。容易看出,IMF1—IMF6 波形迅速衰減,為有用信號; 而 IMF7、IMF8波形衰減很小,是噪聲。同時計算各分量的時間熵分別為2.25、2.02、1.79、1.77、1.69、1.76、2.96和3.00。熵是信號混亂程度的度量,熵值越大,說明信號越混亂無規律。據此也可以判斷IMF7和IMF8 分量為噪聲。
2.3 時域特征與頻域特征
選擇天牛幼蟲蛀木振動信號的波形和持續時間作為時域特征,信號能量隨頻率的分布作為頻域特征識別天牛種類。對VMD降噪后的振動信號進行3層小波包分解,小波基函數選用濾波器截止性能最好的dmey小波。對各節點小波包系數做傅里葉變換,計算各小波節點的頻域能量,以各節點頻域能量在總能量中的占比作為頻域特征參數。圖9為云斑天牛(為害白蠟樹)的蛀食樹干振動信號小波包分解,左側為各結點小波包系數,右側對應的功率譜。從圖中可以看出,前面4個節點信號幅值比后面4個節點信號的幅值大很多,信號能量主要集中在前4個節點,對應頻帶 0~2 500 Hz。因此本文以前4個節點的能量作為特征,判斷天牛幼蟲種類。小波包分解各個節點所對應的頻帶見表3。
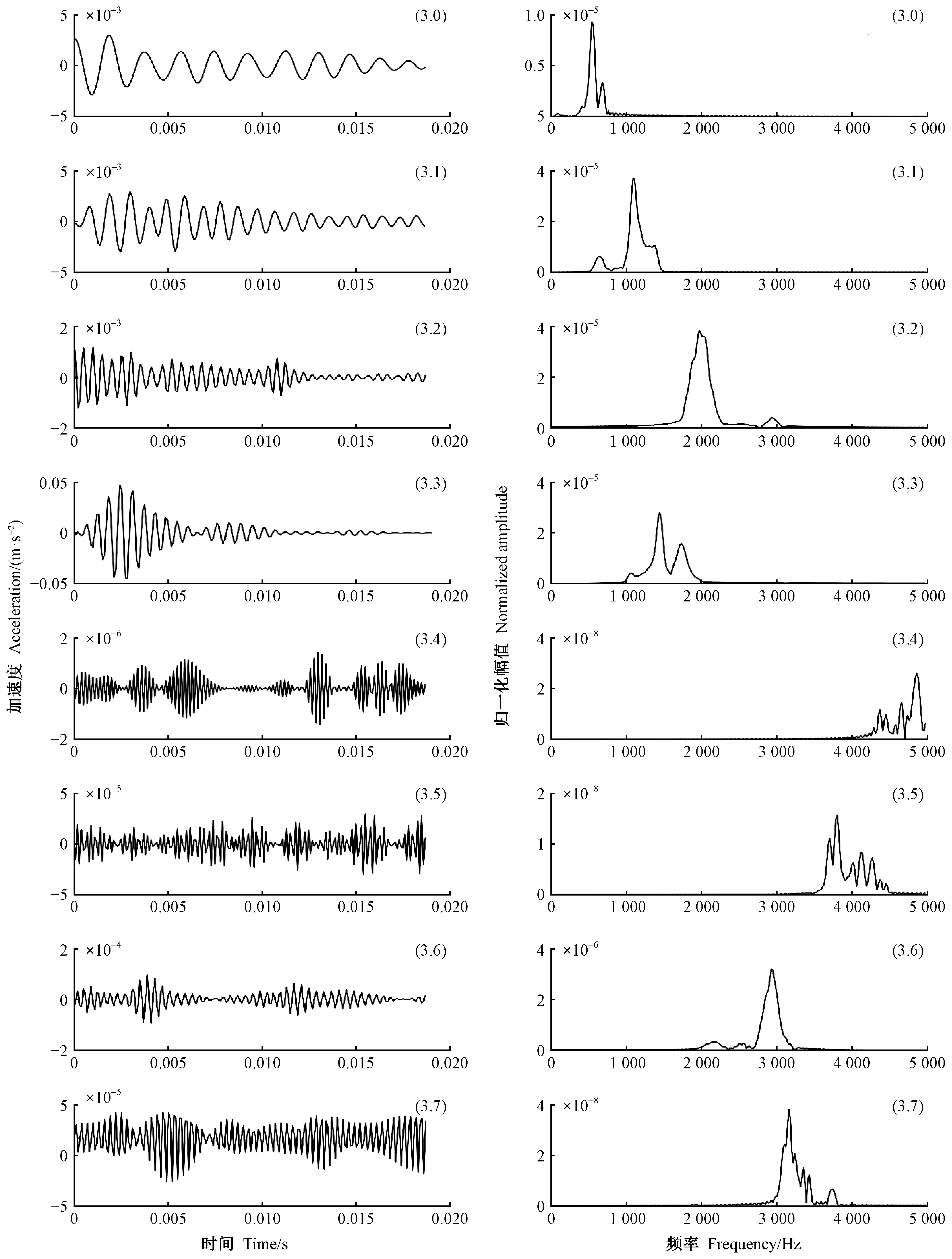
圖9 云斑天牛蛀食白蠟樹樹干振動信號小波包分解Fig.9 The wavelet packet decomposition of the vibration signal of Batocera horsfieldi larva boring tree trunk of Fraxinus chinensis
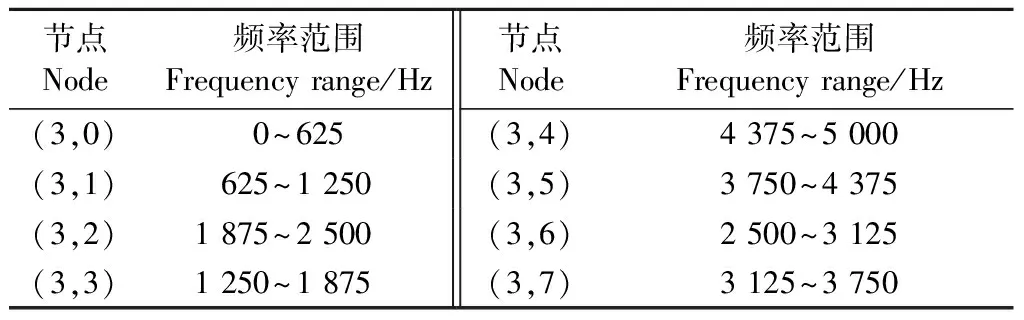
表3 蛀木振動信號小波包分解各結點對應頻帶Tab.3 Frequency range of wavelet packet nodes
2.4 3種天牛蛀食振動時域特征與頻域特征
天牛幼蟲在蛀道內活動引起的樹干振動信號由一系列頻率、幅值差別較大的沖擊信號組成,成分復雜。分析這些振動信號是檢測天牛種類的基礎。在天牛幼蟲活動行為中,蛀食樹干所占比例最大,引起的振動相對幅值也最大,因此可通過分析天牛幼蟲蛀干振動信號識別天牛種類。天牛幼蟲蛀干振動信號的上升沿是一個陡的脈沖,與天牛幼蟲蛀食樹木相對應。隨后能量被樹干結構阻尼消耗,脈沖以近似指數包絡線衰減。蛀木振動信號特征與天牛種類、齡期、個體大小,以及寄主樹木材質、水分、粗細、密度等諸多因素有關。光肩星天牛、云斑天牛與桃紅頸天牛蛀食二球懸鈴木、白蠟樹、紫葉李和西府海棠樹干的振動信號具有不同的時域與頻域特征。圖10—13分別為云斑天牛蛀食白蠟樹干、光肩星天牛蛀食二球懸鈴木樹干、桃紅頸天牛蛀食紫葉李樹干、桃紅頸天牛蛀食西府海棠樹干的振動信號波形。從圖中可以看出,天牛幼蟲蛀食樹干振動的4種信號都是衰減震蕩,但是各自波形特征、頻域特征、能量分布不同。這些差別是識別不同種天牛的依據。表4—6分別為4種信號時域特征的對比、頻域特征對比、小波包分解各節點能量對比。
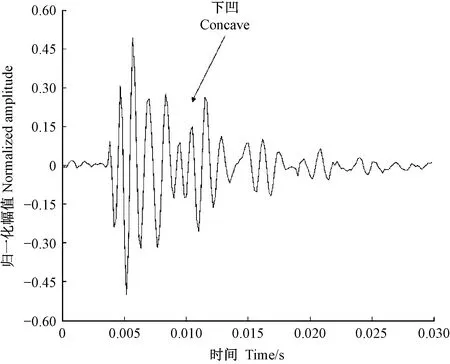
圖10 云斑天牛幼蟲蛀食白蠟樹樹干的振動信號Fig.10 The vibration signal of Batocera horsfieldi larva boring tree trunk of Fraxinus chinensis
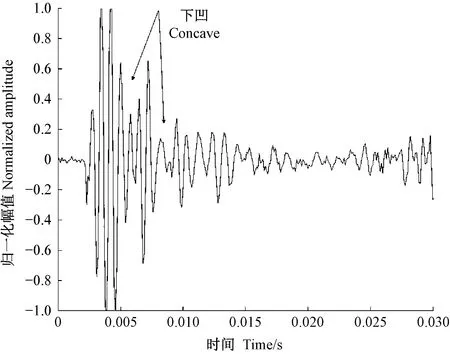
圖11 光肩星天牛幼蟲蛀食二球懸鈴木樹干的振動信號Fig.11 The vibration signal of Anoplophora glabripennis larva boring tree trunk of Platanus acerifolia
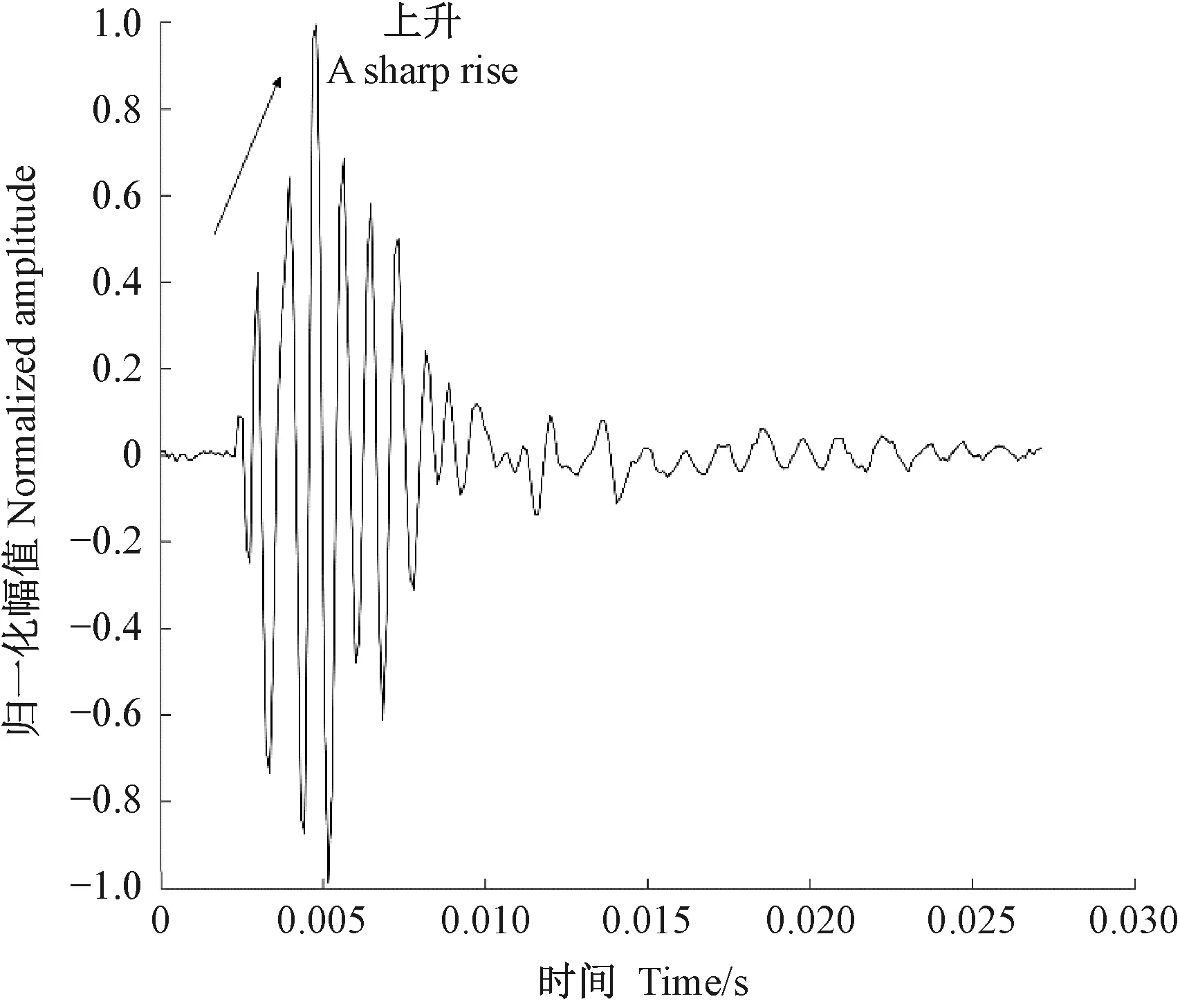
圖12 桃紅頸天牛蛀食紫葉李樹干的振動信號Fig.12 The vibration signal of Aromia bungii larva boring tree trunk of Prunus cerasifera
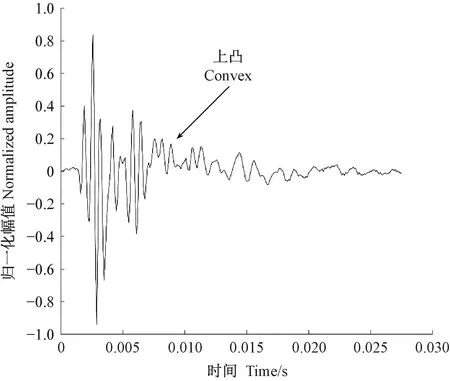
圖13 桃紅頸天牛蛀食西府海棠樹干的振動信號Fig.13 The vibration signal of Aromia bungii larva boring tree trunk of Malus micromalus
從表4可以看出,不同種類的天牛幼蟲在不同的寄主中的蛀干振動波形和持續時間存在明顯差異; 從表5可知,不同種類的天牛幼蟲在不同樹木內的蛀干振動信號頻域特征也不相同; 表6中,不同種類天牛幼蟲蛀木振動信號小波包分解各節點能量分布有明顯不同。時域特征與頻域特征相結合,可準確判斷天牛幼蟲的種類。
3 討論
天牛幼蟲蛀食樹干所引起的振動信號與木材成分、樹干含水量、樹干被蛀情況等多種因素都有關。本次試驗選取河南省新鄉市人民公園的樹木作為研究對象,同種樹木的樹齡相同,樹木生長環境基本一致。在此種情況下,先使用變分模態分解對采集得到的振動信號進行降噪處理,然后對消噪后的信號做小波包3層分解,成功提取到了天牛幼蟲蛀干振動信號的時域特征和頻域特征,可用于天牛種類的判別。

表4 3種天牛幼蟲蛀食樹干振動信號時域特征①Tab.4 Time-domain characteristics of the vibration signal of three longicorn larvae boring tree trunks

表5 3種天牛幼蟲蛀食樹干振動頻域特征Tab.5 Frequency-domain characteristics of the vibration signal of three longicorn larvae boring tree trunks

表6 3種天牛幼蟲蛀食樹干振動信號小波包分解各節點能量Tab. 6 Energy of each wavelet packet nodes of the vibration signal of three longicorn larvae boring tree trunks
天牛幼蟲在蛀道中的行為復雜,包括蛀食樹干、清理蛀道、自衛、爬行等。其中蛀食樹干為咬斷木質纖維的過程(Mankinetal., 2008),振動信號能量顯著高于其他行為,易于提取分析(卜宇飛等, 2017); 其余行為需要改進檢測儀器,開展針對性的行為觀測,提高觀測精度,才能進一步開展有效的分析研究。
天牛幼蟲的齡期與蛀干為害的強度有密切關聯,天牛幼蟲齡期對幼蟲上顎的長度和寬度有顯著影響(張海濱等, 2011)。本研究開展的時期在7月中旬,試驗中桃紅頸天牛和云斑天牛為3~4齡,光肩星天牛為低齡幼蟲,野外環境下不同的寄主和營養條件對幼蟲齡期的準確判斷造成很大困難,相關天牛幼蟲的齡期差異對蛀食振動信號的影響也需要后續研究持續關注。
4 結論
本文提出了基于害蟲蛀食樹干振動信號分析檢測識別天牛幼蟲種類的方法。該方法先用變分模態分解對采集到的振動信號進行降噪處理,再從天牛幼蟲活動引起的振動信號中挑選出蛀食樹干振動信號進行分析,利用小波包3層分解提取蛀干振動信號時域與頻域特征。 云斑天牛、光肩星天牛、桃紅頸天牛幼蟲蛀食二球懸鈴木、白蠟樹、紫葉李和西府海棠樹干的振動信號時域與頻域特征存在明顯差別,采用振動信號分析法能有效識別判斷天牛幼蟲種類。本研究為建立天牛幼蟲蛀木振動信號特征庫,實現天牛幼蟲活體快速林間檢測,制定林區天牛類蛀干害蟲防控方案提供了科學依據。
猜你喜歡
科學大眾(2023年17期)2023-10-26 07:39:14
艦船科學技術(2022年8期)2022-06-05 07:36:28
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:05:56
中國生殖健康(2019年3期)2019-02-01 06:12:26
中國公路(2017年18期)2018-01-23 03:00:38
數學物理學報(2017年6期)2018-01-22 02:26:40
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
