蘇北淺灘筏架附生綠藻群落結構變化及與環境因子的相關性探究*
2021-02-03 01:44:22侯承宗唐學璽
海洋與湖沼 2021年1期
郝 雅 管 晨 侯承宗 唐學璽, 王 影,
(1. 中國海洋大學海洋生命科學學院 青島 266100; 2. 青島海洋科學與技術試點國家實驗室 海洋生態與環境科學功能實驗室 青島 266237)
自 2007 年以來, 大規模綠潮頻發于黃海海域,嚴重破壞了海洋生態環境, 造成了巨大的經濟損失(于仁成等, 2018; 王宗靈等, 2018)。通過DNA 分子鑒定, 黃海綠潮的致災種為石莼屬(Ulva)的滸苔(U.prolifera)(Xiao et al, 2013; Zhao et al, 2015)。通過對滸苔綠潮的溯源調查, 研究人員在滸苔綠潮起源的問題上達成廣泛共識: 黃海綠潮起源于江蘇省蘇北淺灘紫菜養殖筏架上附生的綠藻群落(Liu et al, 2009;Keesing et al, 2011; Zhou et al, 2015; 王廣策等,2020)。
蘇北淺灘筏架養殖區的綠藻群落作為滸苔綠潮的源頭, 在綠潮發生前的綠藻群落組成分析是滸苔綠潮早期防控的主要內容之一。研究人員對紫菜養殖筏架上附著的綠藻群落進行了種類鑒定(Tian et al,2011; Shen et al, 2012), 并且進一步估算了每種綠藻的生物量, 但是由于樣本數量、取樣時間和調查方法的不同, 結果之間存在很大差異。Pang 等(2010)和Shen 等(2012)發現紫菜養殖筏架上沒有或只有少量綠藻附著; Liu 等(2010)的現場調查中則發現有大量的滸苔附著在紫菜養殖筏架上; Han 等(2013)和 Fan等(2015)通過多次現場調查發現, 筏架附生的綠藻群落存在明顯的演替現象, 在不同的時期筏架上綠藻群落的優勢種也不同; 各調查結果不一致的原因可能是由于樣本量較少, 或者是綠藻群落結構的時空變化導致的隨機效應(Xiao et al, 2013)。在早期綠潮斑塊出現之前, 滸苔(U. prolifera)不是筏架附生綠藻群落的主要優勢種, 但是在綠潮暴發后, 滸苔在漂浮藻墊中的比例接近100% (Liu et al, 2015; Zhang et al,2015)。這些研究表明, 群落結構變化是綠潮暴發前蘇北淺灘綠藻群落的鮮明特征, 并且對綠潮的發生具有重要影響, 解析其變化的原因對滸苔綠潮的早期預警具有重要意義。
綠藻群落結構的改變是生物與環境相互作用的結果, 已有研究尚缺乏與環境因素的相關性分析。溫度、鹽度、營養鹽含量和光照強度等環境因素對大型綠藻群落結構組成和演替具有顯著影響(Fong et al,1996; 方松等, 2012)。為了找出導致綠藻群落結構變化的關鍵環境因素, 作者在蘇北淺灘主要的筏架養殖區進行連續采樣, 對綠藻樣品進行了分類鑒定和生物量統計, 并結合環境因子進行了相關性分析, 探明蘇北淺灘綠藻群落結構變化特征的同時, 找出導致筏架附生綠藻群落結構變化的關鍵環境因素, 為滸苔綠潮的早期預警和防控提供生態學資料支持。
1 材料與方法
1.1 調查海域與站位
本次調查區域位于江蘇省南通市如東縣東北部的蘇北淺灘, 該區域是全國最大的條斑紫菜筏架養殖區, 一共設置了 6 個樣品采集站位(圖 1)。綠藻樣品采集時間為 2019 年 3 月 10 日、4 月 15 日和 5 月15 日, 每個采樣站點隨機選擇5 個紫菜養殖筏架, 在筏架上每次刮取50 cm 長度的綠藻樣品, 每個筏架重復取樣3 次。采集后的綠藻樣品在現場用過濾海水清洗掉表面附著的泥沙等雜質, 用吸水紙吸干表面水分, 用樣品袋封裝并編號, 然后置于4°C 的便攜式冰箱(米其林, 中國)中送回實驗室進行形態學分類和分子生物學鑒定。采樣地點的海水溫度、鹽度和pH 值使用手持式野外多參數水質檢測儀(YSI Orion, 美國)測定; 光照強度使用雙輻照儀(卓川, 中國)在晴朗天氣下進行測量, 測量時間為正午11—13 點, 視天氣情況篩除極端測量值, 用光合有效輻射(photosynthetically active radiation, PAR)來表示光照強度的測量結果,單位為 μmol photons/(m2·s)。
1.2 形態鑒定及生物量統計
送回實驗室的綠藻樣品再次用無菌海水沖洗,并用毛刷仔細清潔。清潔后的綠藻以株為單位分開,置于添加有無菌海水的玻璃培養皿中, 用鑷子輕輕展開藻體, 在體式顯微鏡(Olympus SZ51, 日本)下進行綠藻的形態學鑒定(圖 2)。綠藻樣品的形態學鑒定主要依據藻體顏色、基部情況(固著器等)、藻體外形輪廓、主軸和分支特征等 (Tseng, 1983;丁蘭平等, 2009; 余秋瑢等, 2017)。鑒定完畢后的綠藻樣品用吸水紙充分吸干表面水分, 并使用高精度電子天平(BSA124S, 中國)稱重, 藻體重量精確到 1 mg,通過匯總所有物種的生物量來計算綠藻群落的總生物量(濕重)。
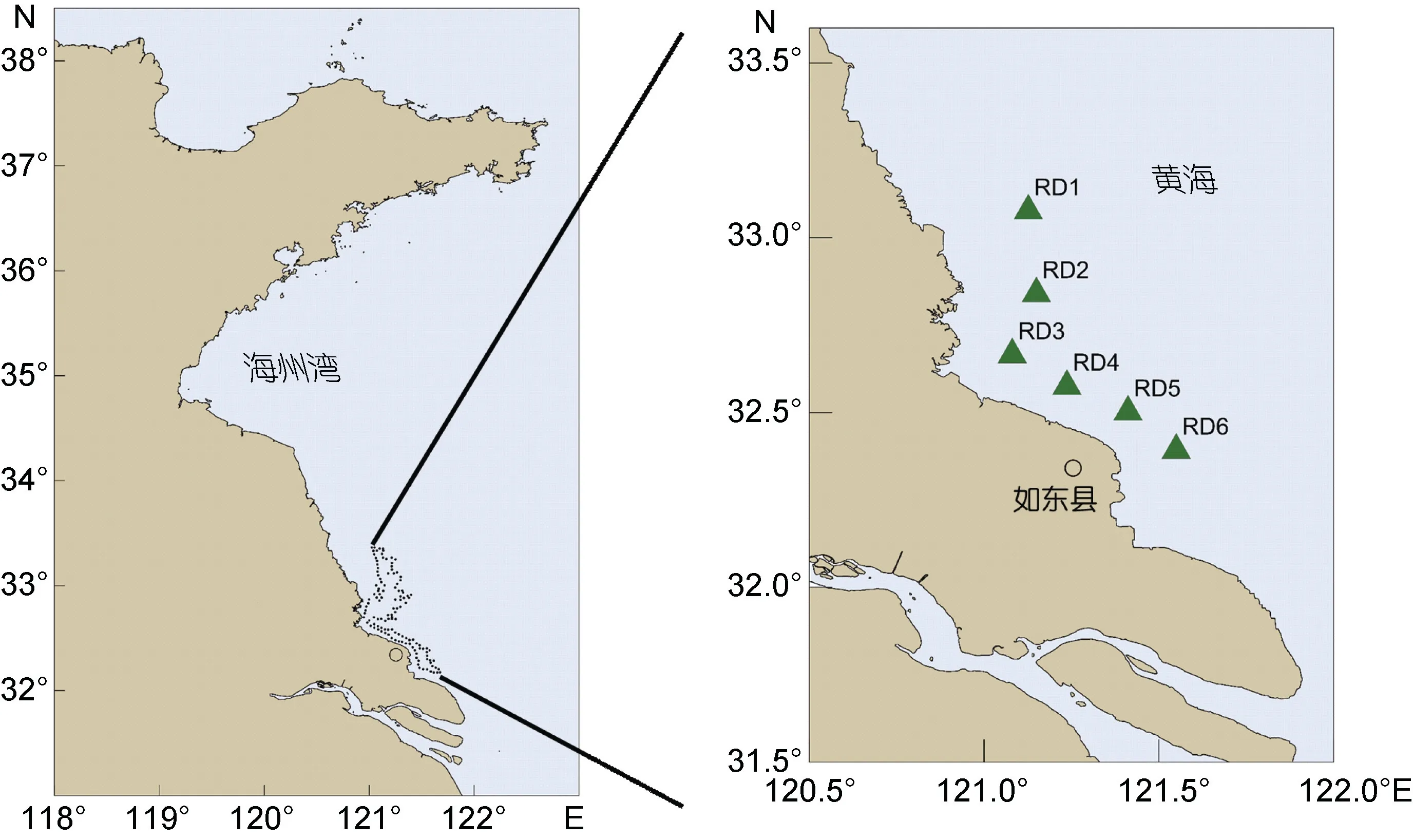
圖1 蘇北淺灘綠藻采集站位Fig.1 The sampling site of green macroalgae in the Subei Shoal
1.3 DNA 提取與序列分析
為了對綠藻形態學鑒定結果進行補充和驗證,本文在3 月和5 月的綠藻樣品中隨機選擇了60 株綠藻樣品進行了 ITS 序列(internal transcribed spacer,ITS)和5S rDNA 間隔區序列(5S rDNA spacer)分析。綠藻樣品用去離子水多次清洗后用吸水紙吸干水分,將藻體置于研缽中加入液氮充分研磨成組織粉末,選用植物基因組 DNA 試劑盒(北京天根生化科技有限公司)進行后續的全基因組DNA 提取。用于序列分析的樣品的命名方式為“采集月份-站位-樣品編號”的方式。提取后的綠藻DNA 樣品被轉移到2mL 凍存管中并置于液氮中保存, 隨后交付上海生工公司(生工,上海 )進行PCR (polymerase chain reaction)、引物合成和ITS 與5S rDNA 間隔區序列的雙向測序, PCR 引物序列如表1 所示(Xiao et al, 2013)。

表1 ITS 與5S rDNA 間隔區 PCR 引物序列Tab 1 The PCR primers of ITS and 5S rDNA spacer
1.4 進化樹分析
將1.3 的測序結果錄入NCBI(美國國家生物技術信息中心) (http://www.ncbi.nlm.nih.gov)進行Blast 同源性比對。使用 MEGA 5 進行了最大似然法(maximum-likelihood, ML)和鄰接法(neighbor-joining,NJ)分析(Tamura et al, 2011), 以 Umbraulva amamiensis作為外類群。通過Bootstrap 方法計算支持率(1000 次重復, 省略支持率小于50%的數值), 采用環式(Circle)進化樹模型構建文本序列分析的進化樹。
1.5 數據處理及分析
所有數據分析均為三個重復, 并以平均標準差(standard deviation, SD)表示。實驗數據采用單因素方差分析(One-way ANOVA)進行檢驗, 如果方差分析顯示有顯著差異, 則采用Student-Newman-Keuls 檢驗進行事后多重比較。本文使用的統計分析軟件為IBM SPSS 22.0 (SPSS Inc, 美國), P<0.05 認為有統計學意義。冗余分析(redundancy analysis, RDA)采用軟件Canoco 5.0 進行。
2 結果與討論
2.1 綠藻樣品的形態學鑒定
通過形態學特征比對, 在采集的綠藻樣品中共鑒定出滸苔(U. prolifera), 盤苔(Blidingia sp.), 緣管滸苔(U. linza) 3 種大型綠藻和無法通過形態學特征進一步區分的綠藻混合種(圖 2), 根據之前的調查結果, 本文中的綠藻混合種可能包含有曲滸苔(U.flexuosa)和扁滸苔(U. compressa)等形態結構較為相似的筏架附生綠藻(Huo et al, 2013; Fan et al, 2015)。
2.2 綠藻樣品的序列分析
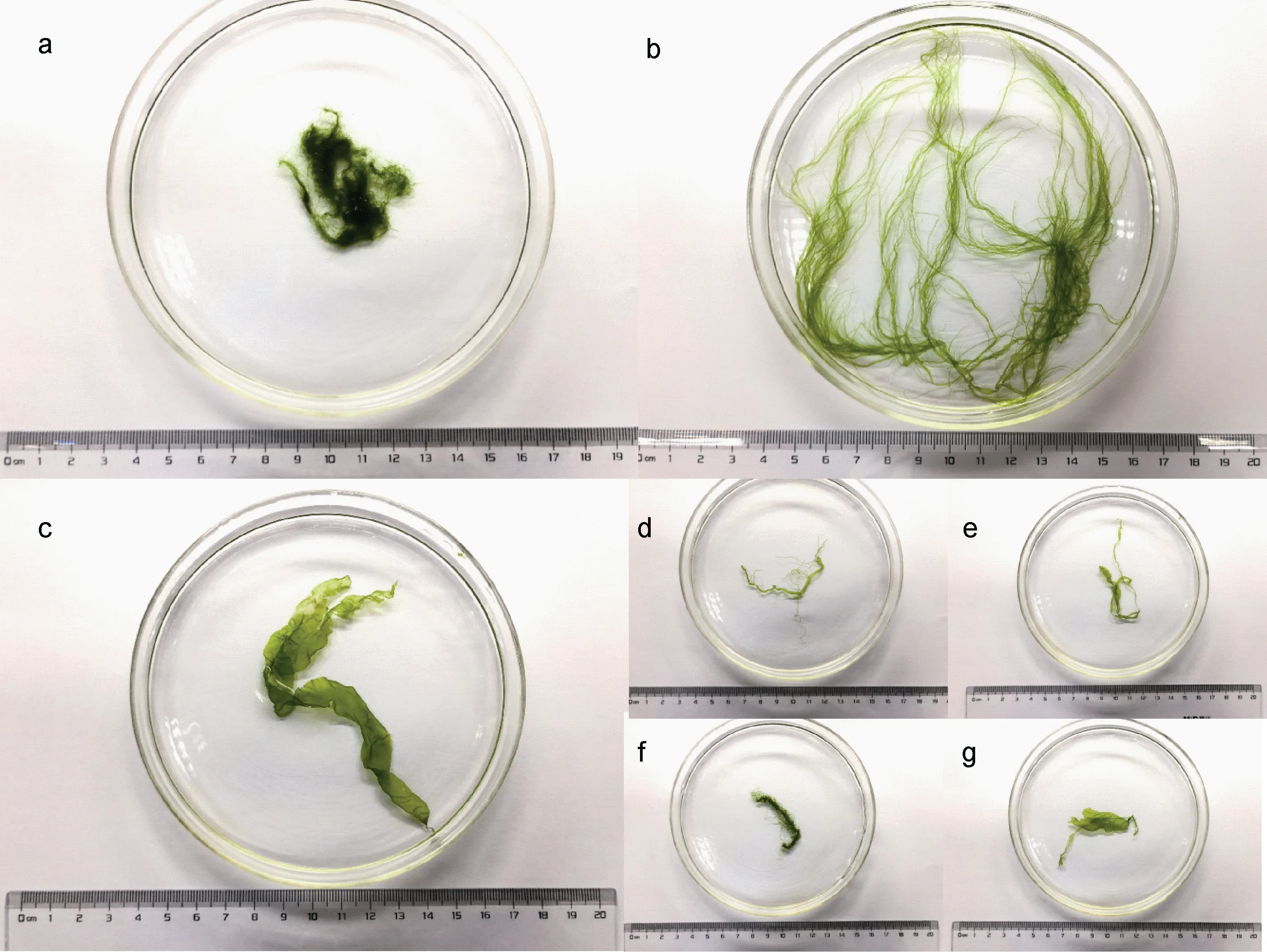
圖2 筏架附生綠藻群落中的各種綠藻Fig 2 Various green macroalgae attached onto the rafts
ITS 序列分析結果顯示, 綠藻樣品主要由石莼屬綠藻構成的復合類種群 LPP (Ulva linza-proceraprolifera)、盤苔(Blidingia sp.)和曲滸苔(U. flexuosa)組成(圖3)。由于ITS 序列無法明確區分LPP 內的綠藻種類, 對分布在LPP 區域內的27 個綠藻DNA 樣品進行了基于5S rDNA 的序列分析。5S 序列分析結果表明, 27 個LPP 樣品中有20 個屬于滸苔(U. prolifera),7 個屬于緣管滸苔(U. linza) (圖4)。
2.3 筏架附生綠藻的生物量變化特征
在生物量統計過程中, 無法通過形態學進行分類的綠藻樣品被合并為石莼屬綠藻(Ulva spp.)。筏架附生綠藻的總生物量從 3 月的29.95 g/m 增加到了 5月的108.41 g/m (圖5b)。研究人員對蘇北淺灘海域綠藻微觀繁殖體的研究表明, 綠藻微觀繁殖體的數量在4 月下旬達到最大值, 筏架附生綠藻生物量在五月達到峰值(Li et al, 2014; Wang et al, 2018), 這與本文的生物量統計結果一致。調查期間綠藻生物量統計結果表明, 蘇北淺灘的筏架附生綠藻群落具有明顯的演替現象(圖6)。3 月在各站點的綠藻群落中沒有發現或者零星出現少量滸苔(U. prolifera), 4 月滸苔在綠藻群落的比例上升到了20%, 5 月滸苔在綠藻群落中的比例達到了接近40% (圖6), 滸苔的生物量從3 月的1.65 g/m 增加到了5 月的31.78 g/m (圖5a); 石莼屬綠藻(Ulva spp.)在綠藻群落中的比例則表現出與滸苔(U.prolifera)相反的變化趨勢(圖 7); 緣管滸苔的生物量在調查期間的變化不顯著(One way ANOVA, P>0.05,圖 5a); 盤苔在調查期間一直是筏架附生綠藻群落中的優勢種(圖5a, 6), Song 等(2015)對蘇北淺灘水體和沉積物的調查中未檢測出盤苔的微觀繁殖體, 并推斷營養生殖是盤苔在筏架區增加種群數量的主要方式, 對于此推論我們還需要進一步的研究求證。
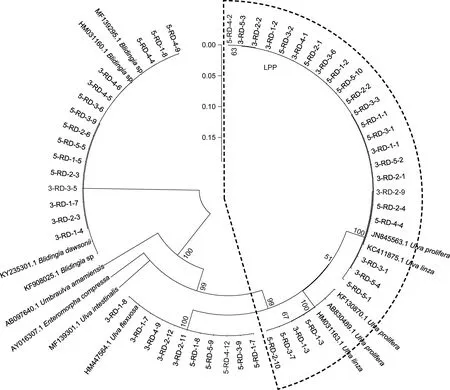
圖3 筏架附生綠藻樣品基于ITS 序列分析的鄰接樹[Neighbor-joining (NJ) tree]Fig.3 Neighbor-joining (NJ) tree of green macroalgae samples on the rafts in Subei Shoal based on ITS sequence analysis
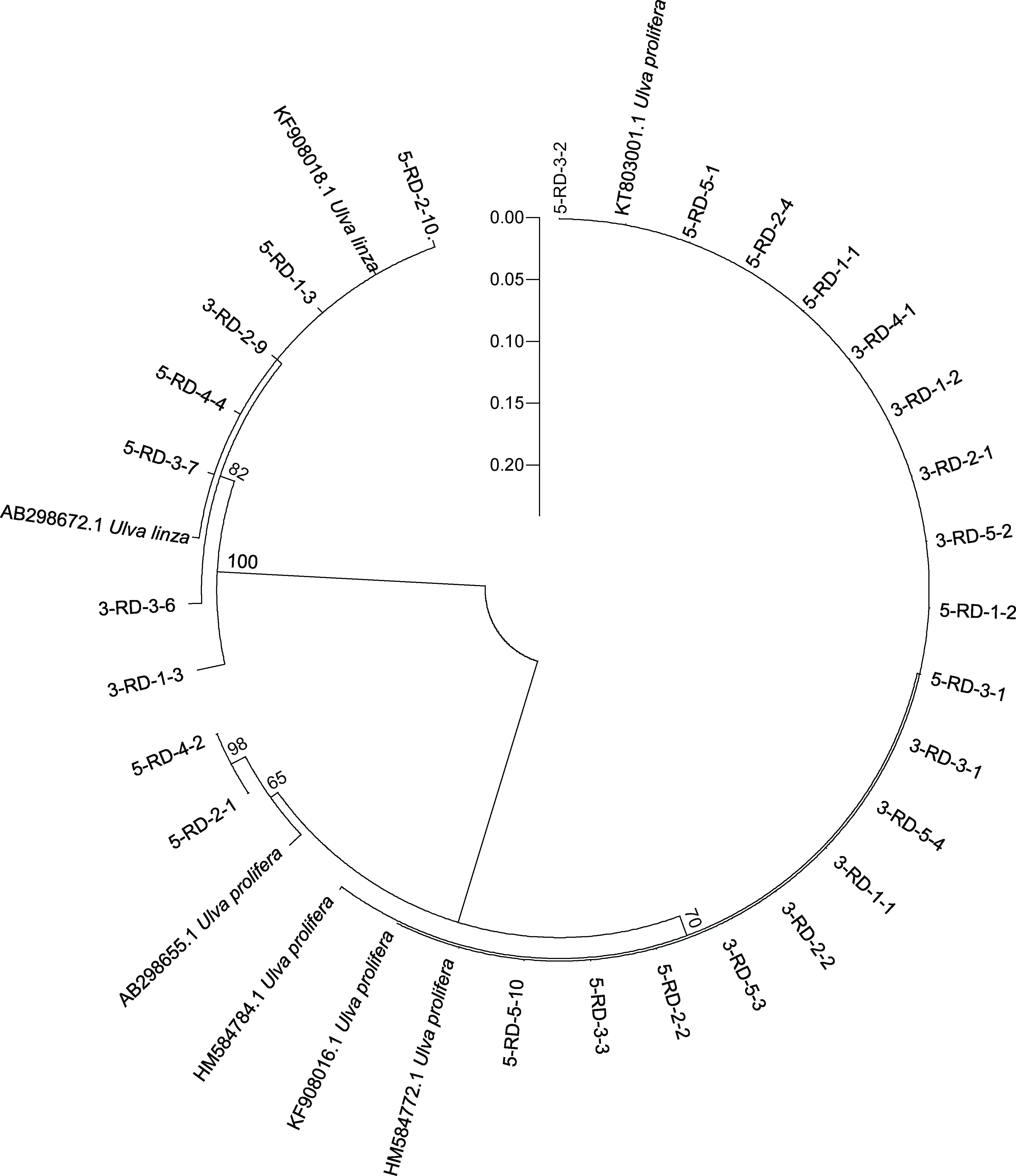
圖4 筏架附生綠藻樣品基于5S 序列分析的鄰接樹Fig.4 Neighbor-joining (NJ) tree of green macroalgae samples on the rafts in Subei Shoal based on 5S sequence analysis
2.4 蘇北淺灘環境因子與綠藻生物量變化的相關性分析
蘇北淺灘紫菜養殖筏架區 3—5 月的環境因子變化情況如表2 所示, 從3 月到5 月, 蘇北淺灘的環境溫度呈顯著上升的趨勢(One-way ANOVA , P<0.05),平均溫度從8.32°C 上升到了18.38°C。海水鹽度、光照強度和海水pH 在調查期間沒有顯著變化(One-way ANOVA , P>0.05)。
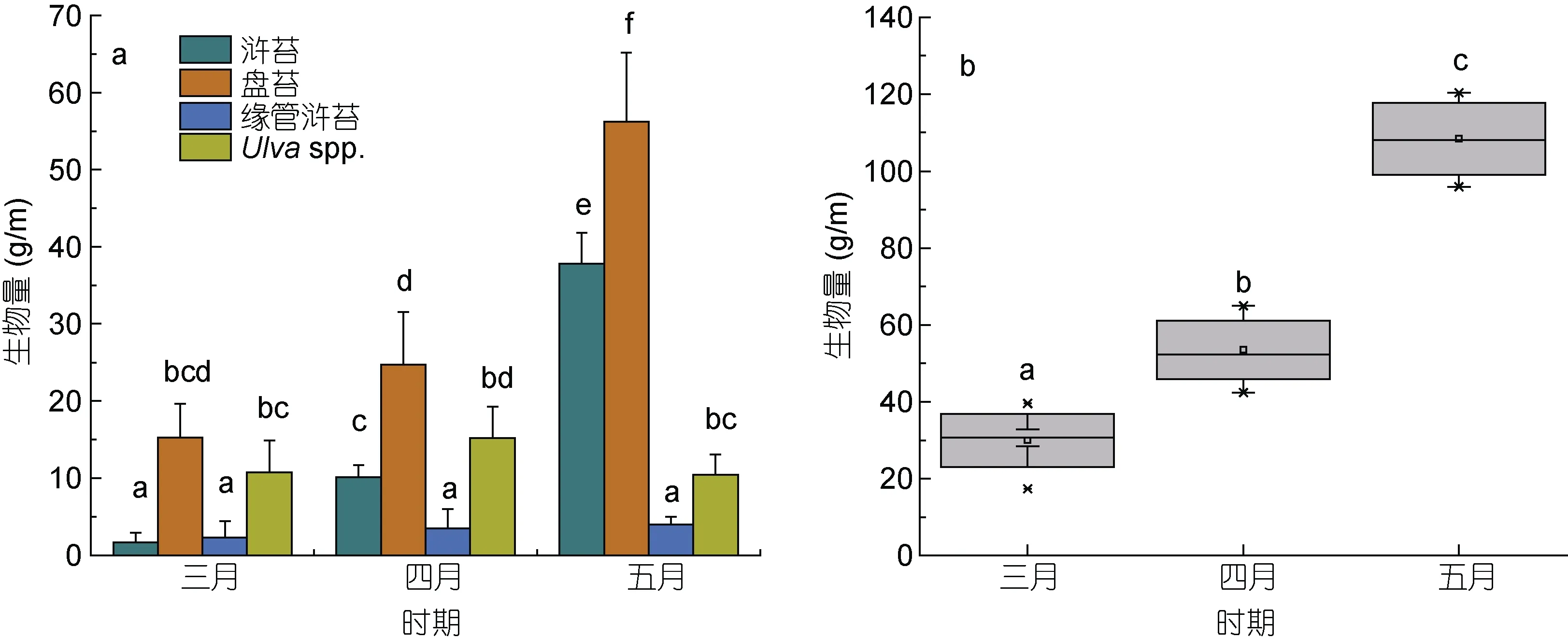
圖5 3—5 月紫菜筏架附生綠藻生物量變化情況Fig.5 Biomass changes of green macroalgae on the rafts in Subei Shoal from March to May
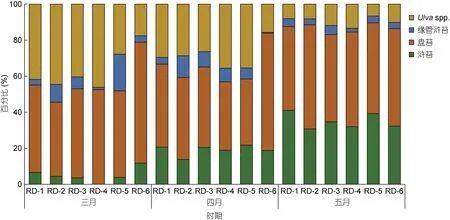
圖6 蘇北淺灘筏架附生綠藻群落的組成比例變化Fig.6 Change in composition proportion of the green macroalgae community on the rafts in Subei Shoal
研究證明不同的大型藻類對環境因子變化的響應存在明顯差異(Dawes et al, 1978), 溫度、鹽度和光照強度可以決定藻類的分布特征(Wiencke et al,1980)。在本研究中, 四種環境因子對生物量變化的總解釋度為59.9% (表3)。溫度對綠藻生物量變化的解釋度最高(47.8%), 因此溫度是導致筏架附生綠藻群落結構改變的主要環境因子(表3)。在本次調查期間,環境溫度與滸苔(U. prolifera)種群生物量呈正相關(圖 7)。Song 等(2015)的研究證明, 環境溫度低于 10°C時滸苔的微觀繁殖體不會生長, Pang 等(2010)的研究也證實在 3—4 月蘇北淺灘沒有或只有很少的滸苔。15—20°C 是滸苔生長較適宜的溫度(王建偉等, 2007),蘇北淺灘在 3—5 月的溫度變化范圍是 8.3—18.4°C(表2), 因此, 從3 月到5 月, 蘇北淺灘的環境溫度逐漸接近滸苔(U. prolifera)生長的適宜溫度, 該區域滸苔種群的生物量也隨之迅速增加。在筏架綠藻群落中,扁滸苔(U. compressa)的最適生長溫度是 10°C (王陽陽等, 2010), 腸滸苔(Ulva intestinalis)和條滸苔(Ulva clathrata)也已被證明是適低溫性的綠藻(Wiencke et al, 1990), 因此5 月上升的環境溫度不利于筏架上適低溫性的綠藻的生存, 逐漸被滸苔等能夠適應較高環境溫度的綠藻取代。
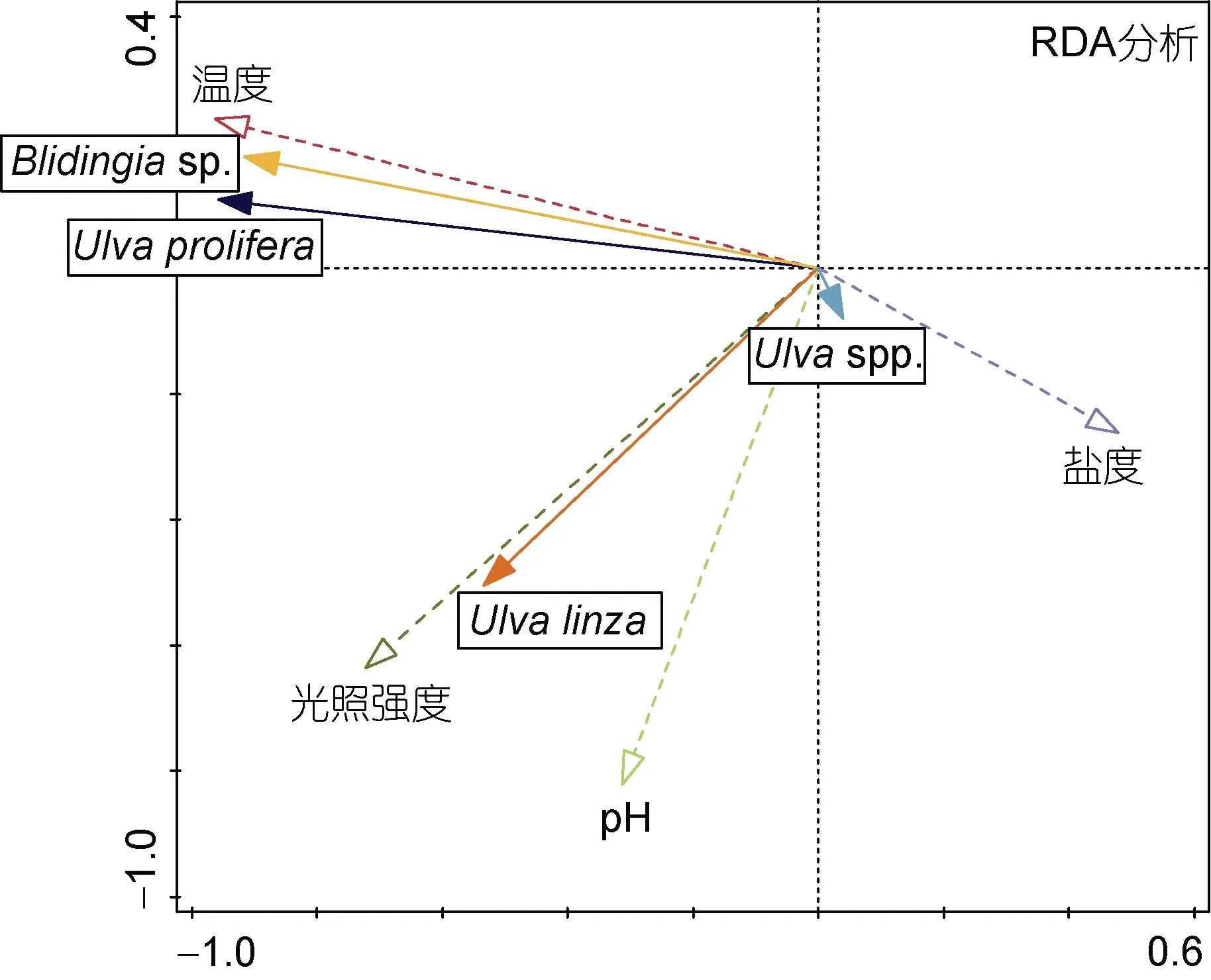
圖7 針對環境變量與綠藻生物量變化相關性的冗余分析(RDA)結果Fig.7 Redundancy analysis (RDA) results of the correlation between environmental variables and biomass changes of green macroalgae
除溫度以外, RDA 分析結果顯示光照強度也對綠藻群落結構產生了較大影響(表3, 圖7), 滸苔和緣管滸苔的生物量對光照強度表現出較強的正相關性(圖 7)。Xiao 等(2016)通過培養實驗發現筏架附生的滸苔對光照強度的耐受性較高, 而Kim 等(2011)也發現發現緣管滸苔對高光強也具有很強的適應性, 這與本次RDA 分析結果相符。盤苔和滸苔的生物量與海水鹽度呈負相關(圖 7), 這可能與調查期間地表徑流和降水量的變化有關。南黃海沿岸河流眾多, 除了長江以外, 還有射陽河和關河等地方河流以及眾多人工運河(趙全基, 1992), 大量的地表徑流的匯入在降低海水鹽度的同時, 提高了蘇北淺灘海域的營養鹽負荷, 為大型藻類的生長提供了外源營養物質補充(Mieszkin et al, 2013), 筏架綠藻群落的生物量變化與海水鹽度的關系還需要進一步的研究求證。
3 結論
(1) 蘇北淺灘紫菜養殖筏架上附生的綠藻群落主要由滸苔、盤苔、緣管滸苔、曲滸苔和其他綠藻5個類群構成。
(2) 綠潮暴發前期(3—5 月), 筏架附生綠藻群落具有明顯的演替現象, 溫度是導致綠藻群落結構改變的關鍵環境因素, 適低溫性的綠藻逐漸被滸苔等能夠適應較高環境溫度的綠藻取代。

表2 蘇北淺灘環境因子變化情況Tab.2 Changes of environmental factors in Subei Shoal
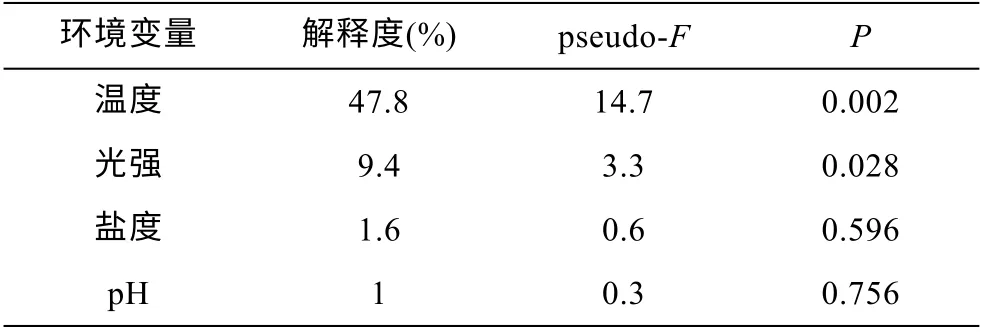
表3 環境因子對綠藻生物量變化的解釋度Tab.3 The interpretation value of environmental factors to the change in biomass of the green macroalgae
(3) 滸苔的生物量從3 月份的1.65 g/m 增加到了5 月的31.78 g/m, 所占比例從低于5%達到接近40%,滸苔生物量的積累為蘇北淺灘早期綠潮漂浮斑塊的出現提供了物質基礎。
