EMS誘變水稻創制抗咪唑啉酮除草劑種質
2021-01-28 01:37:10陳天子余月凌溪鐵張保龍
核農學報 2021年2期
陳天子 余月 凌溪鐵 張保龍
(江蘇省農業科學院/江蘇省農業生物學重點實驗室,江蘇南京 210014)
稻田雜草嚴重影響水稻產量和品質。據統計,稻田雜草致使我國稻谷每年的損失率達15%左右[1]。化學除草是目前廣泛普及的稻田除草方式,包括除草劑對土壤封閉處理和對莖葉處理[2]。化學除草有嚴格的施用范圍,使用不當會引發藥害,造成水稻減產甚至絕產[3]。因此,培育抗除草劑的水稻品種是克服稻田雜草的經濟有效途徑。轉基因技術是培育抗除草劑作物的有效途徑之一,但目前我國尚未批準轉基因水稻的商業化種植,不能在生產上推廣應用。因此培育非轉基因的抗除草劑水稻在生產上意義重大。
乙酰乳酸合成酶(acetolactate synthase,ALS)是支鏈氨基酸(纈氨酸、亮氨酸和異亮氨酸)生物合成的第一個關鍵酶,是磺酰脲類(sulfonylureas,SU)、咪唑啉酮類 ( imidazolinones,IMI )、三唑嘧啶類(triazolopyrimidines,TP)、 嘧啶氧(硫) 苯甲酸類[pyrimidinylthio (oroxy)-benzoates,PTB;pyrimidinylcarboxyherbicides,PCs]和磺酰胺基羰基三唑啉酮類(sulfonylamino-carbonyltriazolinones,SCT)等ALS 抑制劑類除草劑的作用靶標,ALS 某些氨基酸位點的突變可賦予突變體對ALS 抑制劑類除草劑的耐受性[4]。如水稻ALS 的第95、第96、第171、第548、第627 和第628 位氨基酸突變為某種氨基酸時,可對IMI、SU、PC等除草劑產生抗性[5-9]。美國路易斯安納州立大學農業中心通過甲磺酸乙酯(ethyl methanesulfonate,EMS)誘變水稻種子并結合常規育種手段,先后培育了CL121、CL131、CL141、CL161 等系列抗咪唑啉酮除草劑的非轉基因水稻,并聯合巴斯夫公司于2002年以Clearfield 品牌進行商品化種植,成功解決了美國南部水稻種植區最難治的雜草紅稻(red rice)的危害,備受當地稻農的歡迎;該水稻僅2005年種植面積就超過24 萬hm2,占美國水稻種植面積的17%[10]。其中,CL121 和CL141 抗咪唑啉酮水稻是利用其親本材料93AS3510 的ALS 第628 位氨基酸由甘氨酸突變為谷氨酸帶來的抗性,CL161 抗咪唑啉酮水稻則是利用其親本材料PWC16 的ALS 第627 位氨基酸由絲氨酸突變為天冬酰胺而產生的抗性[6,8]。
2018年,巴斯夫公司聯合路易斯安那州立大學農業中心將抗除草劑Provisia 水稻推廣上市[11],該水稻屬于抗乙酰輔酶A 羧化酶抑制劑的非轉基因水稻。生產實踐中,β -三酮類除草劑雙環磺草酮(benzobicyclon,BBC)用于防除水稻田一年生和多年生雜草已有多年,其作用靶標為4-羥基苯丙酮酸雙加氧酶(4-hydroxyphenylpyruvate dioxygenase,HPPD);但BBC 對不同水稻品種的安全性有較大差異[12]。最近,日本科學家從抗BBC 的粳稻克隆了β-三酮類除草劑抗性基因HIS1 (HPPD inhibitor sensitive1),該基因編碼一種Fe(Ⅱ)/2-氧戊二酸依賴性加氧酶,這種加氧酶通過催化β-三酮類除草劑發生羥基化反應而使之失去毒性,從而賦予植物對多種β-三酮類除草劑的抗性;HIS1 基因缺失28 bp 堿基發生無意突變后,水稻對BBC 敏感,HIS1 基因的表達是造成水稻品種對BBC抗、感差異的原因[13]。目前,我國尚沒有抗除草劑水稻品種獲得許可登記,但通過篩選人工誘變群體和自然突變群體,也獲得了一些非轉基因抗除草劑水稻材料。王俊梅等[14]對84 份經過輻照和EMS 誘變后的再雜交水稻材料進行耐草甘膦的篩選,獲得5 份耐草甘膦水稻遺傳資源。金粳818 是天津市水稻研究所利用津稻9618 和津稻1007 雜交經多年系譜法選育而成的常規品種,在水稻資源進行除草劑篩選過程中發現其具咪唑啉酮除草劑抗性[15],并具有與CL161 抗咪唑啉酮水稻同樣的突變位點,即其ALS的基因編碼區第1 880 nt 由G 突變為A,導致第627 位的絲氨酸突變為天冬酰胺[16-17],推測其抗性可能是在有殘留除草劑的大豆田中自然突變所產生[15-16]。廣東創新科研團隊最近也通過化學誘變篩選方法研發出非轉基因抗除草劑水稻材料“潔田稻”[18]。盡管如此,國內研究者創制非轉基因抗除草劑水稻的努力尚未能滿足市場的巨大需求。近年來,我國水稻輕簡化栽培發展較快,免耕和直播稻田雜草的危害日益突出[19-20]。水稻抗除草劑性狀已愈來愈受到水稻育種家和種業公司的重視。
因此,本研究通過EMS 誘變水稻種子,然后用咪唑啉酮類除草劑篩選誘變處理的后代種子,獲得了與前人報道的ALS突變類型相同的抗除草劑材料,也獲得了ALS突變的非轉基因抗咪唑啉酮類除草劑的水稻新材料,并對獲得的抗除草劑材料開展抗性水平鑒定,旨在為水稻抗除草劑育種提供材料。
1 材料與方法
1.1 材料與試劑
水稻試驗材料為秈稻93-11(由江蘇省農業種質資源保護與利用平臺提供)。EMS 為美國Sigmaaldrich 公司產品(貨號M0880);硫代硫酸鈉產自國藥集團化學試劑有限公司;百壟通為水劑型咪唑啉酮類除草劑,由巴斯夫公司生產(有效成分為240 mg·mL-1甲咪唑煙酸),苗后定向噴霧防治一年生禾本科雜草的推薦用藥量為每畝20 ~30 mL,如按每畝兌水45 L計,甲咪唑煙酸的推薦使用濃度范圍為107 ~160 mg·L-1。
1.2 水稻EMS 誘變處理和抗除草劑突變體篩選
取水稻種子20 kg,自來水浸泡2 h 后,用0.5%(w/v) EMS 室溫下浸泡14 h,加入終濃度為25 g·L-1的硫代硫酸鈉中和反應15 min,棄去EMS 溶液,自來水翻動浸泡種子5 次,每次5 min。然后將處理后的種子播種田間,M1植株成熟后,種子混收。取M2種子播種于溫室,播后10 d 后對M2幼苗噴施0.67 mL·L-1百壟通,即相當于160 mg·L-1甲咪唑煙酸,20 d 后選擇正常綠色且明顯長高的抗性株移栽至花盆,收取M3種子。
1.3 抗除草劑突變體ALS 基因克隆與序列分析
根據NCBI 水稻ALS基因保守序列設計擴增ALS基因全長的特異引物OsALS-F 5′-TCGCCCAAACCCAG AAACCC-3′和OsALS-R 5′-CTCTTTATGGGTCATTCAG GTC-3′。以M3抗除草劑單株基因組DNA 為模板,采用TaKaRa PrimerSTAR Max DNA Polymerase 聚合酶擴增ALS基因。PCR 反應體系包括:2×PrimerSTAR Max Premix 10.0 μL、10 μmol·L-1引物各1.0 μL、30 ng·μL-1水稻基因組DNA 1.0 μL,補加無菌水至總體積20 μL。PCR 擴增程序:98℃預變性3 min;98℃變性10 s,58℃退火15 s,72℃延伸2 min,35 個循環;72℃延伸10 min。取5 μL PCR 產物經1%瓊脂糖凝膠電泳檢測,發現有預期2 051 bp 片段后,剩余PCR產物經PCR 清潔試劑盒(美國Axygen 公司)清潔回收后,克隆至pMD19-T 載體(日本TaKaRa 公司),然后轉化大腸桿菌。每個轉化隨機挑取12 個大腸桿菌單克隆進行PCR 檢測,取PCR 結果呈陽性的3 個單克隆,送南京一道生物科技有限公司測序。測序結果采用BioEdit 和SnapGene 軟件分析野生型和突變體ALS基因的DNA 序列差異,確定抗除草劑突變體的ALS突變位點。
1.4 抗除草劑突變體的抗性水平和遺傳穩定性鑒定
取3 個抗性株系(編號M133、M135 和M142)M3種子和野生型(WT)水稻93-11 種子,播種紙杯中,每杯12~15 粒種子。在3~4 葉幼苗期時,以不同濃度甲咪唑煙酸(0、12、24、48、72、96、120、240、600、1 200、2 400、 4 800、 7 200、9 600、12 000 mg·L-1)水溶液進行莖葉噴霧處理,每個濃度重復3 次;噴藥后21 d 測量株高,并以對照(0 mg·L-1甲咪唑煙酸處理)為基準計算株高抑制率。株高抑制率=(對照組株高-處理組株高)/對照組株高×100%,然后按單向分組組內觀察值數目不等資料對每個濃度下的株高抑制率進行方差分析。
1.5 ALS 活性測定
取M142 抗性株系M3和野生型93-11 水稻葉片2~5 g,液氮研磨成粉末,按1 ∶1(w/v)加入提取液[100 mmol·L-1磷酸緩沖液(pH 值7.5),1 mmol·L-1丙酮酸鈉、5 mmol·L-1MgCl2、0.5 mmol·L-1焦磷酸硫胺素、1 mmol·L-1二硫代蘇糖醇],待解凍后繼續研磨勻漿。用移液器將溶解物轉移置冰預冷離心管,12 000 r·min-1、 4℃離心30 min,吸取上清液至新的冰預冷離心管中,獲得粗酶液,并置于冰上,立即用于酶活測定。ALS 活性的測定參照陳以峰等[21]的方法。在2 mL 離心管中依次加入700 ~800 μL 反應緩沖液[50 mmol·L-1磷酸緩沖液(pH 值7.0),200 mmol·L-1丙酮酸鈉、1 mmol·L-1MgCl2、1 mmol·L-1焦磷酸硫胺素]、1~100 μL 不同濃度的甲咪唑煙酸水溶液、100 μL 粗酶液;樣品管總體積為900 μL,混勻。對照管先加50 μL 3 mol·L-1H2SO4,再加入粗酶液。37℃水浴1 h,樣品管加50 μL 3 mol·L-1H2SO4,混勻,60℃孵育15 min,依次加入500 μL 0.83 g·L-1肌酸、500 μL 8.3 g·L-11-萘酚,混勻后60℃水浴15 min,37℃水浴15 min。短暫離心后取上清在ScanDrop 250 分光光度計(德國Analytik Jena 公司)上讀取400~640 nm 范圍的吸光值,取其最大值計算甲咪唑煙酸各濃度下的ALS相對酶活力及ALS 酶活抑制率。ALS 相對酶活力=甲咪唑煙酸加入組的ALS 吸光值/無甲咪唑煙酸加入組的ALS 吸光值×100%,ALS 酶活抑制率=(無甲咪唑煙酸加入組的ALS 吸光值-甲咪唑煙酸加入組的ALS 吸光值)/無甲咪唑煙酸加入組的ALS 吸光值×100%。按農業行業標準NY/T 1155.8-2007[22]和趙斌等[23]方法,以SPSS 統計軟件計算甲咪唑煙酸抑制水稻ALS酶活力至50%時的IC50值。
2 結果與分析
2.1 抗除草劑突變體篩選
甲咪唑煙酸藥效慢,噴施百壟通4 d 后,水稻M2幼苗仍無藥害癥狀;噴施10 d 后,絕大部分M2幼苗葉片變淺綠色,有極少數幼苗葉片仍呈深綠色,且植株明顯增高;噴施15 d 后,非抗性苗葉片明顯失綠、停止生長甚至枯死,抗性苗正常生長,葉片深綠色,株高增長明顯(圖1)。篩選M2種子約50 kg,共計獲得61 個抗性M2單株,成熟后單株收種。

圖1 甲咪唑煙酸(160 mg·L-1)莖葉噴霧15 d 后的M2 抗性株Fig.1 The imidazolinone resistant plantlets in M2 population at 15 d of spray with 160 mg·L-1 imazapic
2.2 ALS 基因序列分析
已知植物對咪唑啉酮類除草劑的抗性是源于ALS基因的突變[4]。本研究利用引物組合OsALS-F/OsALS-R 擴增了野生型和抗百壟通除草劑61 個M2株系的M3單株ALS基因序列,每株系各測試3 ~4 個M3單株,獲得的特異片段為2 051 bp,包括1 935 bp的ALS開放閱讀框。與野生型ALS的開放閱讀框序列相比,突變體ALS基因突變類型可分為三類(表1):1)20 個測序株系在第1 880 nt 由G 堿基突變為A 堿基(G1880A),導致編碼的第627 位氨基酸由絲氨酸(AGT)突變為天冬酰胺(AAT),為已報道的突變類型;2)23 個測序株系在第1 879、第1 880 nt 由AG 堿基突變為CT 堿基(A1879C、G1880T),導致編碼的第627 位氨基酸由絲氨酸(AGT)突變為亮氨酸(CTT),為新發現的突變類型;3)1 個測序株系檢測出同時存在上述兩種突變,即A1879C、G1880T 和G1880A,表明該單株ALS基因還未純合。剩余17 個測序株系中,12個測序株系含有G1880A 突變外,還各檢測到1 個或2個單核苷酸多態性(single nucleotide polymorphisms,SNP) 或 插 入 缺 失(insertion-deletion,InDel) 突 變(C731A、 C531T、 G1763A、 C429T、 G896A、 C414T、G254A、A1409G、C651A、G1888A、C927T、G216A、1 296~1 297 nt 間插入T 堿基);5 個測序株系含有A1879C、G1880T 突變外,還各檢測到1 個SNP 突變或InDel 突變(T1547C、G198A、C1474T、G1888A、1 330 nt處缺失G 堿基)。由此可推斷突變體抗除草劑功能的獲得是由于ALS 第627 位氨基酸由絲氨酸(AGT)突變為亮氨酸(CTT)或天冬酰胺(AAT)所致。
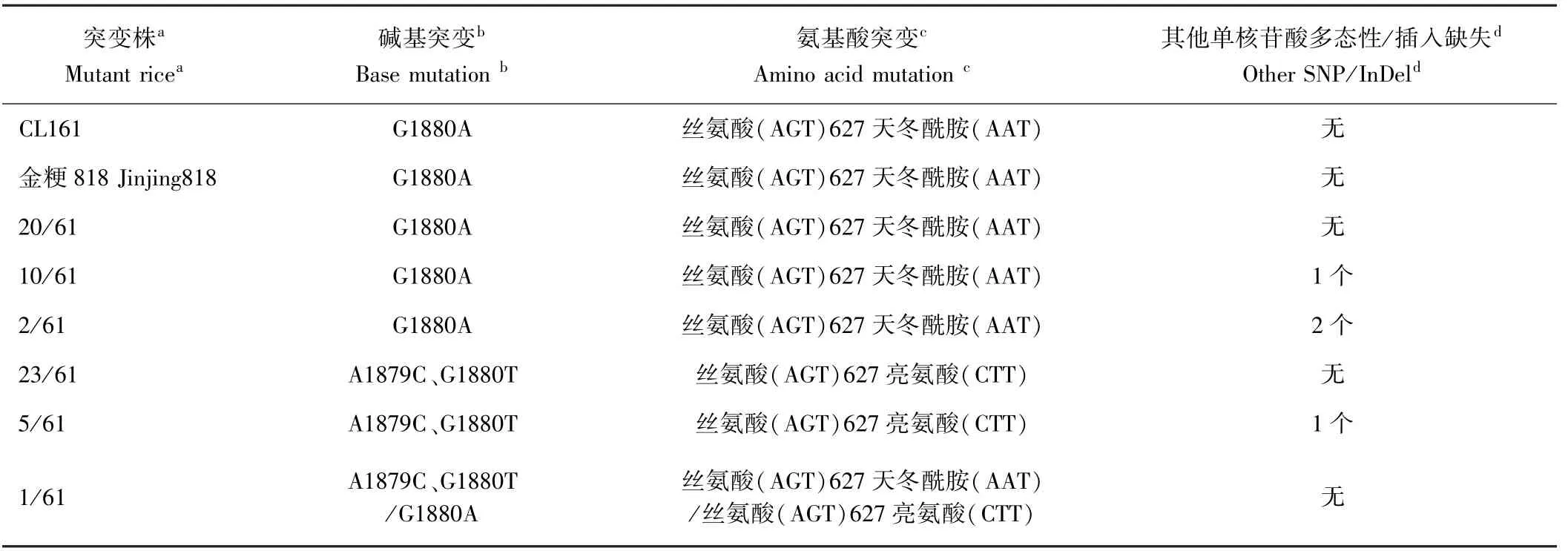
表1 61 個抗咪唑啉酮水稻株系在M3 的ALS 基因型分類Table 1 The genotyping of ALS genes in the M3 generation of sixty-one imidazolidone-resistant rice lines
2.3 抗性水平和遺傳穩定性鑒定
取3 個抗性株系 M133 (堿基突變類型為G1880A)、M135(堿基突變類型為A1879C、G1880T)和M142(堿基突變類型為A1879C、G1880T/G1880A)后代進行抗性鑒定。莖葉噴霧15 d 后,野生型水稻幼苗經12 mg·L-1甲咪唑煙酸噴霧處理后產生明顯的生長抑制、葉片枯蔫等藥害癥狀,且藥害癥狀隨除草劑濃度的升高而加重,而抗性株在1 200 mg·L-1甲咪唑煙酸噴霧后才產生輕微葉片枯蔫癥狀(圖2-A)。抗性株經12~96 mg·L-1甲咪唑煙酸噴霧21 d 后,反而促進生長,當噴霧濃度達120 mg·L-1時才出現生長抑制現象(圖2-B)。野生型水稻幼苗莖葉經14 個甲咪唑煙酸濃度噴霧30 d 后,所有幼苗全部死亡;抗性株M133在1 200 mg·L-1甲咪唑煙酸噴霧后死亡;抗性株M135和M142 在1 200 mg·L-1甲咪唑煙酸噴霧后生長被抑制,濃度達2 400 mg·L-1時植株死亡(圖2-C)。這表明抗性水稻對甲咪唑煙酸的抗性能穩定遺傳且其抗性水平不低于1 200 mg·L-1甲咪唑煙酸,其對甲咪唑煙酸的抗性是野生型的100 倍以上。此外,農藝性狀測定結果表明,抗性株系M135、M142 的每穗粒數比野生型多出11.5%~13.4%,在株高、穗長、千粒重、抽穗期、結實率等農藝性狀上與野生型無明顯差異(表2),具有較好的應用價值。
2.4 ALS 酶活測定
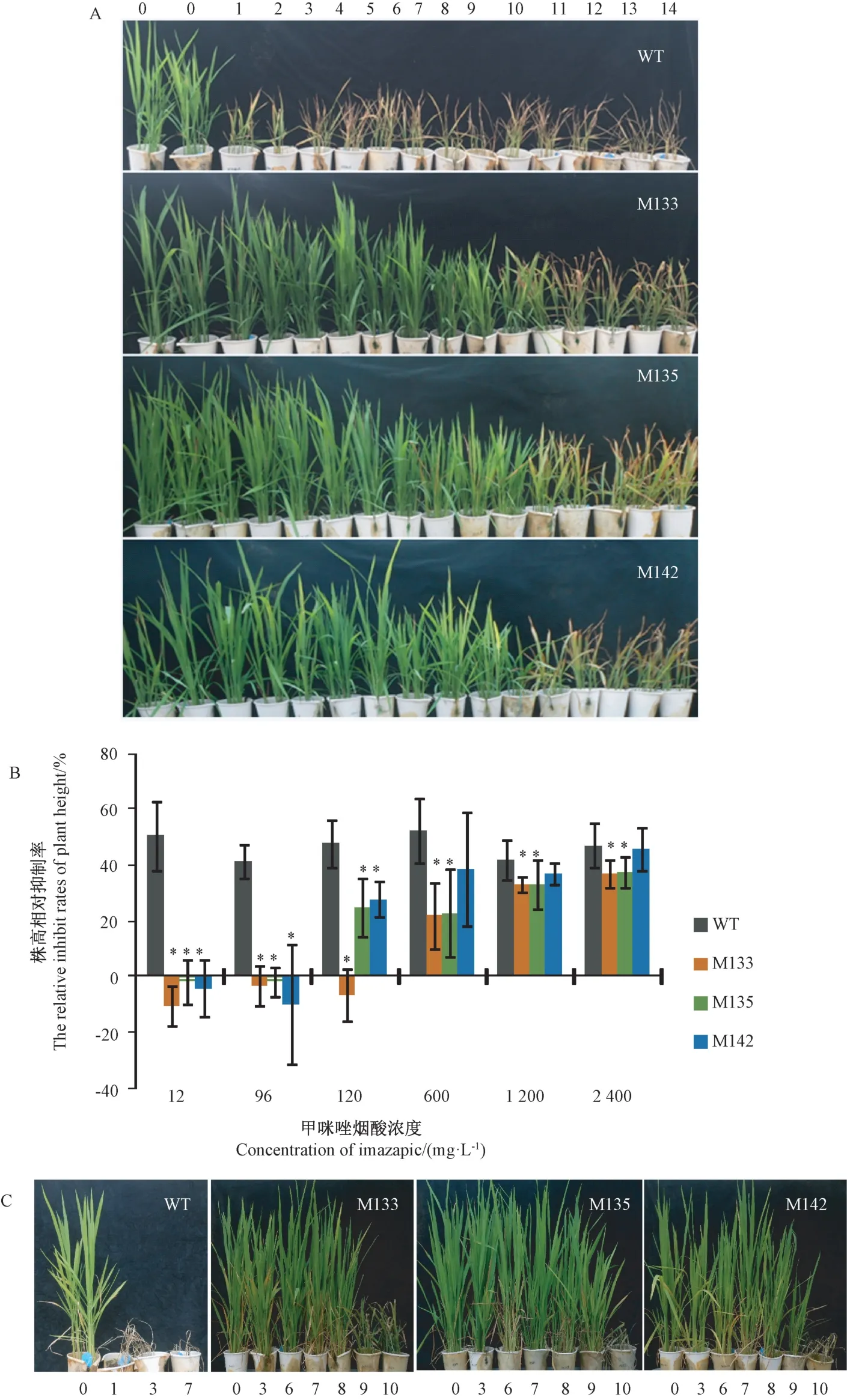
圖2 M3 抗咪唑啉酮除草劑水稻的抗性水平Fig.2 The resistance level of M3 imidazolinone-resistant rice
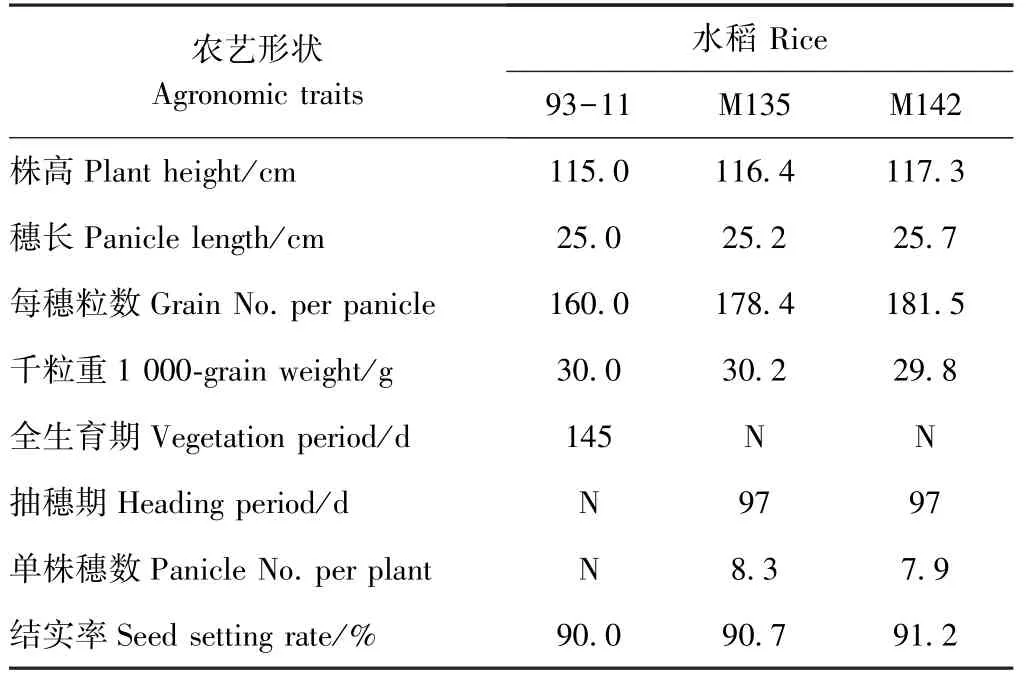
表2 抗咪唑啉酮除草劑水稻的農藝性狀Table 2 The agronomic traits of the imidazolidone-resistant rice
ALS 是ALS 抑制劑除草劑的作用靶標,其活性水平是鑒定植物對ALS 抑制劑抗性的重要生化指標。在M142 抗性株葉片粗酶液與10 μmol·L-1甲咪唑煙酸孵育后,產生淡紅色產物,而野生型水稻葉片粗酶液產生黃色產物(圖3-A),表明M142 在10 μmol·L-1甲咪唑煙酸下仍具有較高的ALS 酶活性。反應產物在400~640 nm 下掃描測定吸收值,結果顯示水稻ALS的紅色復合物最大吸收峰在530 nm (圖3-B)。取水稻粗酶液與甲咪唑煙酸各濃度孵育后的530 nm 吸收值計算ALS 相對酶活力,結果顯示(圖3-C),野生型水稻的ALS 對甲咪唑煙酸極為敏感,ALS 酶活在0.01 μmol·L-1開始受到抑制,并隨甲咪唑煙酸濃度的增加酶活性迅速降低,在甲咪唑煙酸濃度為10 μmol·L-1時ALS 相對酶活力為15.15%;而M142 抗性株的ALS 對甲咪唑煙酸的耐受性明顯強于野生型水稻,在甲咪唑煙酸濃度為10 μmol·L-1時開始受到抑制,在濃度為10 000 μmol·L-1時ALS 相對酶活力為14.42%。以SPSS 軟件對水稻ALS 酶活抑制率的幾率值和甲咪唑煙酸濃度的對數值進行直線回歸分析,求得野生型水稻IC50=2.037 μmol·L-1,M142 抗性株IC50=222.099 μmol·L-1,即M142 抗性株對甲咪唑煙酸的耐受性是野生型水稻的109 倍。
3 討論
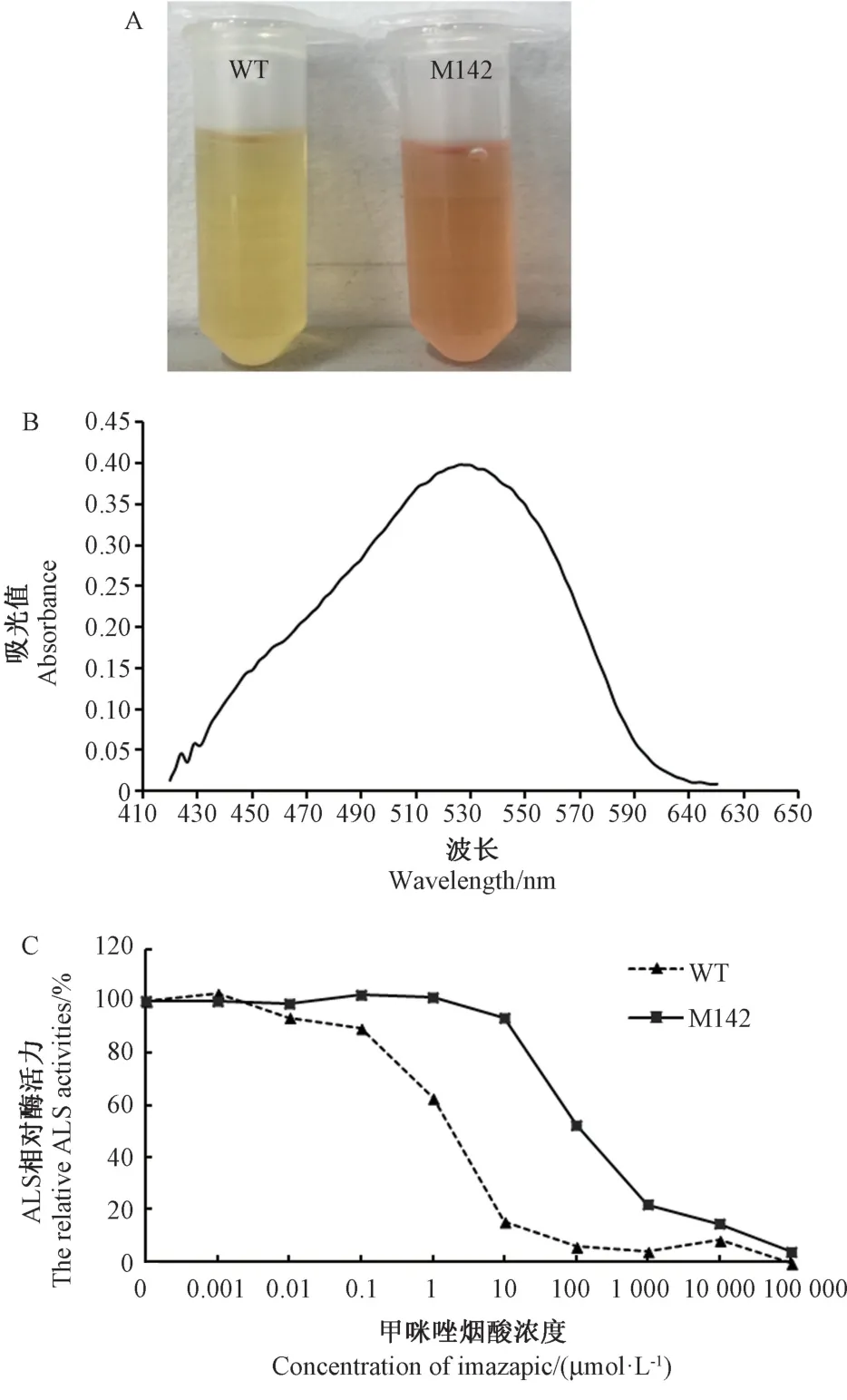
圖3 水稻ALS 酶活性測定Fig.3 The enzyme assay of rice ALS
目前絕大部分抗ALS 抑制劑作物的抗性機制是源于ALS基因堿基突變造成氨基酸殘基位點變異,引起ALS 抑制劑(除草劑)與ALS 蛋白結合性降低,從而使得突變表對該除草劑體現為不敏感或耐受性[4,24],也有個別報道由細胞色素P450 單加氧酶基因CYP72A31、CYP81A6 等編碼代謝酶降解除草劑所致[25-28]。本研究在對抗除草劑突變體進行抗性篩選和抗性表型進行鑒定的基礎上,對部分抗除草劑突變體進行了基因型鑒定,確認這些突變體是在水稻ALS蛋白第627 位絲氨酸(AGT) 被突變為天冬酰胺(AAT)或亮氨酸(CTT)。水稻ALS 第627 位絲氨酸對應于擬南芥ALS 第653 位絲氨酸。該位點氨基酸發生突變后獲得抗ALS 抑制劑除草劑的功能在許多作物和雜草中都已被報道,包括水稻[6,8]、油菜[6,24]、玉米[6]、小麥[6]等,其除草劑抗性功能都是源于單堿基突變造成ALS 第653 位絲氨酸(以擬南芥ALS 氨基酸序列位置為標準)突變為天冬酰胺、蘇氨酸、苯丙氨酸、異亮氨酸[4]。如前所述,CL161 和金粳818 抗性水稻在該位點的氨基酸突變類型是絲氨酸突變為天冬酰胺[6,10,17]。本研究還發現水稻ALS 第627 位絲氨酸的2 個堿基AG 同時突變為CT 后,造成絲氨酸變為亮氨酸,突變體也具有抗咪唑啉酮類除草劑功能,該位點產生亮氨酸突變類型此前鮮見報道。ALS 抗性突變位點的不同、替換氨基酸殘基種類的不同,導致其蛋白結構不同,其抗除草劑特點也不同[29],如含627 天冬酰胺突變的CL161 對咪唑啉酮除草劑的抗性是CL121(含628 谷胺酸突變)的5 倍[30]。隨著對更多EMS 誘變水稻材料進行抗ALS 抑制劑類除草劑篩選和基因型分析,還可能發現ALS 基因產生其他新的氨基酸突變類型能賦予抗除草劑功能。
抗草甘膦轉基因農作物的大面積推廣表明轉基因是培育抗除草劑作物的有效途徑之一。但轉基因作物自面世以來,其環境安全和食品安全等問題一直備受公共部門、學術界和普通公眾的廣泛關注,引發的爭議也在一定程度上阻礙了轉基因作物的發展。如我國轉植酸酶基因玉米和轉Bt 基因抗蟲水稻在2009年已獲得生產應用安全證書,但至今尚未進行商業化生產,其原因之一是基于國內有關轉基因的輿論氛圍,以及我國對轉基因主糧作物產業化進程穩重、謹慎的考慮[31]。所以,我國目前仍未批準抗除草劑轉基因水稻的商業化。相比之下,傳統育種手段培育的作物品種由于沒有導入外源物種基因,更容易被接受和推廣,如抗咪唑啉酮油菜及油菜籽可以自由進入歐盟及其他國家[32],抗咪唑啉酮除草劑的非轉基因Clearfield 水稻在美國也產生了良好的經濟效益[10]。EMS 屬于常用的一種化學誘變劑,可產生豐富的基因組突變及表型突變類型,廣泛用于植物誘變育種[33-34]。本研究通過EMS 誘變水稻93-11 種子篩選到的抗咪唑啉酮除草劑水稻能耐受不低于1 200 mg·L-1甲咪唑煙酸,而野生型水稻在甲咪唑煙酸濃度為12 mg·L-1時就已死亡,表明所獲得抗性水稻對甲咪唑煙酸的抗性水平是野生型的100 倍以上。ALS 酶活測定結果也揭示抗性水稻對甲咪唑煙酸的耐受性是野生型的109 倍。該抗除草劑性狀對咪唑啉酮抗性倍數高,且能穩定遺傳,具備生產應用價值。利用同樣的手段,在華占、1892S 不育系、嘉花1 號、淮稻5 號、臨稻16 號、鎮稻18 號、鎮糯19 號、圣稻19 號、蘇秀867 等多個水稻品種上都成功獲得抗咪唑啉酮除草劑的突變材料。它們對咪唑啉酮除草劑的抗性源于水稻基因組堿基突變產生,無轉基因環境釋放的安全性問題,在控制免耕和直播稻田雜草方面具有廣闊的推廣應用前景。此外,本研究發現低濃度百壟通莖葉噴霧處理對抗性水稻的生長有促進作用的現象。該現象是由除草劑產品本身還是由抗性突變體自身基因型引發產生還有待進一步驗證和確認。
4 結論
本研究通過EMS 誘變獲得的抗咪唑啉酮水稻突變體材料可耐受不低于1 200 mg·L-1甲咪唑煙酸,其抗性水平是野生型的100 倍以上,抗性倍數高,且能穩定遺傳,具有生產應用價值。其抗除草劑功能源于ALS基因開放閱讀框在第1 879、第1 880 nt 由AG 堿基突變為CT 堿基,或在第1 880 nt 由G 堿基突變為A堿基,導致編碼的第627 位氨基酸由絲氨酸(AGT)突變為亮氨酸(CTT)或天冬酰胺(AAT);其中第627 位氨基酸由絲氨酸(AGT)突變為亮氨酸(CTT)鮮有報道。突變后的水稻ALS 對咪唑啉酮類除草劑的耐受性顯著提高,其IC50是野生型水稻的100 倍以上。在當前我國嚴格監管轉基因水稻的背景下,該非轉基因的抗除草劑水稻具有廣闊的應用前景。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
現代農業(2016年5期)2016-02-28 18:42:36
營銷界(2015年23期)2015-02-28 22:06:18
