‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交F1代果實(shí)品質(zhì)的遺傳變異分析*
2021-02-05 07:30:56王鵬飛付鴻博穆霄鵬張建成杜俊杰
中國果樹 2021年1期
王鵬飛,付鴻博,2,穆霄鵬,張建成,杜俊杰
(1 山西農(nóng)業(yè)大學(xué)園藝學(xué)院,太谷030801)(2 黑龍江省農(nóng)業(yè)科學(xué)院鄉(xiāng)村振興科技研究所)
歐李[Cerasus humilis(Bge.)Sok.]屬薔薇科櫻桃屬矮生櫻亞屬植物,又稱鈣果,為我國特有的灌木果樹資源。歐李果實(shí)富含氨基酸、維生素、有機(jī)酸及多種礦質(zhì)元素,并含有較高的花色苷、黃酮醇、黃烷醇、單寧等物質(zhì),是一種兼具生態(tài)效益和經(jīng)濟(jì)價(jià)值的灌木樹種,具有高營養(yǎng)、高抗性、高保健的特點(diǎn),研究和開發(fā)利用前景十分廣闊[1]。隨著生活水平的不斷提高,果實(shí)品質(zhì)日益受到關(guān)注。果實(shí)的外觀品質(zhì)包括了大小、形狀及色澤,這些也是判斷果品商品價(jià)值的重要指標(biāo),極大程度地影響了消費(fèi)者的選擇[2-3]。類黃酮是植物中一類重要的次生代謝物質(zhì),具有強(qiáng)烈的生物活性,在影響果實(shí)的色澤和風(fēng)味等方面有重要作用[4],且具有抗氧化、抗衰老以及抵抗病毒等功能,在人體內(nèi)能夠抑制腫瘤、抵抗誘變和改善人體微循環(huán)[5-6]。
雜交育種是果樹遺傳性狀改良的重要手段之一,目標(biāo)性狀的遺傳變異越大,從雜交后代中選出優(yōu)異單株的概率越高。雜交育種往往能創(chuàng)造出所需的新基因型,其性狀很可能比雙親更有優(yōu)勢,還可以利用雜交手段組配特殊的性狀,以解決生產(chǎn)上的特殊需求,因而雜交育種是育種中一個(gè)非常重要的手段[7-9]。
歐李植株矮小,占地面積小,當(dāng)年播種,第2年即可結(jié)果,是研究后代遺傳分析具有明顯優(yōu)勢的果樹樹種。目前關(guān)于次生代謝物質(zhì)的遺傳變異分析相對較少,本研究通過構(gòu)建歐李F1代群體,研究歐李果實(shí)品質(zhì)性狀的遺傳特點(diǎn)及傾向,是指導(dǎo)歐李雜交育種工作的前提,以期從雜交后代中選育出所需目標(biāo)性狀優(yōu)株,并且為科學(xué)選配親本提供理論依據(jù)。
1 材料和方法
1.1 試驗(yàn)材料
試驗(yàn)材料取自山西農(nóng)業(yè)大學(xué)巨鑫試驗(yàn)園,以‘農(nóng)大6 號’和‘農(nóng)大7 號’為雙親進(jìn)行雜交,雜交后代于2016 年11 月栽植,行株距為0.6 m×0.4 m,按常規(guī)水肥方法進(jìn)行田間管理,管理措施一致,2018 年于‘農(nóng)大6 號’ב農(nóng)大7 號’(正交)雜交后代中選出55 株結(jié)果株,于‘農(nóng)大7 號’ב農(nóng)大6 號’(反交)雜交后代中選出32 株結(jié)果株,測定雙親及雜交后代群體果實(shí)的單果重、縱橫徑、色澤、類黃酮含量和總酚含量。
1.2 試驗(yàn)方法
1.2.1 單果重及縱橫徑的測定
選取10 個(gè)果實(shí),采用電子天平稱量果實(shí)重量,采用電子數(shù)顯卡尺測定縱徑、橫徑,果形指數(shù)用果實(shí)縱徑和橫徑的比值表示。
1.2.2 果實(shí)色澤參數(shù)測定
選取5 個(gè)果實(shí),利用分光測色儀對其色澤進(jìn)行測定。每個(gè)果實(shí)選擇其赤道與縫合線交叉點(diǎn)開始測定,沿赤道每旋轉(zhuǎn)90°測一點(diǎn),共測4 個(gè)點(diǎn),計(jì)算平均值,然后5 個(gè)果實(shí)再算平均值。記錄L*、a*和b*值,并計(jì)算h°(Hue)、C*(chroma)、CI 值;其中L*值(0~100.0)表示亮度,L*值越大則表示亮度越大;a*值表示紅綠,a*值為負(fù)時(shí)表示綠色,為正時(shí)表示紅色,絕對值越大則表示顏色越深;b*值表示黃藍(lán),b*值為負(fù)時(shí)表示藍(lán)色,為正時(shí)表示黃色,絕對值越大則表示顏色越深;h°值表示色度角,C*表示色澤飽和度[2];CI 為正值時(shí)表示紅黃程度,為負(fù)值時(shí)表示藍(lán)綠程度。
1.2.3 類黃酮及總酚含量的測定
(1)提取液的制備。利用超聲輔助提取法[10],用液氮將果實(shí)研磨成粉末,用40%的酸化甲醇作提取液,料液比1∶10(g∶mL),漩渦震蕩混勻,在不高于40 ℃的溫度下超聲提取30 min(40 kHz),后在10 000×g下離心15 min,重復(fù)提取3 次,合并濾液,待測定。
(2)類黃酮含量的測定。采用NaNO2-Al(NO3)3-NaOH比色法測定[10],吸取一定體積的提取液于10 mL 容量瓶,先加5%亞硝酸鈉溶液0.3 mL,搖勻,避光靜置6 min,再加10%六水硝酸鋁溶液0.3 mL,搖勻,避光靜置6 min,再加4%氫氧化鈉溶液4 mL,定容到10 mL,搖勻靜置10 min,510 nm 處測定吸光值。以標(biāo)準(zhǔn)品蘆丁作標(biāo)準(zhǔn)曲線。
(3)總酚含量的測定。采用福林酚比色法[10]測定,吸取一定體積的提取液于10 mL 離心管中,先加入福林酚試劑0.2 mL,搖勻,靜置4 min,加入10%碳酸鈉溶液1 mL,再加入ddH2O 定容至8 mL,在35 ℃下水浴1 h,760 nm 處測定吸光值。以標(biāo)準(zhǔn)品沒食子酸作標(biāo)準(zhǔn)曲線。
1.3 數(shù)據(jù)分析
采用Microsoft Excel 2007 軟件進(jìn)行數(shù)據(jù)分析。
1.3.1 色澤相關(guān)指標(biāo)計(jì)算公式
h°=tan-1(b*/a*);當(dāng)a*<0、b*>0 時(shí),h°=180°+tan-1(b*/a*)
C*=(a2+b2)/2
CI=1 000×[a*/(L*×b*)]
1.3.2 遺傳相關(guān)指標(biāo)計(jì)算公式
超高親比率(%)=∑hp/n×100[11]
超低親比率(%)=∑lp/n×100[11]
變異系數(shù)(%)=S/F×100
廣義遺傳力(%)=[VH-(Vp1+Vp2)/2]/VH×100[12]
hp 表示高于高親的群體后代數(shù)、lp 表示低于低親的群體后代數(shù)、n 為群體總數(shù);S 和F 分別為后代的標(biāo)準(zhǔn)差和平均值;VH 為雜交群體的表現(xiàn)型方差,Vp1 為母本表現(xiàn)型方差,Vp2 為父本表現(xiàn)型方差。
2 結(jié)果與分析
2.1 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代單果重的遺傳變異
由表1 可以看出,正反交后代的單果重平均值均小于親中值,正反交后代中低于低親的比率分別為72.73%和93.75%,具有明顯的衰退變異現(xiàn)象。反交后代單果重平均值低于正交后代,說明當(dāng)‘農(nóng)大6 號’作為父本時(shí),更容易將單果重較小的性狀傳遞給后代。由此可見,在‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代中單果重更易受父本的影響。當(dāng)單果重較大的‘農(nóng)大7 號’作為父本時(shí),超高親比率達(dá)12.73%,可以從后代中選出果個(gè)較大的單株,正反交的變異系數(shù)均超過了20%,在雜交后代中均出現(xiàn)了廣泛分離,選擇潛力較大。正反交廣義遺傳力均較高,在90%以上,表明單果重的變異主要來自遺傳效應(yīng)。從圖1 可以看出,正反交單果重均呈正態(tài)分布,表現(xiàn)為連續(xù)變異。
2.2 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)縱橫徑的遺傳變異
由表2、3 可以看出,正反交后代的縱橫徑平均值均小于親中值,表現(xiàn)為衰退變異,其中正反交縱徑超高親比率均是0,低于低親的比率達(dá)到了92.73%和93.75%。正反交橫徑超高親比率也只有7.27%和6.25%,低于低親的比率均超過了75.00%。正反交縱橫徑的變異系數(shù)均較小,選擇潛力較小。廣義遺傳力均較大,變異主要受遺傳效應(yīng)影響。由
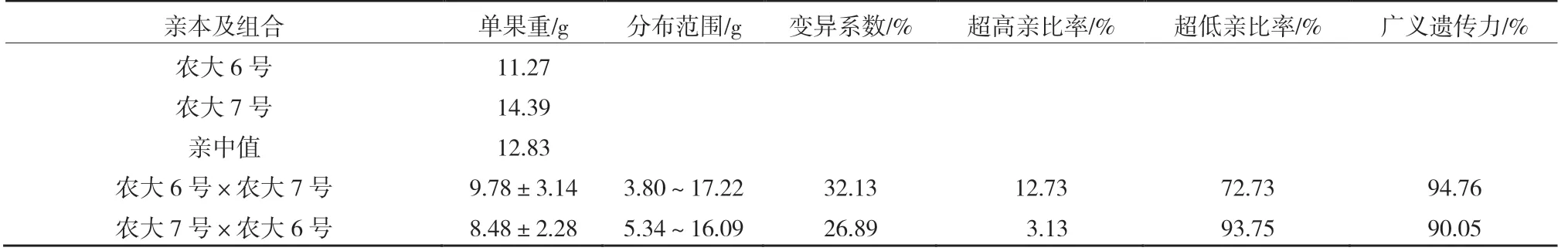
表1 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代單果重的遺傳變異
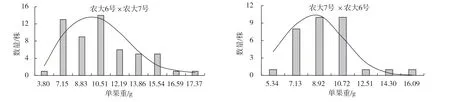
圖1 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代單果重的頻率分布
圖2、3 可以看出,正反交縱橫徑均呈正態(tài)分布,表現(xiàn)為連續(xù)變異。
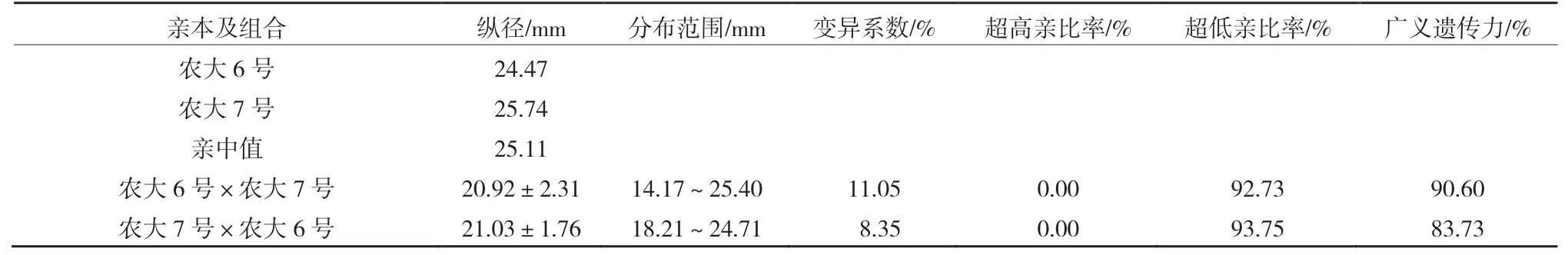
表2 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)縱徑的遺傳變異

圖2 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)縱徑的頻率分布
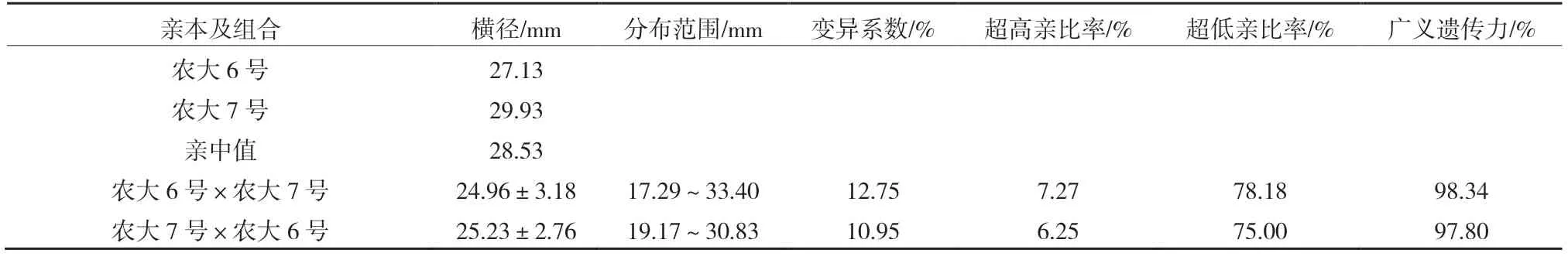
表3 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)橫徑的遺傳變異
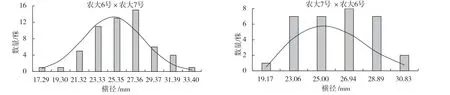
圖3 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)橫徑的頻率分布
2.3 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果形指數(shù)的遺傳變異
由表4 可以看出,正反交后代果形指數(shù)均小于親中值,且低于低親的比率分別為 70.91%和65.63%,表現(xiàn)為明顯的衰退變異。變異系數(shù)均較低,分別為6.82%和8.25%,說明后代選擇潛力較低。廣義遺傳力較高,分別為82.81%和88.15%,表面后代主要受遺傳效應(yīng)影響。
由圖4 可以看出,正反交果形指數(shù)均呈正態(tài)分布,表現(xiàn)為連續(xù)變異。
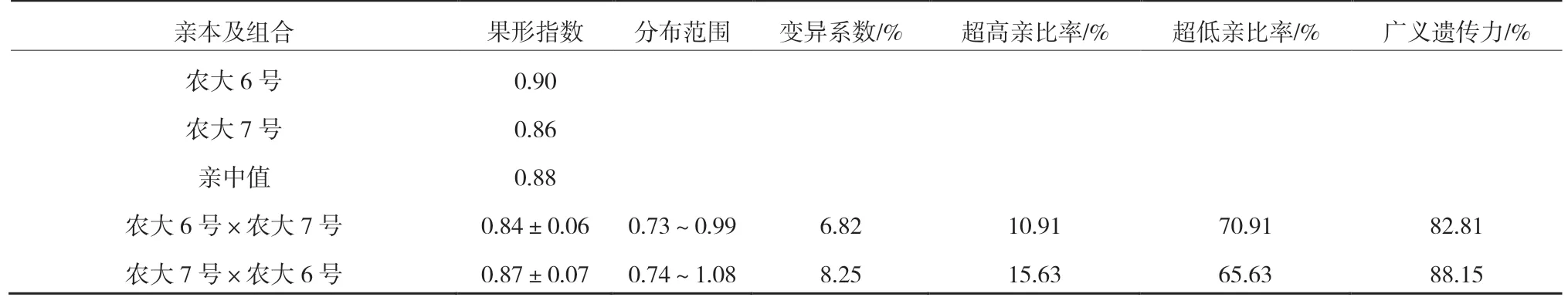
表4 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果形指數(shù)的遺傳變異
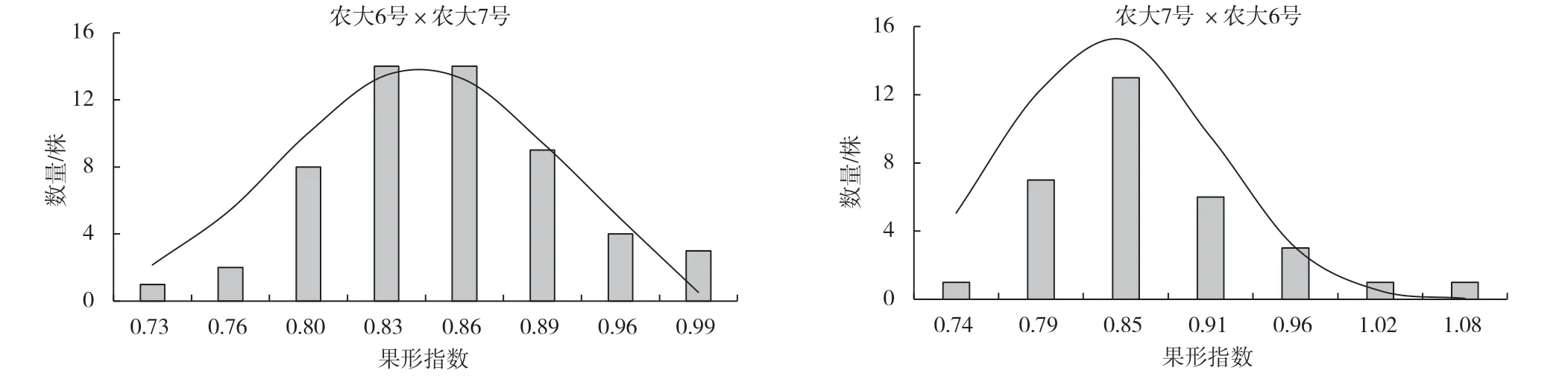
圖4 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果形指數(shù)的頻率分布
2.4 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)類黃酮含量的遺傳變異
由表5 可以看出,正反交果實(shí)類黃酮含量均高于親中值,且類黃酮含量較高的‘農(nóng)大6 號’作為父本時(shí),超高親比率達(dá)到了46.88%,更易篩選出高黃酮類型的歐李單株。變異系數(shù)均超過了20%,說明出現(xiàn)了廣泛分離現(xiàn)象,具有較高的選擇潛力。廣義遺傳力較高,表明果實(shí)類黃酮含量主要受遺傳效應(yīng)的影響。由圖5 可知,正反交果實(shí)類黃酮含量均呈正態(tài)分布,表現(xiàn)為連續(xù)變異。
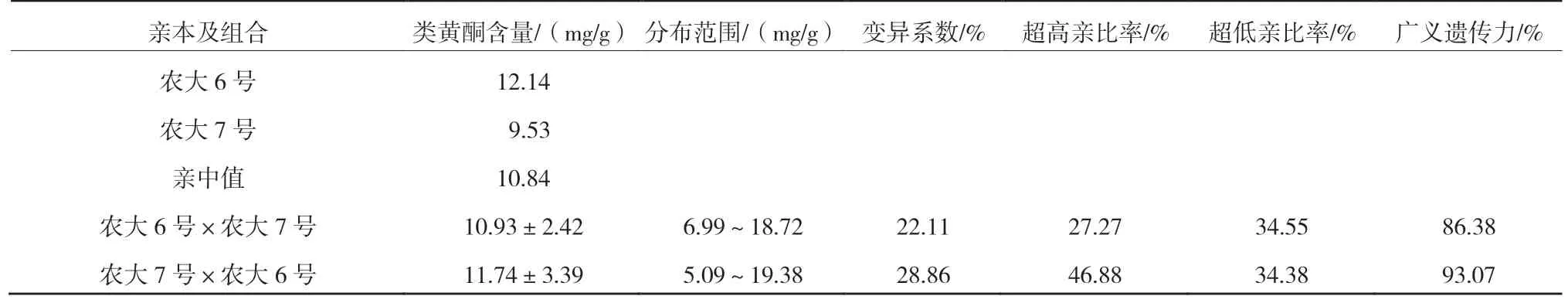
表5 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)類黃酮含量的遺傳變異

圖5 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)類黃酮含量的頻率分布
2.5 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)總酚含量的遺傳變異
由表6 可以看出,正反交果實(shí)總酚含量均高于親中值,且總酚含量較高的‘農(nóng)大6 號’作為父本時(shí),超高親比率達(dá)到了50.00%,更易篩選出高多酚類型的歐李單株。反交后代變異系數(shù)為22.27%,說明出現(xiàn)了廣泛分離現(xiàn)象,具有較高的選擇潛力。廣義遺傳力較高,表明果實(shí)總酚含量主要受遺傳效應(yīng)的影響。由圖6 可以看出,正反交果實(shí)總酚含量均呈正態(tài)分布,表現(xiàn)為連續(xù)變異。
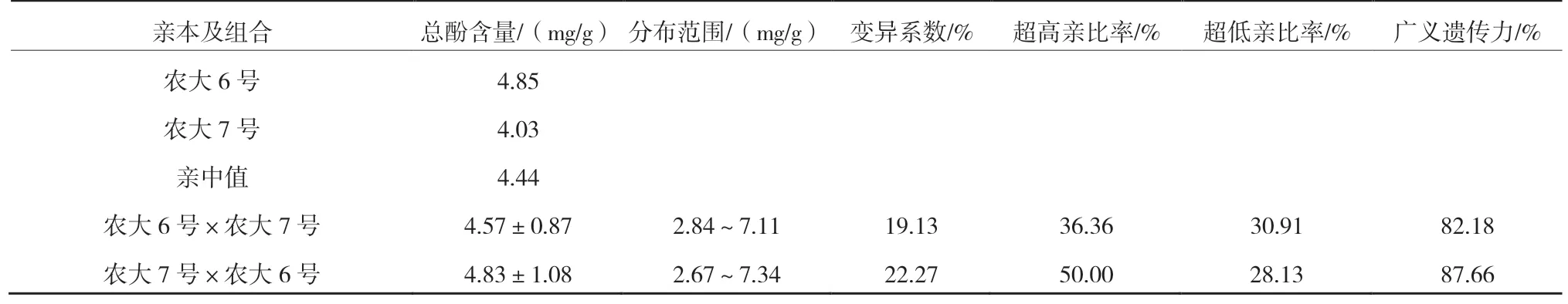
表6 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)總酚含量的遺傳變異
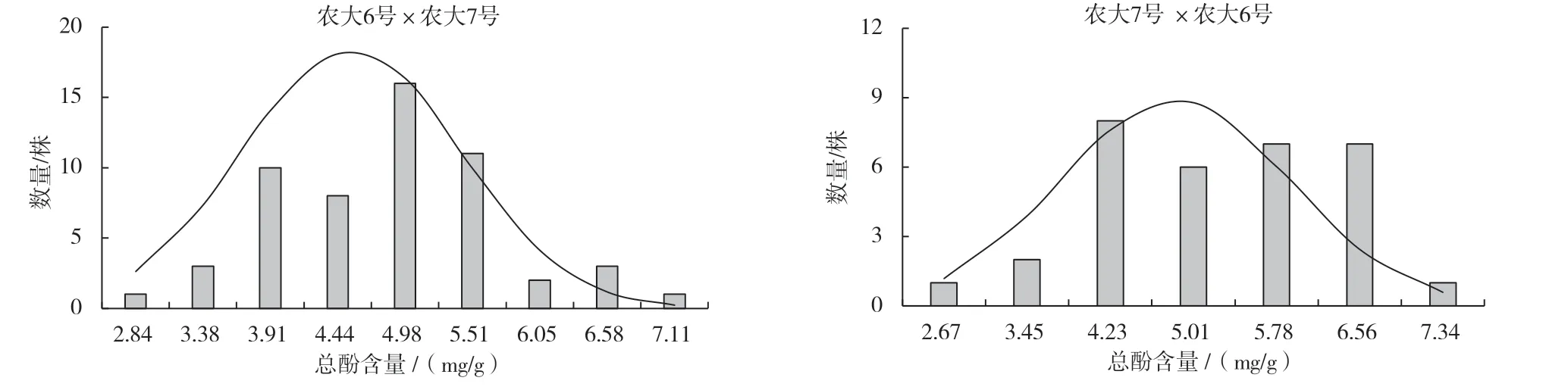
圖6 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)總酚含量的頻率分布
2.6 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)色澤的遺傳變異
由表7 可以看出,歐李后代果實(shí)色澤參數(shù)中只有‘農(nóng)大6 號’ב農(nóng)大7 號’的CI 平均值和‘農(nóng)大7 號’ב農(nóng)大6 號’的h°平均值大于親中值,其余所有的參數(shù)均小于親中值。
正反交所有色澤參數(shù)的變異系數(shù)均超過20%,說明均具有較大的選擇潛力,其中,尤其是b*和CI 這2 個(gè)參數(shù)的變異系數(shù)均超過了50%(表7),表明黃色歐李具有更大的選擇潛力。
廣義遺傳力正反交均是a*值較低(表7),說明紅色果實(shí)更易受到環(huán)境的影響。
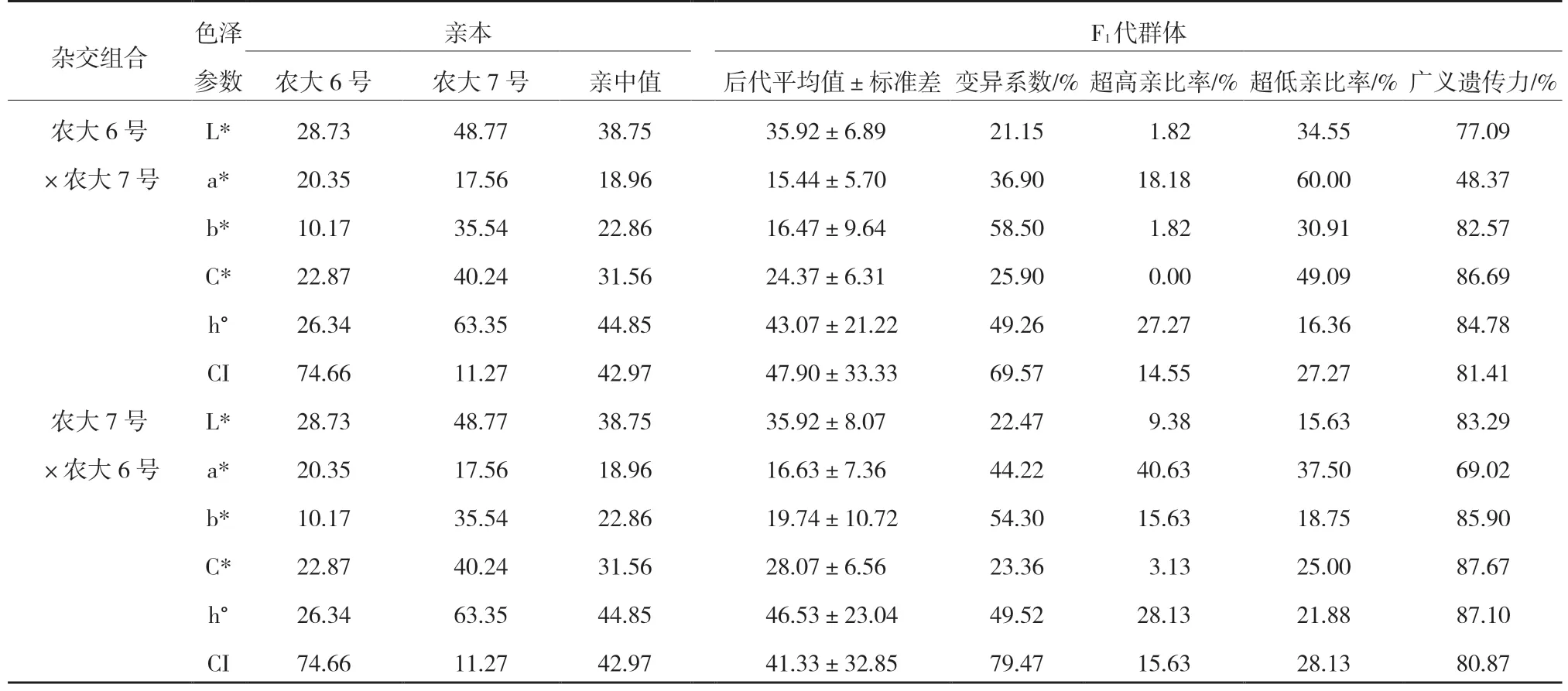
表7 ‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)色澤參數(shù)的遺傳變異
3 結(jié)論與討論
果實(shí)的大小及外觀形態(tài)是消費(fèi)者首先觀察到的外部特征,也是果品非常重要的商業(yè)指標(biāo),因此這也成為育種工作者關(guān)注的重點(diǎn)。本研究中‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)的單果重、縱徑、橫徑和果形指數(shù)均呈正態(tài)分布,且平均值均小于親中值,具有明顯的衰退變異現(xiàn)象。單果重后代變異系數(shù)較大,出現(xiàn)了廣泛分離,因此也出現(xiàn)了超高親的大果型單株。這與不同品種葡萄[11]、梨[13-14]、龍眼[15]和高州油茶果實(shí)[16]等雜交后代研究結(jié)果相一致。縱橫徑和果形指數(shù)的變異系數(shù)均較小,分離程度較低,縱徑未出現(xiàn)高親單株,選擇潛力較小。廣義遺傳力均較大,變異也主要來源于遺傳效應(yīng),不易受環(huán)境影響。
果實(shí)色澤也是消費(fèi)者評價(jià)果實(shí)品質(zhì)的常規(guī)指標(biāo),現(xiàn)已成為最重要的生理性狀和經(jīng)濟(jì)性狀。本研究選擇的是紅色和黃底紅暈2 個(gè)歐李品種作為正反交親本,采用CIE(國際照明委員會1976)L*a*b*色差體系[17]對歐李雜交后代果實(shí)色澤參數(shù)進(jìn)行評價(jià)。所測得色澤參數(shù)變異系數(shù)均較大,后代出現(xiàn)廣泛分離,選擇潛力較大。黃色比紅色的變異系數(shù)大,選擇空間更大,黃色比紅色的廣義遺傳力高,主要受遺傳效應(yīng)的影響,而紅色變化受環(huán)境影響較大。
類黃酮物質(zhì)廣泛分布于各種植物中,在生物的許多生理活動(dòng)中起著重要的作用。它也是人類飲食中來源最廣、含量最豐富的一類多酚化合物[18],也是非常重要的功能性物質(zhì)。本研究中,‘農(nóng)大6 號’與‘農(nóng)大7 號’歐李正反交后代果實(shí)的類黃酮和總酚含量均呈正態(tài)分布,雜交后代平均值均高于親中值,表現(xiàn)出一定的雜種優(yōu)勢。類黃酮和總酚含量較高的‘農(nóng)大6 號’作為父本時(shí),后代平均值較高且超高親比率也較高,更易篩選出高黃酮高多酚類型的歐李單株。后代變異系數(shù)較大,廣泛分離,具有較高的選擇潛力。廣義遺傳力較高,表明果實(shí)類黃酮和總酚含量主要受遺傳效應(yīng)的影響,但總酚遺傳效應(yīng)小于類黃酮,與類黃酮相比更易受到環(huán)境的影響。
本研究通過對‘農(nóng)大6 號’和‘農(nóng)大7 號’歐李及其正反交后代果實(shí)性狀測定后,得出測定的全部性狀均表現(xiàn)為連續(xù)變異,篩選出了超高親的大果型單株和功能性物質(zhì)含量較高的單株,同時(shí)得出以類黃酮和總酚含量較高的親本為父本時(shí),后代更易篩選出超高親單株,為后期選擇雜交親本提供了一定的參考依據(jù)。本研究只選擇了1 個(gè)雜交組合,具有一定的局限性,后期將選擇更多的雜交組合及后代植株,對后代遺傳規(guī)律做進(jìn)一步深入研究。
