棕囊藻屬(Phaeocystis)的種類多樣性及地理 分布特征研究進展*
2021-02-03 01:43:34沈萍萍齊雨藻
海洋與湖沼 2021年1期
沈萍萍 齊雨藻
(1. 煙臺大學海洋學院 煙臺 264005; 2. 暨南大學赤潮與海洋生物學研究中心 廣州 510632; 3. 水體富營養化與赤潮防治廣東普通高校重點實驗室 廣州 510632)
棕囊藻(Phaeocystis)是全球海洋廣泛分布的有害藻華原因種, 也是海洋初級生產力的重要貢獻者, 在極地和近海地區的碳、硫元素的生物地球化學循環、食物網結構及全球氣候變化中都具有極其重要的作用(Verity et al, 2007)。多數棕囊藻具有復雜的異型生活史(heteromorphic life cycle, 亦稱多態生活史polymorphic life cycle), 兼有單細胞和囊體兩種形態(Rousseau et al, 2007)。在富營養條件下, 棕囊藻能在短時間內暴發性增殖形成藻華, 不僅能夠產生溶血性毒素、二甲基硫化物(DMSP/DMS)及硫丙酸等化合物影響海洋生物(van Rijssel et al, 2007), 囊體衰亡后還形成大量泡沫物質, 嚴重影響海洋生態系統的結構與功能, 給漁業養殖造成巨大危害與損失(齊雨藻等, 2002; Schoemann et al, 2005)。棕囊藻藻華因其暴發規模大、持續時間長、危害嚴重, 一直受到高度關注, 是國際、國內有害藻類研究的熱點之一(Qi et al, 2004; 沈萍萍等, 2018)。
1997 年10 月末中國東南沿海首次暴發了特大規模的棕囊藻藻華, 其原因種為我國一種藻華新記錄種——球形棕囊藻(Phaeocystis globosa)(陳菊芳等, 1999; 陳月琴等, 2002)。此后, 陸續在廣東、福建、海南、廣西北部灣等海域都暴發了此種藻華。2004年渤海天津海域也首次錄得球形棕囊藻藻華的發生(曲凌云等, 2008)。截至2017 年底, 中國沿海已報道的棕囊藻藻華高達55 起, 發生面積超過14000 km2(沈萍萍等, 2018)。此種藻華能致養殖漁業遭受巨大損失, 如僅1997 年底廣東拓林灣的網箱養殖損失達9000 余萬(陳菊芳等, 1999)。從2014 年開始, 廣西北部灣海域大量棕囊藻囊體堵塞濱海核電站的冷源取水口, 嚴重影響核電運行安全(曹西華等, 2017), 引起社會極大關注, 也為棕囊藻藻華的研究提出了新的挑戰與要求。
近二十年來我國學者針對棕囊藻進行了諸多研究(沈萍萍等, 2018), 尤其是最近幾年取得了顯著的進展(Wang et al, 2015; 梁大勇等, 2018; 胡曉坤等,2019; 胡章喜等, 2019; Zhang et al, 2020)。但是有關中國沿海棕囊藻形態及分類的研究卻一直較少(陳月琴等, 2002; 覃仙玲等, 2016; 胡章喜等, 2019), 目前報道的僅“球形棕囊藻” 1 種, 絕大多數報道主要依據藻華暴發時的“球形囊體”特征將其認定為“球形棕囊藻”, 但不同地理株出現了明顯的分化(Shen et al,2011; Xu et al, 2017; 胡曉坤等, 2019), 有可能存在隱存種現象。此外, 過去我們關注到東海海域一直沒有球形棕囊藻及藻華的研究報道, 實際上卻有2 種新記錄種存在(即心形棕囊藻Phaeocystis cordata Zingone et Chrétiennot-Dinet 和楊棕囊藻P. jahnii Zingone), 且豐度較高有形成藻華的跡象(Lin et al, 2014)。這一現象表明, 中國沿海有關棕囊藻種類多樣性的研究比較匱乏, 而種類的分類與鑒定是所有研究的基礎和根本, 亟需填補。因此本文就全球范圍內棕囊藻屬的分類及地理分布特征等研究進展作一梳理與總結,為厘清中國沿海棕囊藻的種類及遺傳多樣性現狀提供基礎資料。
1 棕囊藻的分類及形態特征
棕囊藻屬(Phaeocystis)由Lagerheim 始建于1893年, 現隸屬于定鞭藻門(Haptophyta)定鞭藻綱(Haptophyceae)棕囊藻目(Phaeocystales)(見Algaebase條目), 是廣溫廣鹽性的海洋微藻, 廣泛分布于從極地到熱帶地區各大海洋中(Baumann et al, 1994)。此屬的主要特征是既有單細胞型又有群體型; 運動細胞< 10 μm 且具2 條長鞭毛及1 條短直的定鞭毛; 有些種類具不動細胞階段, 細胞單生或聚集成球形或不規則的囊體。由于棕囊藻細胞個體微小且多具有復雜的異型生活史, 具有單細胞和囊體兩種形態, 因此棕囊藻屬種類的確定從一開始即頗多爭論(齊雨藻等,2001)。1892 年, Pouchet 首次報道了1882 年夏季發現于挪威海域的膠質囊狀“棕囊藻”, 由于與綠藻四孢藻屬(Tetraspora)的Tetraspora giraudyi Derbès & Solier(1851)的膠質囊很像而被命名為Tetraspora poucheti Hariot (Pouchet, 1892)。1893 年Lagerheim 在瑞典Koster 群島海域發現相似種類并質疑這兩個海洋棕色藻類不該屬于淡水綠藻四孢藻屬, 他因此建立新屬棕囊藻屬(Phaeocystis Lagerheim), 將 Tetraspora poucheti Hariot 重新命名為波切棕囊藻[Phaeocystis pouchetii (Hariot) Lagerheim]并以此為棕囊藻屬模式種。原屬四孢藻屬的淡水綠藻T. fuscescens Braun ex Kützing 與T. giraudyi (Derbès & Solier)也被相繼歸于棕囊藻屬, 即 P. fuscescens (Braun) De Toni 與 P.giraudyi (Derbès and Solier) Hamel。
隨后, 球形棕囊藻(Phaeocystis globosa Scherffel,1899)和南極棕囊藻(P. antarctica Karsten, 1905)分別在德國北海及南極海域被發現, Scherffel (1900)在球形棕囊藻中首次詳細描述了棕囊藻的第三根鞭毛“定鞭體(haptonema)”。1911 年, Büttner 從德國Kiel 港又分離鑒定 2 個種 P. sphaeroides Büttner 和 P.amoeboidea Büttner, 這2 個種具有肉眼可見的棕色囊體, 細胞內具1 個黃棕色葉綠體且運動細胞具2 條等長的鞭毛, 卻因細胞內僅有1 個葉綠體而受到質疑(Medlin et al, 2007)。1922 年, 在南極洲又發現一新種Phaeocystis brucei Mangin, 細胞大小為5—6 μm, 每個細胞含有2 個金黃色葉綠體, 囊體直徑1—2 mm,此種囊體與球形棕囊藻和南極棕囊藻的囊體具有明顯的差別在于存在一些與囊體通過黏液小徑相連的獨立細胞簇。早期由于受光學顯微觀察技術的限制,很難獲得單細胞的詳細形態結構, 因此主要基于囊體形態與地理分布差異將不同棕囊藻種分開。1955年Kornmann (1955)詳細比較了球形棕囊藻和波切棕囊藻的囊體形成過程, 發現兩個種的囊體發展過程很相似, 大約在16 細胞時期才出現不同, 因此他認為球形棕囊藻是波切棕囊藻的幼體形式, 囊體形態特征從此后成為一個不太可靠的分類標準。
時隔多年直到1979 年, Moestrup (1979)綜合利用電子掃描及透射顯微鏡在新西蘭海域發現了只有單細胞形態的新種P. scrobiculata Moestrup (蜂窩狀棕囊藻), 至此已被描述的棕囊藻共有9 種。Sournia (1988)將從建屬以來發表過的這9 種棕囊藻重新審訂, 發現大多數種的鑒定不充分, 囊體特征易發生變異, 而單細胞缺乏必要的亞顯微結構特征描述如鞭毛長度、體表鱗片、附屬物等特征, 他剔除了明顯不符合棕囊藻屬特征的兩個淡水種P. fuscescens 與P. giraudyi, 還有3 個種如P. amoeboidea、P. brucei 及P. sphaeroides等種類自首次描述過后一直沒有獲得純培養而無法深入鑒定, 而對于兩個常見種類P. pouchetii 和 P.globosa 進行細致研究后卻沒有將二者區分開, 因此Sournia 認為確定的只有兩個種: 一是僅發現單細胞形態的蜂窩狀棕囊藻(P. scrobiculata Moestrup), 二是能夠形成囊體的波切棕囊藻[P. pouchetii (Hariot)Lagerheim, 包括球形棕囊藻在內]。為避免混淆, 1990年代中期前大多數海洋生態學家遵從Sournia 的“兩大類”建議來報道棕囊藻。有學者認為這一分類過于簡化, 如Baumann 等(1994)依據地理分布、溫度和光的耐性、囊體形態等特征將棕囊藻細分為4 個不同的種: 即蜂窩狀棕囊藻、球形棕囊藻、波切棕囊藻及南極棕囊藻。
其后隨著觀察技術的發展和研究的深入, 棕囊藻的分類特征有了新的補充, 運動細胞的特征如細胞大小、鞭毛及定鞭體的長短、星形線狀分泌物以及體表鱗片等成為重要的分類標準(Parke et al, 1971;Zingone et al, 1999; Peperzak et al, 2000; Jacobsen,2002)。特別是星形線物質特征, 即運動細胞產生并釋放一種纖維狀的幾丁類物質, 能夠形成五角星形或九角星形的模式(圖1), 這個特征成為棕囊藻屬一個重要且特有的分類標準(Chrétiennot-Dinet et al, 1997),用來鑒定不同棕囊藻運動單細胞的種類, 如波切棕囊藻、球形棕囊藻及南極棕囊藻(P. antarctica Karsten)細胞分泌物質的特征為五角星形, 而蜂窩狀棕囊藻的分泌物質特征卻是九角星形。
后來隨著分子標記技術的應用, Medlin 等(1994)利用18S rRNA 基因序列對上述分類結果進行了遺傳水平的確定, 首次從分子水平上將球形棕囊藻與波切棕囊藻兩個種區分開。1999 年Zingone 等(1999)又根據傳統形態和核糖體SSU rRNA 分子特征分析在地中海海域發現2 個新種, 即P. cordata (心形棕囊藻)和P. jahnii (揚棕囊藻)。基于形態學及分子生物學分析, Medlin 等(2007)認為已有的對棕嚢藻的形態學分類研究不足以界定本屬, 應予修正。為此, 她們對棕嚢藻屬做了如下界定:
棕嚢藻(Phaeocystis Lagerheim 1893, Medlin &Zingone emended)
運動細胞具兩條等長或不等長鞭毛及一條短直定鞭體; 葉綠體1—4 個側生; 細胞常為2 種不同大小的平滑鱗片覆蓋。某些種類具有射出的細胞器。生活史復雜, 包括形成不動細胞的階段, 但此種狀況不是所有種類皆如此。不動細胞通常無附屬物及鱗片, 細胞單生或排列成球形、有裂瓣的、具色鞘或不規則的膠質囊體; 如有附屬物, 通常短而不完整。
最近, Andersen 等(2015)從阿拉伯海域發現了1種新的單細胞種類——冠狀棕囊藻(Phaeocystis rex),此種僅觀察到單細胞形態存在, 其細胞個體較大, 直徑6—10 μm, 長8—15 μm, 尤其外被的多層不定形有機鱗片和指狀突起的葉綠體特征, 較其他棕囊藻種有顯著的差別, 與Medlin 等(2007)所界定的棕囊藻屬的基本特征亦不完全吻合, 但系統進化分析結果卻顯示冠狀棕囊藻是介于單細胞種類和群體型種類的中間進化類型。

圖1 棕囊藻的星形線分泌物特征Fig.1 The star-like filaments of Phaeocystis
至此已被描述的棕囊藻有 10 個種, 分別是:Phaeocystis amoeboidea Büttner 變形蟲狀棕囊藻、P.antarctica Karsten 南極棕囊藻、P. brucei Mangin 布魯斯棕囊藻、P. cordata Zingone et Chrétiennot-Dinet 心形棕囊藻、P. globosa Scherffel 球形棕囊藻、P. jahnii Zingone 揚棕嚢藻、P. pouchetii (Hariot) Lagerheim 波切棕囊藻、P. rex Andersen, Bailey, Decelle & Probert冠狀棕囊藻、P. scrobiculata Moestrup 蜂窩狀棕囊藻、P. sphaeroides Büttner 圓形棕囊藻等。其中7 個種已經過DNA 分子鑒定或電鏡形態特征檢驗, 而剩余P.amoeboidea (1910) 、 P. sphaeroides (1911) 及 P.brucei (1922) 3 個種類由于在原文無明確的附圖及可疑性狀(如只有1 個葉綠體及無定鞭毛), 在此后的文獻中再也沒有被論及, 故本文按照“棕囊藻屬種的檢索表”介紹7 種確定棕嚢藻的綜合形態特征(Medlin et al, 2007; Rousseau et al, 2007)(圖2, 3, 4):

棕嚢藻屬種的檢索表
(一) 群體型:
(1) 波切棕嚢藻
Phaeocystis pouchetii (Hariot in Pouchet)Lagerheim (Pouchet, 1892) (模式種)(圖2a, 3a)
囊體呈裂瓣狀, 囊體內細胞往往4 個聚堆。具鞭毛階段的細胞為球形, 平均直徑5 μm, 2 條鞭毛等長約11 μm, 定鞭體很短1—2 μm。細胞外有2 類鱗片:一種為環片狀0.24 μm×0.25 μm; 另一種為卵圓形且邊緣微卷曲的鱗片0.9 μm×0.15 μm, 覆蓋于大鱗片之下層。細胞核后位, 葉綠體2 塊且內生蛋白核, 高爾基體居二者其間。
(2) 球形棕嚢藻
Phaeocystis globosa Scherffel (Scherffel, 1900)(圖2b, 3b, 3c)
囊體呈球形, 囊體內細胞均勻分布。具鞭毛階段的細胞大小3—6 μm。2 條鞭毛等長, 長度為體長1.5倍。定鞭體的長度是鞭毛的1/4 至1/3。體表鱗片有2類: 環片狀(0.18 μm×0.19 μm)及小形底墊的卵圓狀鱗片(0.10 μm×0.13 μm)。具2 片葉綠體, 細胞核后位, 具高爾基體及多數簇生潴泡(cisternae)。
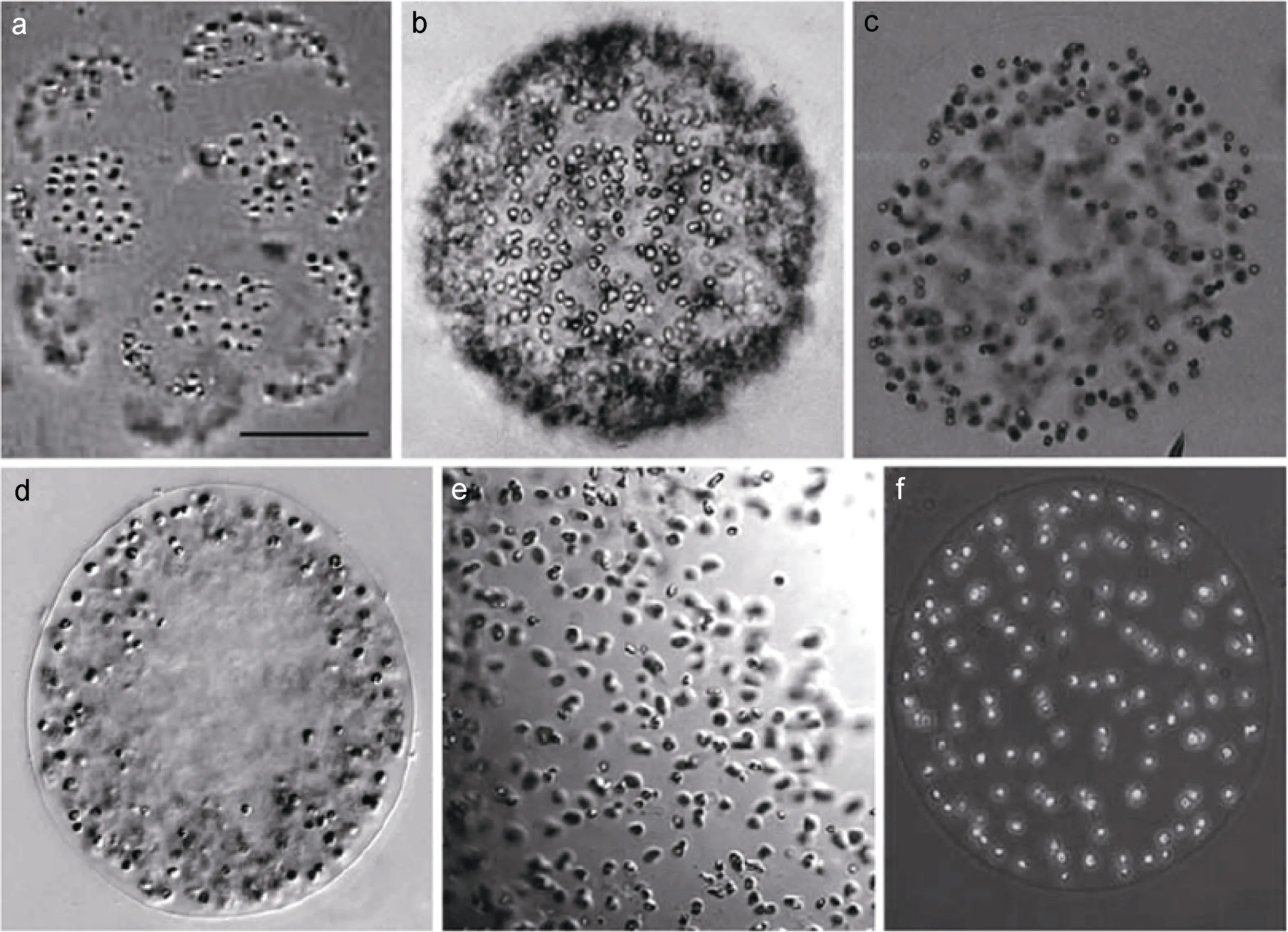
圖2 群體型棕囊藻囊體的光學形態特征(引自Medlin et al, 2007)Fig.2 Light microscopic micrographs of colony stages of Phaeocystis (from Medlin et al, 2007)
(3) 南極棕囊藻
Phaeocystis antarctica Karsten (1905)(圖2c, 2d, 3d)
囊體為圓形囊狀, 細胞散布在近囊體表層, 囊體會隨年齡增長而變成扭曲狀或伸長。囊體內細胞呈圓形或稍近四邊形, 直徑4—6 μm, 通常具有2 個或罕見有4 個拉長的金棕色葉綠體, 每個含1 個紡錘狀蛋白核, 高爾基體位于兩個葉綠體之間。具鞭毛階段細胞大小及形狀變化較多, 小的圓形細胞大小約4 μm長, 而大的卵圓形或梨形細胞長約6 μm; 具2 條明顯不等長且尖頭的鞭毛(分別長7—12 μm 及12—18 μm),定鞭體長3—4.5 μm 且末段未見凸脹。大部分時間鞭毛細胞表面無鱗片覆蓋, 并且無觀察到星形線分泌物。若有鱗片覆蓋, 為圓形(直徑0.20 μm)(Zingone et al, 2011); 或大小不同的卵圓形分別為 0.27 μm×0.19 μm 及0.18 μm×0.14 μm。有鱗片覆蓋的細胞同時能形成五角星形線結構(中心0.8 μm 寬)。
(4) 揚棕囊藻
Phaeocystis jahnii Zingone (Zingone et al, 1999)(圖2e, 3e)
囊體為不定形囊狀, 囊體內細胞疏松排布(亦有認為此種屬單細胞類型), 大小約為 6—8.5 μm, 具2—4 個葉綠體。鞭毛細胞近圓形, 直徑3.5—5 μm, 具有2 條不等長的鞭毛(長8.5—12 μm 及5.5—6.5 μm),鞭毛末端凸脹不明顯。外覆細胞鱗片, 其表面上的放射紋很不明顯。藻體呈桔黃色。
(二) 單細胞型:
(5) 心形棕囊藻
Phaeocystis cordata Zingone &Chrétiennot-Dinet (Zingone et al, 1999)(圖3h, 3i, 3j)
單細胞三角形、心形或橢圓形, 間或扁平, 末端微尖, 細胞大小平均為3—3.5 μm 長, 3—4 μm 寬, 厚度約為2.5 μm。具2 條稍不等長鞭毛, 其長度分別為5.5—7.5 μm 及 4.5—6 μm, 定 鞭 體 很 短 長 度 為2.2—2.5 μm。外覆鱗片卵圓形, 大鱗片 0.25 μm×0.18 μm, 小鱗片為0.18 μm×0.13 μm。
(6) 蜂窩狀棕囊藻
Phaeocystis scrobiculata Moestrup (Moestrup,1979)(圖4a, 4b)
藻體單細胞直徑約為8 μm。具2 種鱗片, 大小分別為0.6 μm×0.45 μm 及0.19 μm×0.21 μm, 2 根鞭毛及1 根定鞭體都較球形棕囊藻的長一倍。
本種首先在新西蘭發現, 目前已在多個海域有分布記錄。
(7) 冠狀棕囊藻
Phaeocystis rex Andersen, Bailey, Decelle &Probert (Andersen et al, 2015)(圖4c, 4d)
藻體單細胞, 直徑6—10 μm, 長8—15 μm, 外周由多層不定形的有機磷片包被。細胞末端有時有疣狀突起, 細胞具有2 個葉綠體, 每個葉綠體呈前伸手指狀突起, 內有1 蛋白核。不動細胞及運動細胞都具有2條較短的鞭毛及1 條定鞭體。模式樣本來自阿拉伯海。
(三) 未確定種類及形態(Undescribed species/morphotype)
近年來有更多的棕囊藻新種或新形態被發現(Medlin et al, 2007; Doan-Nhu et al, 2010a; Rousseau et al, 2013; Novarino, 2014; Andersen et al, 2015)。Medlin 等(2007)報告了地中海海域分離的3 個未確定種: Phaeocystis sp.1 (PML559)(圖5), Phaeocystis sp.2(圖2f), Phaeocystis sp.3 (圖3g)。其中Phaeocystis sp.1和Phaeocystis sp.3 為單細胞種類, 而Phaeocystis sp.2能夠形成囊體, 其運動細胞形狀不規則, 2 條鞭毛明顯不等長且定鞭體無凸脹末端; 細胞表面無鱗片及纖維(圖2f); 雖然18S rDNA 序列分析表明其屬于球形棕囊藻分支, 但是ITS rDNA 序列分析卻無法與其他球形棕囊藻株系聚類。Doan-Nhu 等(2010a)曾描述過越南沿海及南海廣泛分布的一種棕囊藻單細胞形態(Phaeocystis spp.), 細胞近圓形, 大小約3 μm, 2 條鞭毛可能等長, 定鞭體不清楚, 能夠形成五角星形結構, 具有2 種大小不同的圓形平坦的鱗片且與已知棕囊藻種類皆不同, 但由于原文及照片描述比較模糊無法進一步確定。
Novarino(2014)報道了地中海西部海域6 種棕囊藻單細胞形態(Morphotype 1—6), 其中Morphotype 1—3 分別對應了波切棕囊藻、球形棕囊藻及蜂窩狀棕囊藻的運動細胞形態, 而其余3 種(Morphotype 4—6)與已知的棕囊藻種類皆不相符(圖6—8)。其中Morphotype 4 為此海域中繼Morphotype 2(即球形棕囊藻)之后的第二高豐度的細胞形態(圖6), 其細胞大小為3—3.5 μm, 具不等長鞭毛, 沒有定鞭體。基于鞭毛的長度, 此類細胞分為兩種類型, 類型1(Form-1)細胞鞭毛分別長為7—8 μm 和5—6 μm 并能觀察到五角星形線(圖6a), 類型2(Form-2)細胞鞭毛長15—20 μm和10—12 μm 但未觀察到星形線結構(圖6e); 細胞表面覆蓋2 種鱗片, 大的橢圓形為0.44 μm×0.31 μm 和小的圓形鱗片平均直徑為0.13 μm (圖6b—6d), 此種最有可能為棕囊藻新物種(Novarino, 2014)。Morphotype 5 在地中海的分布及數量稀少, 大多數為3—4 μm細胞, 具有 2 條不等長鞭毛分別為 20—30 μm 和15—22 μm 長(圖7)。五角星形線結構中間部分無孔,被纖維物質緊密圍繞。細胞表面的橢圓形鱗片非常類似波切棕囊藻, 但是大小有較大差別。此形態與波切棕囊藻經常同時出現, 因此推測很可能是波切棕囊藻的某個生活史階段(Novarino, 2014)。Morphotype 6僅觀察到一個樣品, 其直徑約3 μm, 2 條不等長鞭毛分別長5 和3.5 μm(圖8)。細胞被10 個未完全展開的纖維絲包所包圍, 未見五角星形或九角星形結構(如果這些纖維絲能夠完全展開, 可能會形成類似五角星線結構)。外覆鱗片只有1 種類型大小約0.60 μm×0.51 μm, 鱗片大小及外形類似蜂窩狀棕囊藻, 但沒有觀察到其特征性的九角星形線形結構。目前一個共性問題是尚不能確定這些細胞型能否代表生物學上獨立的棕囊藻種類, 還是同一個種不同生活史階段的細胞形態, 需要進一步結合分子遺傳信息、生活史研究及傳統形態學觀察來綜合確定。
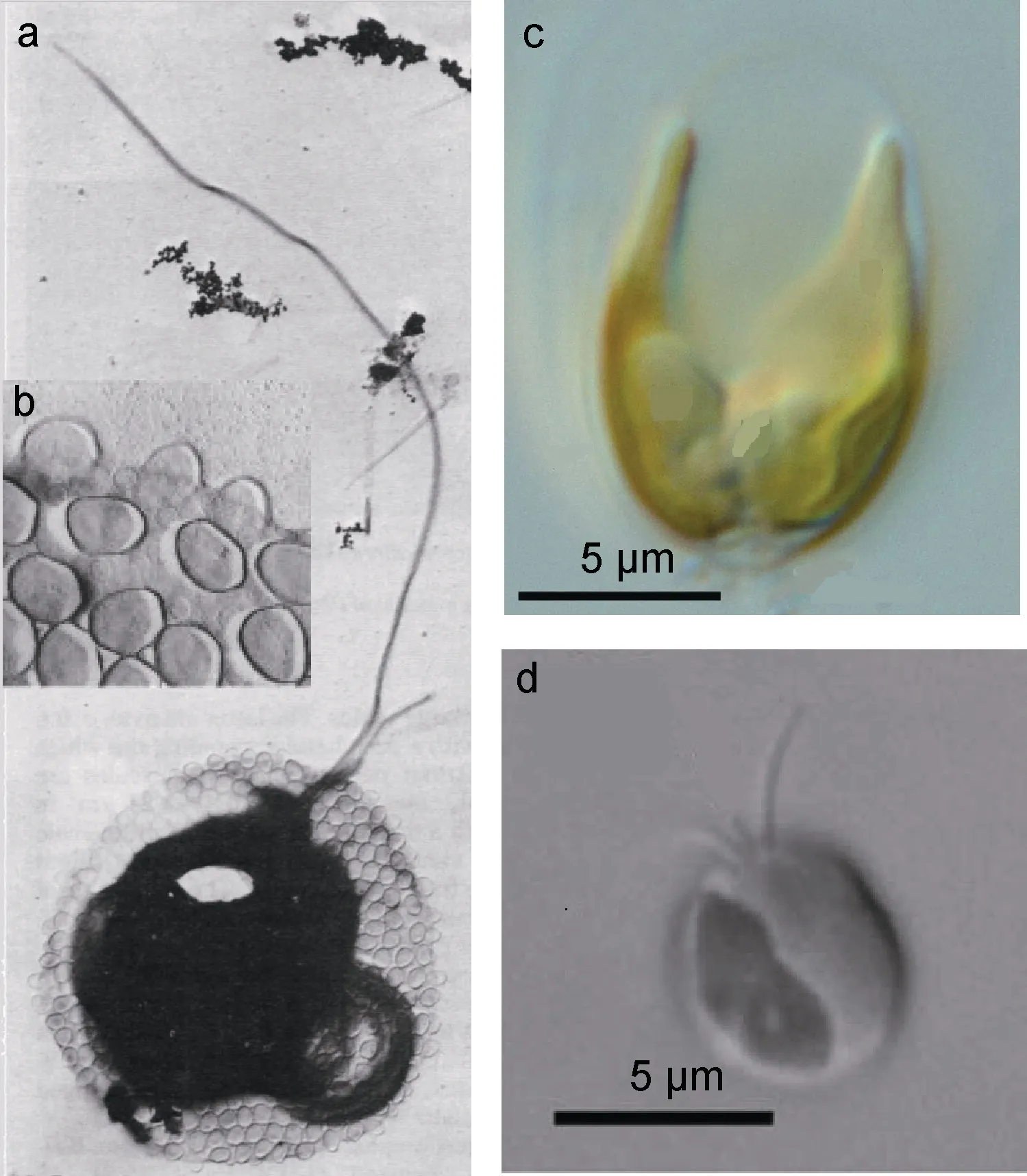
圖4 單細胞型棕囊藻(蜂窩狀棕囊藻和冠狀棕囊藻)的細胞形態特征(引自Moestrup, 1979; Andersen et al, 2015)Fig.4 Microscopy of solitary Phaeocystis (from Moestrup, 1979;Andersen et al, 2015)
雖然未確定的棕囊藻種類或形態較多, 僅從地中海海域就描述了6 種未定種(株)(不包括已確定的5種: 波切棕囊藻、球形棕囊藻、蜂窩狀棕囊藻、心形棕囊藻及楊棕囊藻), 但是它們是否屬于表型、生理、基因抑或生態的多態性還不能確定。棕囊藻復雜的生活史是理解問題的關鍵所在。在所有棕囊藻種類中,有7 種已確定具有多個形態迥異的生活史階段, 除了能夠形成囊體的波切棕囊藻、南極棕囊藻和球形棕囊藻, 還包括楊棕囊藻、心型棕囊藻、蜂窩狀棕囊藻及冠狀棕囊藻(Vaulot et al, 1994; Peperzak et al, 2000;Rousseau et al, 2007; G?bler-Schwarz et al, 2013;Andersen et al, 2015)。因此考慮到種內多態性的存在,這些未確定種類及形態必須要在明確其生活史的基礎上才能得以確定。再者由于獲得信息不對稱, 如早期的種類描述中只有囊體形態而沒有細胞超微結構及鱗片等電鏡觀察特征, 后來的研究中有些種類不能分離培養而無法獲得其DNA 序列信息, 加之棕囊藻本身復雜的異型生活史, 增加了太多的不確定性,使得棕囊藻種類的鑒定變得更為錯綜復雜。
但不管怎樣, 不論從形態學還是分子遺傳信息來講, 目前棕囊藻屬的種類多樣性被低估是毋庸置疑的(Medlin et al, 2007)。越來越多的超微型真核生物研究表明, 定鞭藻類是熱帶、亞熱帶海洋生態系統中重要的初級生產者, 是2—8 μm 真核生物類群的優勢種, 但對于其種類組成還遠不了解, 大部分種類以“類群”和“未培養克隆(Uncultured clone)”的形式出現(Liu et al, 2009; Masquelier et al, 2011; Lin et al, 2014;Shih et al, 2019)。因此棕囊藻屬可能遠不止目前已描述的10 種, 一方面由于棕囊藻單細胞個體微小, 在常規顯微檢測中很容易被忽略; 另一方面由于棕囊藻具有較強的生態適應性, 它們的分布極其廣泛, 但目前調查取樣的范圍與樣品還遠遠不夠, 尤其是基因庫中大量“未培養克隆”種類的存在, 表明全球范圍內棕囊藻種類多樣性的工作還有很大的研究空白,有待于結合實驗室分離培養、形態觀察及分子鑒定等技術手段進行深入探索與發現。
2 棕囊藻的系統進化及地理分布
棕囊藻是對溫度和鹽度適應性非常廣的海洋微藻,生境多樣, 從南北極到赤道的沿海及大洋生態系統中皆有分布(Schoemann et al, 2005; Verity et al, 2007)。
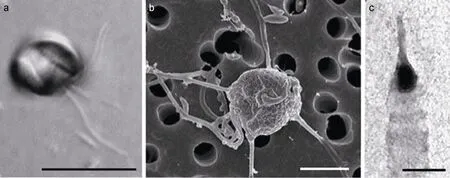
圖5 地中海海域報道的未定種棕囊藻鞭毛藻細胞特征 [Phaeocystis sp. 1 (PML 559), 引自 Medlin et al, 2007]Fig. 5 LM, TEM and SEM micrographs of flagellated stages of Phaeocystis sp. 1 (PML 559, from Medlin et al, 2007)
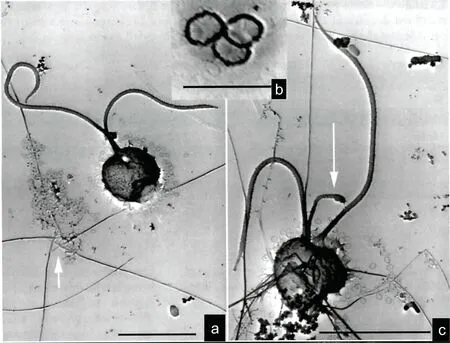
圖7 地中海海域棕囊藻鞭毛藻細胞形態5 (Morphotype 5)(引自Novarino, 2014)Fig.7 Phaeocystis morphotype 5 from Mediterranean Sea (from Novarino, 2014)
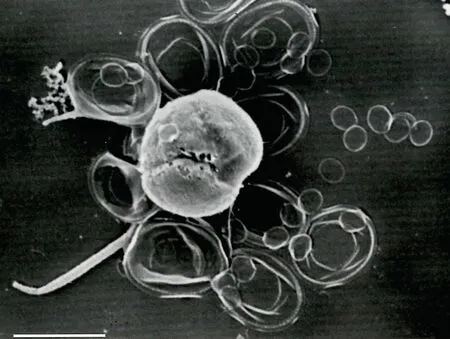
圖8 地中海海域棕囊藻鞭毛藻細胞形態6 (Morphotype 6)(引自Novarino, 2014)Fig.8 Phaeocystis morphotype 6 from Mediterranean Sea (from Novarino, 2014)
群體型棕囊藻由于能夠形成肉眼可見的囊體且能引發大規模藻華而備受關注, 其豐度極高(發生藻華時細胞豐度可達106—108cells/L), 其中經常發生藻華的種類有南極棕囊藻、波切棕囊藻及球形棕囊藻,并且具有特定的地理分布范圍: 南極棕囊藻主要分布在南極周邊海域; 波切棕囊藻分布在北半球高緯度的北冰洋、北大西洋海域; 球形棕囊藻主要分布在北大西洋、北海沿海等溫帶海域及熱帶、亞熱帶海域。這些群體型棕囊藻特定的地理分布特征具有其生理學基礎, 野外觀察和實驗室研究表明南極棕囊藻的溫度耐受范圍介于<-2—14°C, 最適生長溫度為4.5°C; 波切棕囊藻溫度耐受范圍介于<-2—14°C, 其最適生長溫度為 8°C; 而球形棕囊藻耐受范圍為<-0.6—22°C, 最適生長溫度為16°C, 后者的溫度范圍及最適生長溫度(尚未包括熱帶亞熱帶海域球形棕囊藻株系)明顯高于前兩者(Jahnke et al, 1987; Jahnke,1989; Medlin et al, 1994)。尤其是隨著海域富營養化的加劇以及全球氣候的異常, 球形棕囊藻藻華的發生范圍不斷擴大, 逐漸覆蓋熱帶、亞熱帶海域。1990年夏季在科威特灣首次發生棕囊藻藻華(Al-Hasan et al, 1990); 1996 年夏季在阿拉伯海外海又暴發了球形棕囊藻藻華(Madhupratap et al, 2000); 1997 年10 月中國東南沿海首次暴發球形棕囊藻藻華后在南海沿海迅速擴展, 其藻株耐受水溫高達30°C 以上, 最適溫度為25°C, 明顯高于北溫帶海域的球形棕囊藻株系(Shen et al, 2011; Xu et al, 2017); 越南沿海從2002—2008 年幾乎年年都暴發大規模的球形棕囊藻藻華(Doan-Nhu et al, 2010b); 南海沿海球形棕囊藻藻株由于形成的囊體巨大(直徑超過3 cm)且能耐受高溫,形成了非常獨特的球形棕囊藻分支(王小冬等, 2012;Medlin et al, 2007; Smith et al, 2014; Liu et al, 2015)。
相對于群體型種類, 單細胞種類棕囊藻發現較晚, 第一種即蜂窩狀棕囊藻于1979 年才在新西蘭海域發現(Moestrup, 1979), 但這并非表明單細胞棕囊藻的細胞豐度低, 實際上這些種類的細胞豐度亦較高, 如心形棕囊藻在地中海海域細胞豐度最高可達105cells/L (Zingone et al, 1999), 而中國東海北部海域心形棕囊藻和楊棕囊藻的混合豐度超過107cells/L(Lin et al, 2014)。由于棕囊藻單細胞個體微小, 而且常規固定方法很容易引起細胞變形, 同時又需要專門的研究工具才能準確鑒定, 因此單細胞種類的研究相對較少, 其地理分布記錄零星而分散: 如之前蜂窩狀棕囊藻只在新西蘭有記錄, 揚棕囊藻及心形棕囊藻僅在地中海中部海域發現(圖9a, Schoemann et al,2005), 新種冠狀棕囊藻僅在阿拉伯海域有記錄。但隨著研究的發展, 近年來單細胞種類的分布記錄不斷增加: 蜂窩狀棕囊藻陸續在越南沿海及南海(Doan-Nhu et al, 2010a)、澳大利亞(Hallegraeff, 1983)、熱帶大西洋(Estep et al, 1984)、北太平洋(Baumann et al, 1994)、格陵蘭島(Moestrup et al, 2003)、巴西(Bergesch et al, 2008)、地中海西部(Novarino, 2014)等海域被發現, 遍布南北半球的熱帶、溫帶及寒帶海洋, 表明此種應該是世界廣布性種類; 心型棕囊藻和楊棕囊藻在中國東海及南海海域(Lin et al, 2014; Wu et al, 2015)、斯堪的納維亞沿海(Guiry et al, 2020)、新西蘭(Guiry et al, 2020)、北太平洋、印度洋、紅海等(Decelle et al, 2012)均有報道并且豐度較高, 分布范圍涵蓋了赤道到溫帶的廣闊海域, 亦驗證了“全球廣泛分布種”的推測(Zingone et al, 1999)。
除此以外, 尚有大量的“未確定種類”存在并且遍布全球各大海洋(圖9a, Schoemann et al, 2005), 如前述地中海海域就有多種未確定種類或細胞形態, 還有很多“未培養克隆”棕囊藻種類的存在, 不僅分布極其廣泛, 而且豐度較高(Lin et al, 2014; Wu et al, 2015)。有些棕囊藻還以共生的形式存在(Decelle et al, 2012),研究表明這些棕囊藻與放射蟲共生, 分布廣泛且豐度極高, 遍布全球開闊大洋, 包括太平洋、印度洋、紅海、地中海、英吉利海峽、南大西洋和南極洲。其中在亞熱帶太平洋、熱帶印度洋及地中海海域發現的幾種未確定種棕囊藻(Phaeo1, Phaeo2), 其進化地位位于棕囊藻系統進化的最基部分支(早于心形棕囊藻和楊棕囊藻分化), 可為棕囊藻暖水起源的重要佐證(Decelle et al, 2012)。而且地中海海域出現的未確定種(Phaeo1, Phaeo2)能否與前面所描述的未確定種(Phaeocystis sp.1—sp.3)及形態(Morphotype 4—6)相對應起來尚不得而知, 后續可以通過建立藻類純培養將分子克隆信息與細胞形態特征有機結合起來進行研究。
綜上所述, 棕囊藻屬的全球分布范圍尤其是新種的分布, 需要進一步重新審視, 我們在Schoemann等(2005)的基礎上將至今發表的相關信息進行了補充與更新(圖9b; 主要參考Guiry et al, 2020)。前后對比發現有兩個特點, 一是種類分布上, 球形棕囊藻、心型棕囊藻及蜂窩狀棕囊藻的地理分布范圍有顯著的增加, 尤其是心型棕囊藻和蜂窩狀棕囊藻, 已被證實為全球廣泛分布種, 因此預計其地理分布記錄會越來越多; 二是分布范圍上顯示了 3 個熱點區域(Hotspot), 1 個位于地中海海域, 1 個在中國南海海域及其鄰近太平洋海區, 1 個位于印度洋阿拉伯海域,這幾個熱點位于低緯度熱帶、亞熱帶及其鄰近海區,種類多樣性較高, 尤其是地中海海域是浮游植物多樣性極高的地區(López, 1994), 推測極有可能還存在其他棕囊藻種或新種, 與前述的地中海海域出現多種未確定種(株)的現象相呼應。
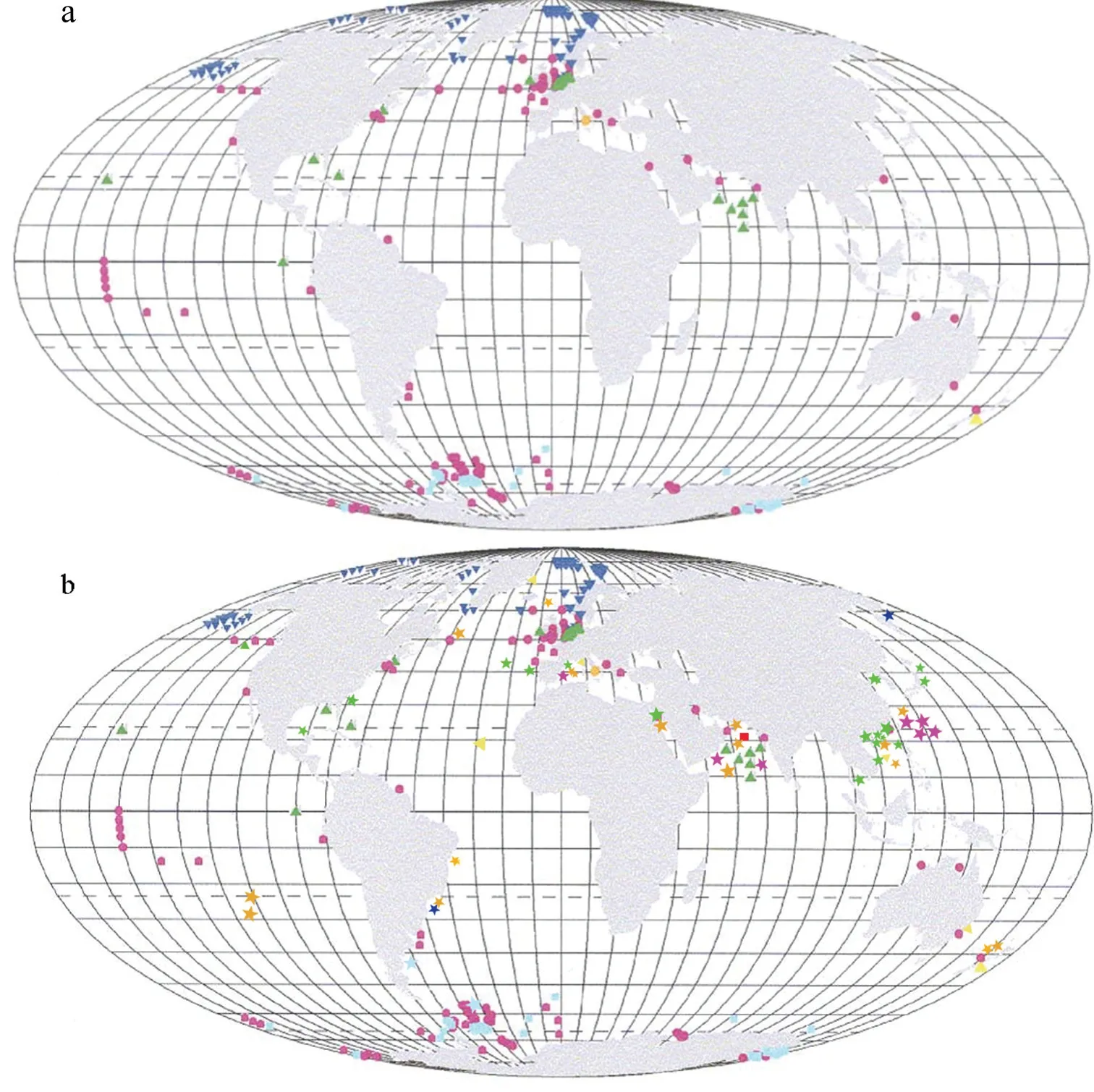
圖9 棕囊藻的地理分布比較圖(改自Schoemann et al, 2005)Fig.9 Geographical distribution of the genus Phaeocystis(modified from Schoemann et al, 2005)
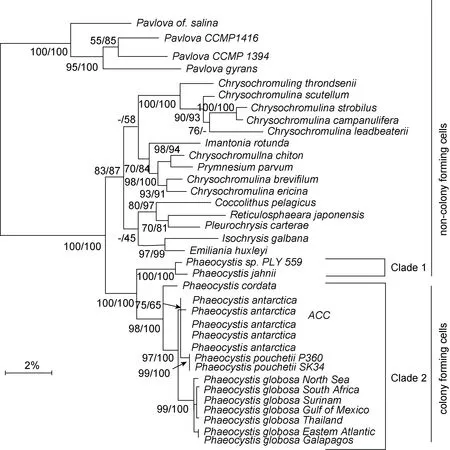
圖10 基于18S rDNA 序列構建的17 個棕囊藻種(株)和其他定鞭藻類的最大似然法系統發育樹(fastDNAml)(Medlin et al,2007)Fig.10 Maximum-likelihood phylogeny (fastDNAml) of 17 Phaeocystis species/strains and other prymnesiophytes inferred from 18S rDNA (Medlin et al, 2007)
Lange 等(2002)利用18S rDNA 及ITS1 rDNA 基因序列對全球分布的22 株棕囊藻進行了系統發育分析, 發現單細胞類型地中海楊棕囊藻(P. jahnii)和心形棕囊藻(P. cordata)位于系統進化樹的基部分支, 分化時間較早, 應該為全球廣泛分布種(圖10)。而群體型種類分化時間較晚, 又各自演化為一冷水型復合種(北極的波切棕嚢藻和南極的南極棕嚢藻)和一暖水型復合種(包含球形棕嚢藻在內)。地中海區域的楊棕囊藻和心形棕囊藻可能是約500 萬年前大西洋海水灌入干涸的地中海盆地而引入的(Hsü et al, 1977)。而南極棕囊藻和波切棕囊藻雖然分布在地球兩極, 地理分布距離最遠, 但親緣關系最近, 位于進化樹的同一分支, 由共同的祖先進化而來(圖10, Medlin et al,2007)。可以說, 前面所觀察到所有形態及生理上的變異皆具有其堅實的遺傳基礎, 證實源于不同氣候區(即北極、南極和溫帶/熱帶地區)的群體型棕囊藻的種水平上的分離。相反, ITS1 表現出大量的種內及種間遺傳分化, 除南極棕囊藻外, 其他棕囊藻種不同地理株ITS1 存在多重拷貝, 尤其球形棕囊藻可能是包括至少三個種(亞種)在內的復合種(Medlin et al, 2007),這也可以解釋為什么球形棕囊藻的分布范圍能夠跨越不同的氣候區(溫帶、亞熱帶、熱帶等)而南極棕囊藻只局限于南極附近海域。
Medlin 等(1994)以rRNA 基因為分子指標同時結合Ochman (1987) rRNA 分子鐘概念對定鞭藻類進行了研究分析, 提出棕囊藻為一種暖水起源的全球性分布藻類, 隨著地球氣候變化逐漸分化出冷水種。之后Medlin 等(1997)又根據球石藻化石記錄對定鞭藻綱分子鐘進行了校正, 將棕囊藻起源估算至7500萬年前。隨后全球氣溫驟降, 出現不同溫度水團, 冷水種得以分化。根據校正后的定鞭藻分子鐘推斷冷水種從暖水種中分化時間約在6000 萬年前, 南極德雷克海峽的形成使大西洋和太平洋得以連通從而形成環繞南極大陸的南極環流(ACC), 將南極大陸沿岸水團與北部水團有效分隔, 棕囊藻逐漸被隔離分化出一支南極冷水種(Medlin et al, 2007)。波切棕囊藻和南極棕囊藻雖然分布于地球兩極, 但卻是親緣關系最近的姊妹株, 南極海域這一冷水種即是它們的祖先(Medlin et al, 1994)。這表明擴散不是從現今的暖水種向兩極擴散演化成現今的冷水種, 而是跨越了熱帶地區的兩極之間的基因流。研究發現這2 種冷水種的進一步分化約在2000—1500 萬年前第三紀的中新紀,全球海洋有一次短暫的變暖事件(Crame, 1993)。而在此之前, 全球水溫相對較低, 允許南極海域的冷水種棕囊藻存活并越過赤道區域到達北半球。但是這次短暫變暖事件使地球升溫, 將南北兩極的棕囊藻隔離分化, 最終形成南極的南極棕囊藻和北半球的波切棕囊藻。Darling 等(2000, 2004)在有孔蟲的研究中也發現南北兩極之間確實存在跨越熱帶地區的基因流,為棕囊藻從南極向北極的傳播分化過程提供了間接證據。而最近Wu 等(2015)在熱帶海域南海西南沿海冷渦區域發現了大量南極棕囊藻和波切棕囊藻的存在, 與暖水種球形棕囊藻一起貢獻了9%—67.1%的葉綠素生物量。這些冷水種類到底從哪里來, 是本地起源還是外來物種, 其適應機制如何, 還無從考究,但這一發現或許能夠為棕囊藻兩極之間的傳播途徑提供直接的證據。
除了單細胞型和群體型, 近來還發現了介于二者之間的中間進化類型, 即2015 年在阿拉伯海域發現的新種冠狀棕囊藻, 目前僅觀察到單細胞形態存在, 但其細胞個體較其他棕囊藻種類都大, 尤其外被的多層不定形有機鱗片和指狀突起的葉綠體特征,與其他棕囊藻種有顯著的差別(Andersen et al, 2015)。系統進化分析結果顯示, 冠狀棕囊藻是介于單細胞種類(心形棕囊藻和揚棕囊藻)和群體型種類(球形棕囊藻、南極棕囊藻、波切棕囊藻)的中間進化類型, 在遺傳距離上更接近南極棕囊藻, 表明囊體的形成是后來衍生的特征(圖11, Andersen et al, 2015)。有關此種的地理分布范圍、生活史等諸多問題尚不得而知,但這一中間進化類型的發現無疑對于我們完整理解棕囊藻屬的系統進化具有重要意義。
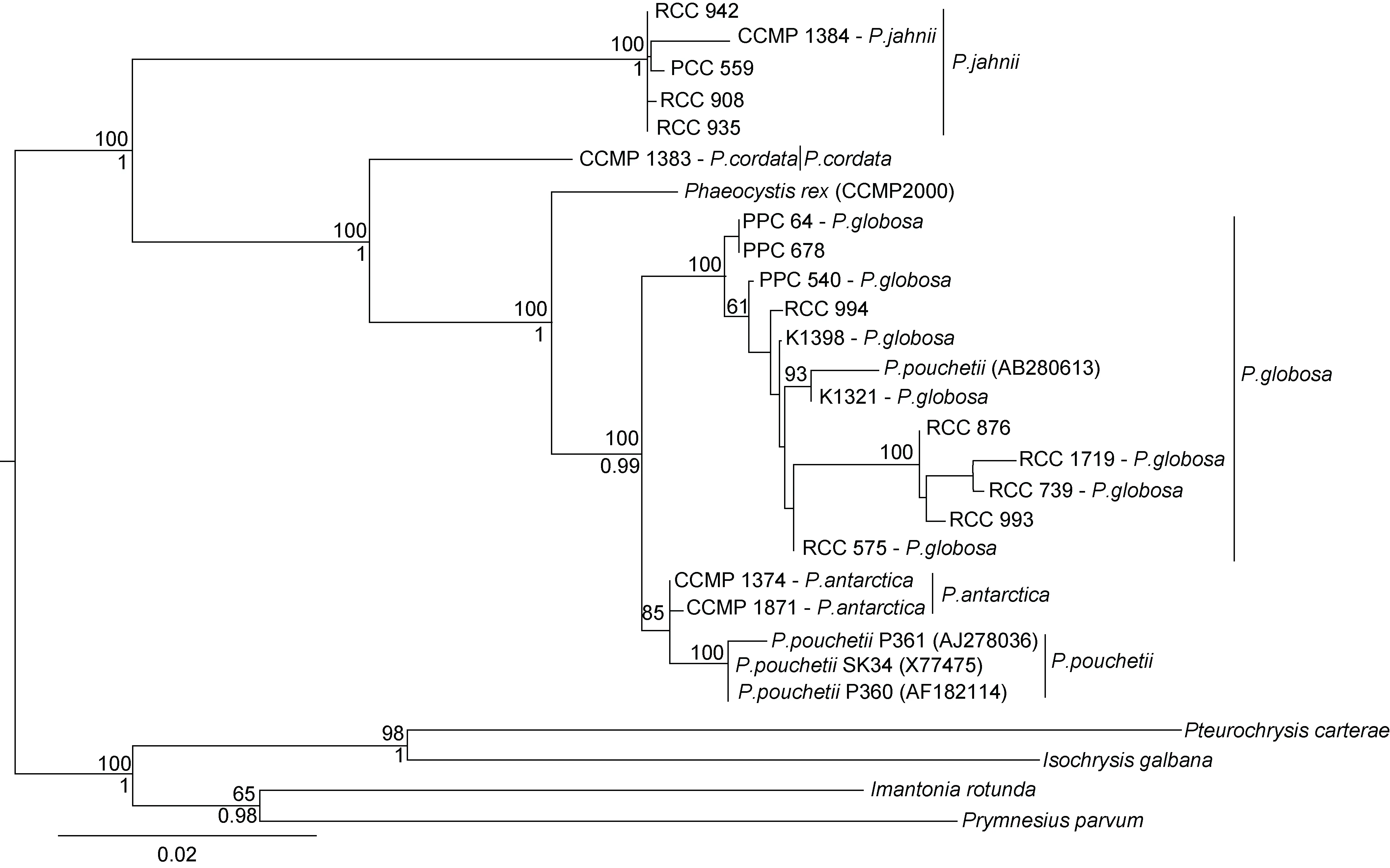
圖11 利用最大似然法(RAxML, 28 個種類, 4824 個對齊位置)基于5 種基因序列(線粒體16S rRNA, 18S rRNA and 28S rRNA, 質粒 psbA and rbcL 基因)構建的可培養棕囊藻種類的系統發育樹(Andersen et al, 2015)Fig.11 Phylogenetic tree of Phaeocystis inferred from a concatenated alignment of five genes (the ribosomal 16S rRNA, 18S rRNA and 28S rRNA, and the plastidial psbA and rbcL genes) using Random Axelerated Maximum Likelikhood(RAxML, 28 taxa with 4824 aligned positions) (Andersen et al, 2015)
3 展 望
根據棕囊藻屬不同類型種類發現的先后順序,如群體型種類先被發現, 后來發現的種類多為單細胞型, 推測那些未被描述的種類或者新種都應以單細胞種類為主(Andersen et al, 2015)。而且隨著環境樣品分子標記及高通量測序等深度、廣度技術的應用,出現了更多的“未培養克隆”棕囊藻種類(Lin et al,2014; Wu et al, 2015), 不僅分布極其廣泛, 而且豐度都較高。從生物多樣性的角度來看, 這些遺傳信息所代表的多樣性還需要從形態學上進一步相互驗證。但無論如何, 新的分子生物學技術的應用對于深入了解全球范圍內棕囊藻屬的種類多樣性及其地理分布特征、發現新的棕囊藻種類都具有極大的推動作用。棕囊藻屬的種類多樣性研究, 正面臨前所未有的機遇與挑戰, 相信今后在傳統的分類學與分子生物學組學等技術相結合的基礎上, 將更加清晰揭示全球棕囊藻的物種多樣性、遺傳多樣性與生態系統多樣性分布格局。
