植物群落和土壤對人工草地恢復演替的協同響應
2021-01-28 14:00:54潘攀王長庭胡雷劉斯莉李潔
生態環境學報 2020年12期
關鍵詞:植物
潘攀,王長庭*,胡雷,劉斯莉,李潔
1. 西南民族大學青藏高原研究院,四川 成都 610225;2. 西南民族大學生命科學與技術學院,四川 成都 610225
三江源區作為青藏高原的腹地,是高寒草甸的集中分布區(王琰,2015),近年來由于受到氣候變化(全球變暖)、鼠蟲危害及人類活動(過度放牧)的影響,90%以上的草地出現了不同程度的退化(李穗英等,2007),部分嚴重退化為“黑土灘”,對當地和下游地區生態安全、社會和經濟的可持續發展造成了惡劣的影響(Dong et al.,2010)。為了遏制高寒草甸的退化,目前已有許多的修復技術應用于該地區(例如圍封、禁牧、施肥等),但由于高寒嚴酷自然條件的制約,僅靠單一的禁牧封育在短期內難以恢復“黑土灘”退化草地,只有通過建植人工草地等人為措施的干擾才能實現草地的快速恢復或重建(史惠蘭等,2005)。而建植人工草地需經歷物種入侵、定居繁殖到群落穩定的復雜生態過程,且由于草種相對單一,生態位集中,容易造成競爭,波動性大(Britton-Simmons et al.,2008)。研究指出,在人工草地演替過程中,輔助以人工干預手段仍然需要 12年其地上植物群落生產力、多樣性才能與原始植物群落相似,土壤環境12年后也仍然難以達到原始狀態(Baer et al.,2002)。其中在恢復7—9年,采用多年生禾本科植物建植的人工草地植物豐富度、蓋度、生物量下降,草地易發生逆向演替,出現二次退化現象(Dong et al.,2012;吳曉慧等,2019)。但也有研究表明,恢復演替8年后,人工草地植被開始達到第一個相對穩定的階段,有較好的恢復效果,隨著恢復年限的增加,植物群落和不同功能群物種豐富度和多樣性開始階段性波動,并在建植16 a左右,人工草地植物群落高度和蓋度顯著下降(武勝男等,2019)。實質上,人工草地的恢復演替過程是物種多樣性不斷變化、優勢物種不斷更替、植物物種生活型不斷演變的過程。在該過程中,不僅地上植物群落發生顯著變化,土壤養分狀況(王學霞等,2012)、土壤酶活性(王啟蘭等,2010),以及連接植物和土壤的根系(字洪標等,2016;樊博等,2020),包括生物量、分布格局和根土比也隨之發生變化。植物根系、土壤理化性質、土壤酶活性等的變化又反作用于地上植物群落,協同影響地上植物群落的變化,從而決定了人工草地的恢復演替過程(王啟蘭等,2009)。
目前關于人工草地生態系統長時間序列演替的規律仍存在較大爭議,尤其是人工草地恢復演替過程與地下生態過程的互作機制尚不清楚。而將地上植物群落與土壤環境,特別是地下根系相結合,進行地上—地下耦合演替恢復機制的研究,才有可能準確評估和預測人工草地演替方向、動態過程和恢復的時間閾值。因為地下土壤指標驅動著地上植物群落的變化,是人工草地演替的重要驅動因子,在一定程度上決定了人工草地的演替方向(張錦華,2006)。如土壤含水量和養分供應能增加植物地下生物量并提高植物生產力(Chapin et al.,1987;Yuan et al.,2010;Chen et al.,2018);其中氮素是限制植物生長最主要的因子,氮素的增加會促進少數植物的生長,從而減少植物多樣性(Stevens et al.,2004);土壤酶作為土壤的重要組成部分,對土地利用方式和管理措施的改變反應迅速,土壤酶活性在一定程度上表征了土壤微生物分解和利用土壤養分的能力(馬榮榮,2015),間接影響了地上植物群落的養分供給,且酶活性也是退化草地土壤恢復的重要指標之一(楊殿林等,2020);同時,根系是土壤和植物的動態界面,是土壤資源的直接利用者和產量的重要貢獻者(周本智等,2007),地下根系的吸收和轉運決定了地上植物群落的生長狀況,影響生態系統的演替進程(王長庭等,2012)。其中根土比(R/S)作為衡量根系在土壤中的生物量大小和空間分布規律的重要指標,其大小反映出不同退化演替階段高寒草地生態系統地下根系和土壤“載體”量的動態變化趨勢(字洪標等,2016)。因此,本研究選擇了三江源區不同建植期的人工恢復草地,測定了人工草地土壤理化性質、酶活性、根土比和地上植物群落特征等參數,利用冗余分析(RDA)和Pearson相關等進行綜合分析,旨在研究:(1)不同建植年限人工草地植物群落的動態演替特征及其驅動因子;(2)不同建植年限人工草地植物群落和土壤環境的協同響應。
1 研究區概況
研究區位于青海省果洛藏族自治州瑪沁縣,地理坐標 33°34′—34°30′N,99°54′—100°29′E,平均海拔4150 m。該區屬于典型的高原大陸性氣候,春季干旱多風,冬季寒冷漫長。年平均氣溫為-1.7 ℃,最冷月1月平均氣溫-14.8 ℃,最熱月7月平均氣溫9.8 ℃。年平均降雨量600 mm,主要降水集中在5—9月,約占年降水量的 80%。土壤類型為高山草甸土,高寒嵩草草甸為該地區主要的草地類型。由于多年來的氣候變化、鼠害肆虐,加之草地管理制度不健全、過度放牧等問題,導致其原生植被遭到不同程度的破壞,草地呈現出大面積退化的趨勢。
2 研究方法
2.1 試驗樣地設計
選擇青海省果洛藏族自治州瑪沁縣大武鎮軍牧場的 4個面積為 20 hm2,建植年限分別為 5 a(34°28′10′′N,100°21′01′′E,海拔 3740 m)、6 a(34°28′09′′N,100°21′03′′E,海拔 3738 m)、9 a(34°27′41′′N,100°12′35′′E,海拔 3757 m)、13 a(34°21′45′′N,100°29′58′′E,海拔 3949 m)的人工草地為試驗樣地,4塊人工草地均為垂穗披堿草(Elymus nutans)、冷地早熟禾(Poa crymophila)、中華羊茅(Festuca sinensis)和星星草(Puccinellia tenuiflora)按照 4∶1∶0.5∶0.5(每公頃播種重量比)的比例混播,播種量為72 kg·hm-2,千粒重分別為:垂穗披堿草:2.46 g、冷地早熟禾:0.22 g、中華羊茅:0.67 g、星星草:0.61 g。播種前先清除試驗地雜草,翻耕土地,精細平整,播種后覆土1—2 cm,并以150 kg·hm-2混合肥作基肥。
2.2 植物樣品采集和測定
于2015年植物生長旺季(8月中旬),在4塊樣地內分別隨機選取3個面積為50 m×50 m的平行樣地,在每個平行樣地內隨機選擇6個50 cm×50 cm的小樣方進行植被群落調查。在各個小樣方內記錄樣方總蓋度及所有植物物種數。植被調查結束后,將地上植被按物種齊平地面刈割,分裝進信封,帶回實驗室60 ℃、48 h烘干至恒質量,并稱質量(Li et al.,2014)。于2015年10月進行凋落物采集,在4塊樣地內分別隨機選取3個面積為50 m×50 m的平行樣地,在每個平行樣地內隨機選擇 6個 50 cm×50 cm的小樣方進行凋落物的收集,帶回實驗室60 ℃、48 h烘干至恒質量,并稱質量。本試驗選擇Shannon- Wiener多樣性指數和Simpson多樣性指數對植物多樣性進行評價。各指標計算公式(張金屯,2004)如下:
Shannon-Wiener多樣性指數:

Simpson多樣性指數:

式中,Pi為種i的個體數與群落中全部物種個體數的比值;S為群落中物種數目。
2.3 土壤樣品采集和測定
植被群落調查結束后,在選取的每個平行樣地內用內徑5 cm的土鉆分層(0—10、10—20 cm)按“S”型鉆取5鉆土壤,均勻混合為1個土壤樣品,每個樣地3個重復,做好標記,放入有冰袋的采集箱中運回實驗室。其中一部分樣品過1 mm篩后保存于4 ℃冰箱用于酶活性及速效養分的測定,另一部分樣品經風干后過0.15 mm篩用于測定土壤養分(鮑士旦,2000)14-21。
根土質量比采用土鉆法(任繼周,1998),在每個觀測樣方,用內徑5 cm土鉆在0—10 cm和10—20 cm土層取樣,每層取5鉆,重復3次,樣品過篩得到土壤樣品并稱質量,清水沖洗得到根系樣品,分別在60 ℃和105 ℃烘至恒質量,稱干質量,根土質量比=根質量/土壤質量。用環刀法測定 0—10 cm和10—20 cm土壤容重(SBD)(孔凡偉,2010),每個樣地重復3次。土壤含水量(SWC)測定采用烘干法;土壤pH測定采用電位法,水土比為2.5∶1;采用重鉻酸鉀氧化滴定法測定土壤有機質(SOM),凱氏定氮法測定全氮(TN),鉬銻抗比色法測定全磷(TP),碳酸氫鈉浸提-鉬銻抗比色法測定速效磷(AP),原子吸收分光光度法測定全鉀(TK),四苯硼鈉法測定速效鉀(AK)(鮑士旦,2000)22-108。
采用比色法測定土壤蔗糖酶、纖維素酶、酸性磷酸酶、脲酶、硝酸還原酶和過氧化氫酶的活性(關松萌,1986;王榛等,2018),酶活性測定均進行3次重復。
2.4 數據處理與分析
原始數據采用Excel 2007進行整理,采用IBM SPSS Statistics 21.0進行統計學分析,對土壤理化性質和植物群落指標(多樣性、豐富度、地上生物量、蓋度和凋落物)進行單因素方差分析,不滿足方差齊性的指標先進行log函數轉換,滿足條件后用最小顯著極差法(LSD)進行不同處理間的差異顯著性檢驗(P<0.05),不同土層間的差異則采用獨立樣本t檢驗;用線性回歸分析對土壤酶活性與建植年限進行一次、二次擬合,最佳模型以最小赤池信息值(AIC)為準;用Canoco 4.5對植物群落特征與土壤因子的相互作用關系進行排序分析,在降趨對應分析(DCA)得到第1軸梯度最大為0.144(小于3)的前提下選擇冗余分析(RDA);用Pearson相關性分析土壤理化性質與植物群落特征和酶活性之間的相關性;采用Origin 8.5進行繪圖。
3 結果與分析
3.1 不同建植年限人工草地植物群落特征的變化
根據研究區不同建植年限人工草地植物群落特征的變化(圖 1)所示,隨建植年限的增加,Simpson指數、Shannon-Wiener指數、物種豐富度先降低后增加,地上生物量、蓋度和凋落物呈“N”字型變化。Simpson指數和Shannon-Wiener指數在建植6年顯著低于其他年限,分別為0.85和1.32;物種豐富度、地上生物量和凋落物在建植 13年顯著增加,分別為 8.67、374.63、44.09 g·m-2。
3.2 不同建植年限人工草地土壤理化性質的變化
根據研究區不同建植年限人工草地土壤理化性質的變化可知(表1),隨著建植年限的增加,0—10 cm和10—20 cm土層的R/S均不斷增加,在建植13年分別達到該土層最高的3.58和1.17。SWC在0—10 cm土層呈“N”字型變化,在10—20 cm土層呈“V”字型變化,其中兩個土層在建植9年的SWC均為各處理中最低,分別為7.81%和7.08%。土壤pH隨著建植年限的增加呈現先增加后降低的趨勢,且各處理之間差異顯著(P<0.05),在 0—10 cm和10—20 cm土層中均在建植13年達到最低的6.13和6.95。就土層間而言,隨著土層增加,R/S顯著降低(P<0.05),土壤pH顯著升高(P<0.05),SWC呈降低趨勢,但土層間差異不顯著(P>0.05)。SBD在不同建植年限及不同土層中均無顯著差異(P>0.05),變化范圍在 1.35—1.49 g·cm-3。

表1 不同建植年限人工草地土壤理化性質的變化Table 1 Change of soil physicochemical properties in artificial grassland of different cultivation periods
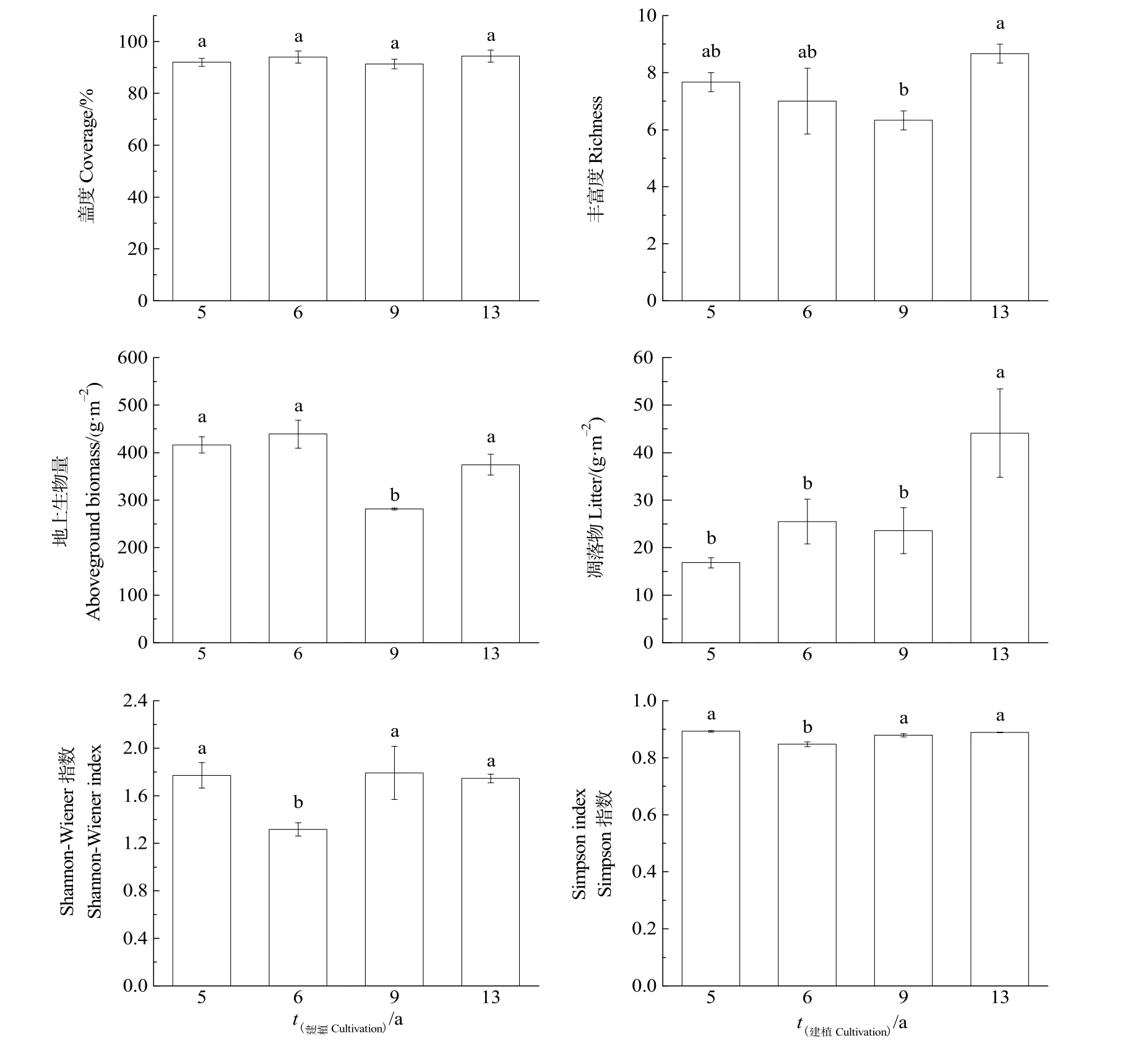
圖1 不同建植年限人工草地植物群落特征的變化Fig. 1 Change of plant community characteristics in artificial grassland of different cultivation periods
在0—10 cm土層,隨建植年限的增加,SOM呈“V”字型變化;土壤 TN、TP、TK、AP、AK呈“N”字型變化,且SOM、TN、TP、TK及AP均在建植9年達到最小值,分別為75.67、5.45、0.54、18.84 g·kg-1及 3.73 mg·kg-1。在 10—20 cm 土層,隨建植年限的增長,SOM、TN、TK呈“V”字型變化;TP、AP呈“N”字型變化;AK不斷增加。且SOM、TN、TP及AP均在建植9年達到最小值,分別為 62.47、5.09、0.63 g·kg-1及 3.08 mg·kg-1。就土層間比較而言,SOM、TN及AK隨土層加深顯著降低(表1)。
3.3 不同建植年限人工草地土壤酶活性的變化
對人工草地建植年限與土壤酶活性進行擬合,最優擬合函數均為二次函數(圖 2,表 2)。在 0—10 cm土層,隨建植年限的增加,土壤蔗糖酶、纖維素酶、酸性磷酸酶、脲酶、硝酸還原酶和過氧化氫酶活性先降低后增加,其中脲酶、硝酸還原酶和過氧化氫酶活性與建植年限有較強的相關性(R2>0.7);蔗糖酶、酸性磷酸酶、脲酶、硝酸還原酶和過氧化氫酶活性均以建植 9年最低,分別為1583.40 mg·g-1·h-1、2.33 mg·g-1·h-1、0.34 mg·g-1·h-1、0.027 μg·g-1·h-1、24.07 μg·g-1·h-1(平均值)。在 10—20 cm土層,隨建植年限的增加,蔗糖酶、脲酶、硝酸還原酶和過氧化氫酶活性先降低后增加,而纖維素酶、酸性磷酸酶活性先增加后降低,其中脲酶、硝酸還原酶和過氧化氫酶活性與建植年限的相關性最強(R2>0.7);蔗糖酶、脲酶和硝酸還原酶活性以建植 9 年最低,分別為 1388.52 mg·g-1·h-1、0.36 mg·g-1·h-1、0.050 μg·g-1·h-1。

圖2 土壤酶活性隨建植年限增加的變化規律Fig. 2 Change of soil enzyme activities with the increase of planting years
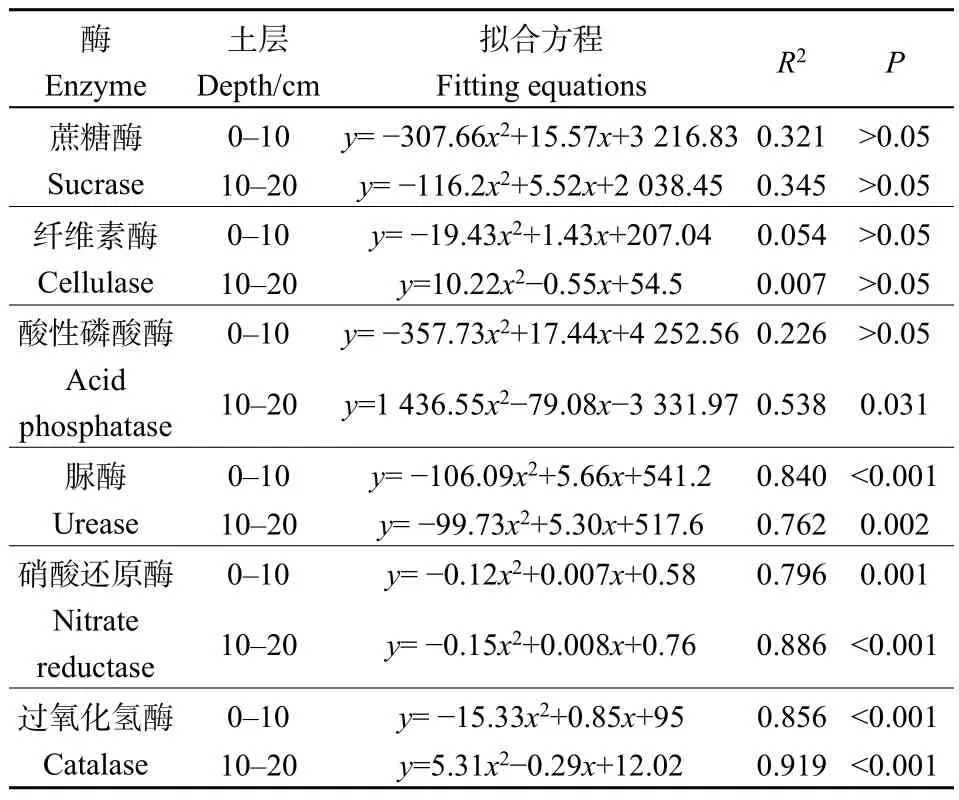
表2 土壤酶活性隨建植年限變化規律的擬合結果Table 2 Fitting results of soil enzyme activities with the change of planting years
3.4 人工草地植物群落與土壤環境因子間相關性分析
3.4.1 人工草地植物群落與土壤環境因子的 RDA排序
對人工草地植物群落特征與土壤理化指標進行了RDA排序(圖3),軸1和軸2分別解釋總變異的63.2%和15.1%,土壤AP、AK、TP和酶活性(Enzyme activities)對人工草地植物群落的影響最顯著(P<0.05)(表3)。植物生物量(Biomass)和蓋度(Coverage)方向相同,表明在群落演替過程中植物生物量和蓋度顯著相關,R/S則與凋落物(Litter)和物種豐富度正相關。不同土壤理化性質線段長度及其與植物群落指標的夾角表明,影響植物生物量的主要因素為TN、SOM、酶活性(Enzyme activities)和SWC,影響植物蓋度的主要因素為酶活性、SWC和TK,植物多樣性與土壤TN含量呈負相關。

圖3 不同建植年限土壤理化性質和植物群落特征的RDA分析Fig. 3 Redundancy analyses for soil physicochemical properties and plant community characteristics
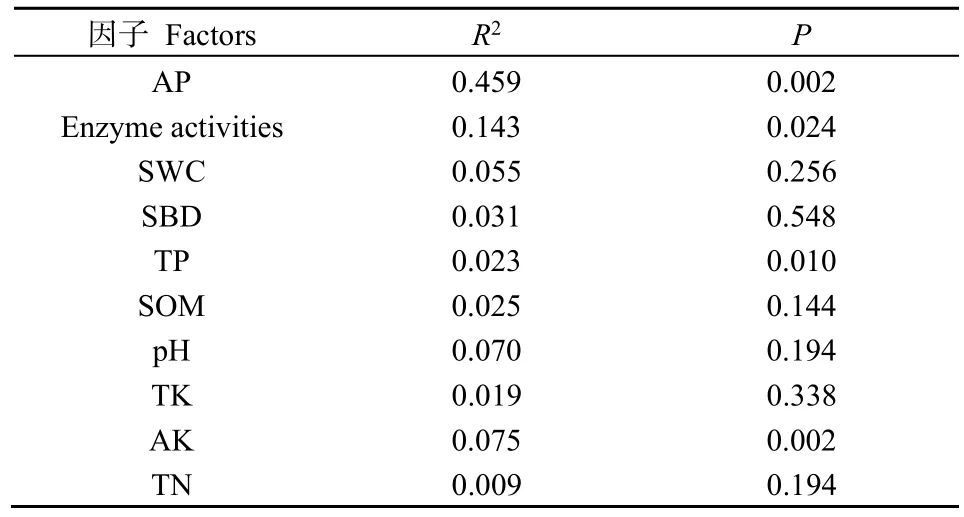
表3 冗余分析蒙特卡洛置換檢驗結果Table 3 Monte Carlo permutation test results of redundancy analysis
3.4.2 人工草地植物群落與土壤環境因子的相關性分析
植物群落特征與土壤理化性質的Pearson相關性表明(表4),植物生物量與土壤TN和SOM呈極顯著正相關(P<0.01),與 SWC呈顯著正相關(P<0.05);凋落物與土壤AP、TP和AK呈極顯著正相關(P<0.01),與R/S呈顯著正相關(P<0.05);植物多樣性與土壤pH呈顯著負相關(P<0.05)。土壤酶活性與土壤理化性質的相關性結果表明,土壤蔗糖酶活性與SOM、TN、SWC和pH呈極顯著正相關(P<0.01);土壤酸性磷酸酶活性與 SOM和 pH呈顯著正相關(P<0.05);土壤脲酶、硝酸還原酶和過氧化氫酶活性與土壤TN和SWC呈極顯著正相關(P<0.01),土壤脲酶活性與 SOM 呈極顯著正相關(P<0.01),硝酸還原酶、過氧化氫酶活性與SOM呈顯著正相關(P<0.05)。
4 討論
4.1 高寒人工草地植物群落的演替過程
時間梯度上植物群落物種多樣性的變化,在一定程度上可以反映出植被的演替變化(武勝男等,2019)。本研究中,植物多樣性在建植6年最低,而地上生物量在建植6年最高,建植9年最低。這與Dong et al.(2010)的研究結果相似,人工草地的建植會顯著降低物種豐富度和多樣性,但是能改善植被地上生物量。草地生態系統中氮素是限制植物生長最主要的因子(Mooney et al.,1987),氮素的增加會促進少數植物的生長,減少植物多樣性,同時增加地上生物量(Wedin et al.,1996;Gough et al.,2000;Stevens et al.,2004)。本研究中土壤TN在建植5—6年內顯著增加,提高了植物生物量。而建植9年,土壤養分顯著降低,養分資源的限制導致植株個體趨向于變小(Geng et al.,2007),根據最優分配模型,植物物種將按照一定比例向根系分配更多的資源,以提高其對有限土壤養分的吸收效率(Gedroc et al.,1996)。因此,建植9年人工草地植物地上生物量和土壤養分均顯著降低,但地下根系量增加。這可能是植物由有性繁殖向無性繁殖轉變的表現,建植9年后,因為無性繁殖逐漸占主導,植物群落自下而上逐漸恢復,植物生物量和豐富度增加。且增加的根系提高了土壤的保水能力和養分吸收效率(樊小林等,1995),進一步促進了地上植物的生長,在建植 13年植物豐富度、多樣性和生物量顯著升高。

表4 植物群落特征和酶活性與土壤理化性質間的相關性Table 4 Correlation between plant community characteristics, enzyme activities and soil physicochemical property
4.2 不同建植期土壤理化性質的變化
人工草地的建植對土壤質量具有顯著的改善作用,能加速碳氮循環和土壤養分的恢復過程(Ren et al.,2007)。本研究發現,土壤養分、SWC在建植6年顯著增加,和Wu et al.(2010)的研究結果一致。在人工草地恢復演替初期,地上植物生物量的增加和凋落物的積累向土壤輸入大量養分(Dong et al.,2010),同時植被蓋度和根系的增加,減少了土壤水分蒸發(歐延升等,2019),土壤保水能力提高,SWC增加,SBD降低。隨著恢復演替的進行(6—9年),由于放牧壓力的增加,畜蹄的踩踏會增加土壤緊實度,從而阻礙地表水分和養分下滲(Zhao et al.,2009),歸還土壤的植物凋落物減少,土壤水分和養分含量顯著降低。家畜的啃食會影響地上植被的分布特征及生物量,從而間接影響土壤養分的分布(常晶晶等,2018),使土壤養分在建植9年顯著降低,植物地上部分生長受阻,根系分配養分隨之增加,植物根土比逐漸升高。建植9年后,由于放牧干擾導致優質牧草比例下降,毒雜草增加,從而減少了家畜的啃食(王長庭等,2009),植物開始自我調節和恢復,土壤結構和土壤肥力得以改善。與地上植物群落動態變化特征相似,土壤養分和含水量隨人工草地恢復演替同樣呈現“N”字型變化規律,但由于土壤容重不變,根土比呈現增加趨勢,人工草地仍然處于正向恢復演替。
4.3 不同建植期土壤酶活性的變化
土壤酶類是土壤中具有生物活性的蛋白質,主要來自于土壤微生物釋放、植物根系的分泌以及土壤動植物殘體腐解過程的釋放(王啟蘭等,2010;范珍珍等,2018),是反映生態系統養分循環的重要指標(Acosta et al.,2018)。土壤酶活性的變化可以用來指示高寒草甸的演替階段和退化程度(胡雷等,2014)。本研究中,6種土壤酶活性隨人工草地恢復演替均呈現“N”字型變化規律,表現為建植9年的土壤酶活性顯著低于建植5、6和13年,這是由于:(1)建植 9年,地上生物量和凋落物生物量最低,輸入的土壤養分,尤其是SOM含量顯著降低導致酶作用底物的減少,抑制了土壤酶的活性(Acosta et al.,2007;林玉等,2019);(2)土壤含水量在建植9年也達到最低,在一定程度上降低了土壤動物和微生物活性,從而對土壤酶活性產生抑制作用;(3)土壤酶活性與土壤 pH顯著相關,如土壤酸性磷酸酶受土壤pH值的影響較大(張焱華等,2007),土壤 pH越接近中性,酸性磷酸酶活性越高(黃宇等,2008)。在建植 6—9年,土壤pH從7.34降到6.25,導致0—10 cm土層土壤酸性磷酸酶活性降低。土壤深度對酶活性影響也顯著,酶活性(除硝酸還原酶活性)隨土層加深而降低,與多數研究結果一致(文都日樂等,2010;Ananbeh et al.,2019)。一方面,植物0—10 cm土層的根系分布顯著高于10—20 cm土層,根系的分泌物是微生物豐富的能源物質(王長庭等,2009)。其次,土壤表層的凋落物和腐殖質累積較多,有機質含量高,土壤有機質作為酶合成的前體,能增加土壤中的酶源(如:土壤微生物生物量),在酶物理穩定性中具有重要作用(Acosta et al.,2007)。隨著土層加深,根系分布、土壤有機質含量和微生物數量及活性的降低是導致酶活性降低的重要原因。
4.4 植物群落特征與土壤環境的協同關系
人工草地恢復演替過程中,地上植物群落、土壤理化性質和土壤酶活性均呈現相似的“N”字型動態變化特征,證明了地上植物群落和土壤環境因子之間具有協同效應,且與草地恢復演替存在明顯的關聯性。人工草地建植初期的施肥等人為干預措施,增加了土壤養分供給,充足的養分資源促進了植物群落的生長(Chapin et al.,1987),特別是土壤氮素的增加顯著提高了植物生物量,間接促進了植被蓋度和后期地表凋落物的增加,從而提高了SWC和土壤養分的有效性。隨著演替的進行,由于管理不當(過牧),土壤肥力和SWC下降,土壤pH顯著增加,從而抑制了生物群體的養分供應能力(Brady et al.,2002),建植7—9年植物地上生物量和豐富度顯著降低。同時,土壤養分的限制改變了植物對養分資源的分配模式(Gedroc et al.,1996),加大了對根系的投資力度。且由于植物群落中各物種對于環境變化響應的對策不同,群落優勢種發生改變,莎草科等通過根系分蘗進行無性繁殖的物種取代禾本科成為草地優勢種。建植10—13年,人工草地正向演替的主導因素由土壤養分逐漸轉變為植物根系。地下根系的增加,有利于保持土壤水分,提高根系對養分的吸收效率,并增加土壤酶源。土壤酶類通過參與碳氮磷等的循環增加了植物多樣性(胡雷等,2014),保證了植物光合作用的進行和根系的發育(戴凌,2014;De et al.,2020),土壤-植物復合系統的功能得以恢復。
5 結論
(1)建植 5—6年,土壤理化性質與植物生物量、蓋度及凋落物量等植物群落特征的協同演替促進了人工草地的恢復,土壤養分和水分是此階段恢復演替的主導因子。
(2)建植 7—9年,草地土壤養分、酶活性、植物生產力和豐富度顯著降低,但由于植物根系不斷增加,人工草地仍處于正向演替。
(3)建植 10—13年,人工草地正向演替的主導因素由土壤養分轉變為植物根系,草地也由有性繁殖主導轉變為無性繁殖主導,開始自下而上的恢復,植物地上生物量和豐富度隨之增加,草地向正向演替發展。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
