MicroRNAs在肝纖維化進展中的作用
2021-02-07 06:09:06尤鴻美卜芳田孟宏武潘雪銀張亞飛
中國藥理學通報 2021年2期
關鍵詞:小鼠
尤鴻美,王 凌,卜芳田,孟宏武,潘雪銀,張亞飛,王 傲,黃 成,李 俊
(安徽醫科大學藥學院,炎癥免疫性疾病安徽省實驗室,安徽 合肥 230032)
1 研究背景
肝纖維化是指當肝臟受到持續性慢性刺激后,以Ⅰ型膠原占據主導的肝臟細胞外基質(extracellular matrix,ECM)過度沉積在肝實質,最終導致肝臟結構紊亂的可逆性瘢痕修復過程[1]。病毒性和自身免疫性肝炎、飲酒、非酒精性脂肪性肝炎,導致銅或鐵超載的代謝性疾病,毒素和膽道阻塞等是致使肝纖維化發生的常見病因。在肝臟中,活化的肝星狀細胞(hepatic stellate cells,HSCs)、門脈成纖維細胞、骨髓來源的肌成纖維細胞前體以及上皮間質轉化后的肝細胞等均是細胞外基質的生產細胞,其中活化的HSCs是生產細胞外基質的主要驅動力[2]。在正常生理條件下,HSCs處于靜息狀態,僅占肝臟總細胞數的5%-8%,存在于竇周間隙中并儲存大量脂滴和維生素A,而當肝臟受到持續性慢性刺激時,損傷的肝細胞和定居在肝臟內的枯否細胞釋放炎癥因子誘導HSCs活化并分化為α-SMA陽性的肌成纖維樣細胞。激活的HSCs會產生包括Ⅰ型和Ⅲ型膠原、纖連蛋白以及層黏連蛋白在內的細胞外基質成分,最終沉積在局部組織損傷部位。其中,竇周間隙過量的ECM會形成肝細胞與肝竇腔之間的一道物理屏障,導致肝細胞血流量減少,肝功能下降,進一步發展成為肝硬化,而目前肝臟移植手術是針對肝硬化的唯一治療手段,卻又局限于其并發癥以及供體的缺乏。因此,現階段,在肝纖維化尚未發展致肝硬化之前采取有效措施來逆轉肝纖維化是唯一可靠的治療方案。
MicroRNAs(miRNAs)是一類長度約22-25個核苷酸的單鏈RNA,由內源性發夾狀轉錄本產生[3]。MiRNAs主要通過靶向mRNA的3′-非編碼區(3′-UTR)來抑制mRNA翻譯或促進其降解從而發揮調控生理和發育過程等作用[4],因此其自身的表達必須要受到嚴格精密的調控才能保證正常的細胞功能。MiRNAs表達譜的變化對多個生物過程有較大影響,如細胞分化、凋亡、代謝、以及內分泌等。而且miRNAs的異常表達常伴隨有癌癥、心臟疾病、肝臟疾病、腎臟疾病等病理的過程發生及發展。因此,miRNAs作為預后指標以及診斷標志物的潛在作用已被越來越多的研究和認識。本篇綜述主要意在總結目前關于miRNAs調控HSCs活化與增殖在肝纖維化發生發展中的作用及其作為治療靶點和生物標志物的潛在作用。
2 肝纖維化的發病機理
肝纖維化是一種由各種損傷因素持續作用在肝臟所引發的可逆轉的瘢痕修復反應,其主要特征為ECM的過度沉積。如果患者沒有得到及時有效的治療,肝纖維化會進一步發展成為肝硬化,并伴隨有一系列并發癥,如肝功能喪失、門靜脈高壓、腹水、肝性腦病甚至肝細胞癌,因此肝纖維化已經成為全球重大衛生負擔之一[5]。當明確致病因素時,一些抗病毒藥物或免疫抑制劑可預防或減輕部分患者的肝纖維化,然而多數患者缺乏有效的治療方法,最終導致肝硬化以及預后不良。肝纖維化的進展與細胞外基質的數量和成分密切相關,在正常肝臟中,細胞外基質的產量在合成與降解之間有著精密的調節,處于高度動態平衡狀態,而在慢性肝損傷的過程中,合成的細胞外基質遠遠超過降解部分,并且其成分的改變也直接導致了肝纖維化的產生。健康肝臟中,ECM僅占肝臟組織切片相對面積的3%以及大約0.5%濕重,纖維化晚期階段的肝臟含有比正常健康肝臟多6倍的細胞外基質,主要成分包括Ⅰ型膠原、Ⅲ型膠原、Ⅳ型膠原、纖維連接蛋白、波狀蛋白、層粘連蛋白、彈性蛋白等。而增生性的肌成纖維細胞是過量ECM的主要細胞來源[6],例如,在肝、腎、肺等器官的慢性纖維增生性疾病中,肌成纖維細胞的存在是這類疾病重要的共同特征,它是一種收縮性極強的纖維母細胞樣細胞,通常由間充質細胞系轉分化而產生,這一過程成為“活化”。血小板衍生生長因子(PDGF),轉化生長因子β(transforming growth factor-β,TGF-β)等旁分泌因子和別的一些生長因子、細胞因子、趨化因子等均會激活細胞增殖、遷移、細胞外基質蛋白分泌的信號通路,促進間充質細胞向肌成纖維細胞的轉分化“活化”。已有研究發現,一些細胞群,如組織定居或骨髓來源的成纖維細胞、周細胞、上皮細胞等均可以轉分化為肌成纖維細胞,在四氯化碳(CCl4)誘導的肝纖維化及膽管結扎導致的小鼠肝纖維化模型中,82%-96%的肌成纖維細胞均來自于HSCs的活化[7],也就是說活化的HSCs是肝內ECM的主要生產細胞。因此抑制HSCs的活化并誘導凋亡是治療肝纖維化的關鍵[8]。20世紀70年代的早期臨床報告表明晚期的肝纖維化也是可逆的,之后的大量研究進一步證實了該結論,并且發現肝纖維化的消退伴隨有HSCs的凋亡、老化或轉分化為失活態以及細胞外基質的降解。綜上所述,在肝纖維化的治療過程中,將HSCs作為治療靶標,抑制其活化增殖,促進活化HSCs凋亡及衰老或誘導其轉化為失活的狀態來減少活化HSCs數量和ECMs的沉積量是治療肝纖維化的重要手段。
3 MiRNAs的生物合成與作用機制
MiRNAs 是一類內生的,由19-24個核苷酸組成的短序列非編碼RNA分子,保守性較高。MiRNA 的生物合成過程需要較多步驟的加工程序,首先RNA聚合酶Ⅱ在基因組的不同區域轉錄的初級轉錄物為幾百個核苷酸分子長度的原始miRNAs (pri-miRNAs),pri-miRNAs又在RNaseⅢ家族酶-Drosha的作用下被加工成70nt含發夾結構的前體miRNAs (pre-miRNAs),其中發夾結構的3′端向外突出兩個核苷酸,一個為5′磷酸,另一個為3′羥基,這是RNaseⅢ的經典產物,隨后輸出蛋白-5識別到發夾二核苷酸突出信號,并將pre-miRNAs轉運到細胞質。在細胞質中,pre-miRNAs的5′磷酸、3′突出以及莖環部位被RNaseⅢ家族酶-Dicer酶識別并結合,Dicer是一種“分子標尺”,可以將pre-miRNAs裂解為特定長度的雙鏈miRNA,其中一條鏈最終形成成熟的miRNAs,并與RNA誘導的沉默復合物(RNA-induced silencing complex,RISC)結合形成miRNAs核酸蛋白復合體發揮作用,另一條鏈則被降解。MiRNAs指導RISC于轉錄后水平下調目的基因的表達有兩種沉默機制:若mRNA與miRNA可以完全互補,那么該mRNA則會被RISC特異性降解掉;若兩者不能完全互補,那么mRNA就不會被降解,RISC則會阻止mRNA作為翻譯模板從而抑制蛋白質的生物合成[9]。
4 MiRNAs在肝纖維化中作用的最新研究
多年研究表明,大量miRNAs通過調節HSCs的活化,增殖,凋亡以及衰老等過程而參與肝纖維化的發生發展階段,因此通過調節miRNAs的表達水平進而調控HSCs的活化狀態最終實現對肝纖維化的治療作用將為肝纖維化的治療提供新的潛在治療方案。
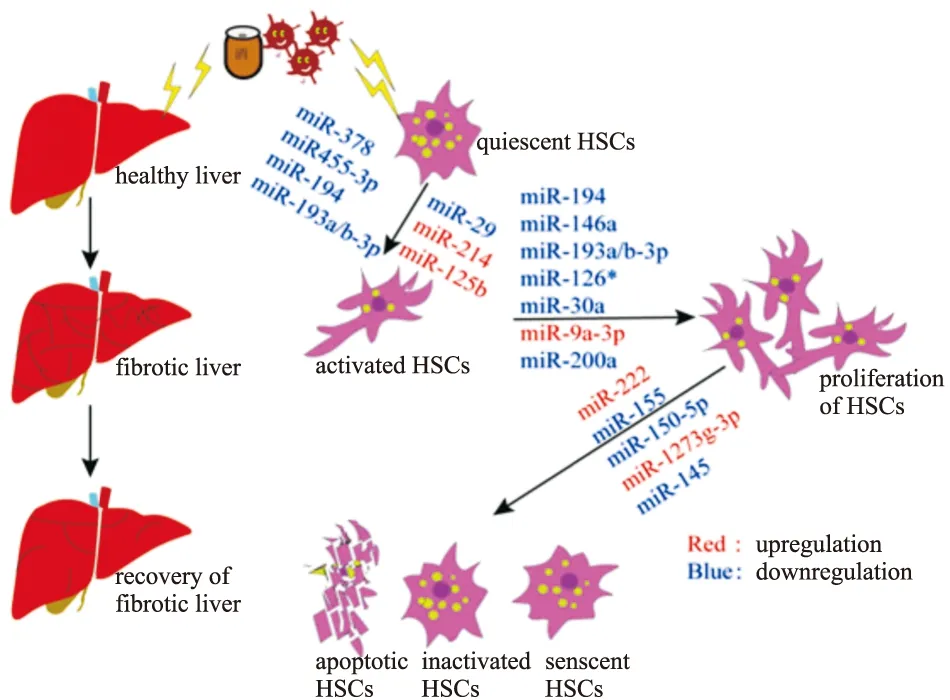
Fig 1 The regulatory effects of miRNAs on HSCs in progression and reversal of liver fibrosis
4.1 MiRNAs對HSCs活化及相關信號通路的調節正常的健康肝臟中,HSCs處于靜息狀態,當炎癥或慢性刺激損傷肝臟時,TGF-β及PDGF等炎性細胞因子會以自分泌或旁分泌的形式作用于HSCs,進而誘導其活化。而HSCs的活化是早期肝纖維化發生發展的中心事件,因此,抑制HSCs的活化有利于肝纖維化的逆轉恢復。近期,關于肝纖維化進展中HSCs與miRNAs之間的相互作用相繼被報道。
大量文獻已經報道miRNAs與HSCs的活化增殖及相關通路相關,Hyun等[10]研究發現miR-378家族(miR-378a-3p,miR-378b以及miR-378d等)在CCl4誘導的小鼠肝纖維化模型以及TGF-β誘導激活的HSCs中表達量顯著下降,并且能夠通過靶向轉錄因子Gli3抑制HSCs的活化,過表達miR-378a-3p可減少Gli3和其他促纖維化基因的含量但是HSCs失活指標膠質纖維酸性蛋白的表達顯著增加。Wei等[11]通過微陣列分析發現,miR-455-3p在TGF-β誘導的HSCs活化過程中表達下調,并且在CCl4,高脂喂養以及膽管結扎所致的3種小鼠肝纖維化模型中,miR-455-3p表達水平也明顯降低,進一步的功能學探究發現miR-455-3p可通過與熱休克蛋白1(Hsp1)的mRNA3′-UTR結合,抑制Hsp47/TGF-β/smad4信號通路最終抑制HSCs的活化。Wu等[12]報道miR-194在人肝纖維化組織及激活態HSCs中表達含量下降,高水平的miR-194可通過靶向蛋白激酶AKT2來減少HSCs中α-SMA、COL1α1表達從而致使HSCs失活,以及通過阻滯細胞周期從而抑制HSCs增殖,減緩肝纖維化進程。此外,研究者發現miR-193a/b-3p在con-A誘導的小鼠肝纖維化中表達下降,用慢病毒載體系統過表達miR-193a/b-3p后,小鼠肝纖維化癥狀明顯減輕[13]。MiR-29已被報道也是在肝纖維化中呈現下調狀態并具有抗纖維化功能的小RNA,TGF-β則能通過下調miR-29并激活HSCs以及促進ECM的沉積加重肝纖維化癥狀[14]。Kwiencinski等[15]發現在活化的HSCs中敲低miR-29可增加血小板衍生生長因子-B、胰島素樣生長因子-Ⅰ等促纖維化基因的表達。
近幾年,逐漸增多的研究表明,異常表達的miRNAs水平可能預示著肝纖維的不良預后。Ma等[16]通過對靜息態和活化態HSCs進行芯片分析,發現miR-214在活化HSCs中表達明顯上調,并且體外過表達miR-214后發現HSCs活化增多并且促纖維化基因和ECM等明顯增加,而miR-214發揮這一效應可能是通過抑制Hedgehog信號通路的負性調節因子Sufu的表達。為了深入探究其臨床功能,miR-214拮抗劑被注射進肝纖維化小鼠中,研究結果發現miR-214敲除組小鼠體內Sufu表達增加,肝纖維化癥狀也顯著減輕。You等[17]發現,miR-125b在肝纖維化進展期其含量在活化HSCs而非肝細胞中明顯上調,體內外抑制miR-125b后,導致HSCs活化被抑制,肝纖維化進程也被顯著延緩,深入進行機制研究發現,miR-125b可能是通過直接靶向Stard13來增強RhoA的活性,最終增加α-SMA表達水平和HSCs收縮性能。
4.2 MiRNAs對HSCs增殖及相關信號通路的調節研究發現HSCs增殖與miRNAs之間有著密不可分的關系,何等[18]報道miR-146a的表達在不同濃度TGF-β刺激的HSCs中呈現劑量依賴性下調,而當向HSCs中轉染miR-146a模擬物后,TGF-β誘導的HSCs的增殖則被明顯抑制。Du等[19]報道Wnt1和Wnt5a是miR-146a-5p的直接靶點,miR-146a-5p可能通過Wnt/β-catenin信號通路抑制HSCs增殖和膠原沉積等。Ju等[13]研究發現,在con-A誘導的肝纖維化模型中miR-193a/b-3p表達下調,相關的機制研究表明miR-193a/b-3p可以通過直接靶向CAPRIN1和TGF-β2,抑制細胞周期相關蛋白Cyclin D1及Cyclin E1的表達,從而抑制HSCs的增殖,肝纖維化癥狀減輕。MiR-126*在活化的HSCs中含量明顯下降,過表達miR-126*后,可明顯抑制TGF-β誘導的HSCs增殖并伴隨血管內皮因子的表達下調[20]。在CCl4誘導的大鼠肝纖維化模型中,miR-30a在原代HSCs中表達明顯下降,過表達miR-30a則可通過抑制上皮間質轉化過程,使HSCs活化和增殖減少,α-SMA、COL1α1表達下降。Qi等[21]報道,miR-9a-3p在HSCs中過表達可明顯降低其潛在靶基因Sirt1的表達,上調miR-9a-3p可促進HSCs的增殖,遷移以及活化。Sun等[22]報道在TGF-β1誘導的活化HSCs以及CCl4誘導的小鼠纖維化肝臟中,miR-200a表達水平下調。進一步的研究表明miR-200a,可通過靶向TGF-β2和β-catenin,來抑制TGF-β和wnt/β-catenin信號通路,從而抑制HSCs增殖,減緩肝纖維化進程。
4.3 MiRNAs對HSCs凋亡及相關信號通路的調節HSCs凋亡常常發生在肝纖維化逆轉階段,因此促進HSCs的凋亡將有助于肝纖維化病理的恢復。Ogawa等[23]報道miR-222在人類肝纖維化進程中表達量增加,并且在LX-2細胞中,miR-222的表達與α-SMA,COL1α1呈現相同趨勢,進一步研究發現,miR-222可以與細胞周期蛋白依賴性激酶抑制劑-1B(CDKN1B)的3′UTR區結合,抑制下游靶蛋白,促進表達NF-κB,最終抑制HSCs的凋亡。Dai等[24]報道miR-155在肝硬化患者的血清和肝臟組織以及活化HSCs中含量下調,在體外實驗中,增加miR-155含量后可顯著抑制上皮間質轉化過程及細胞外調節蛋白激酶1信號通路并誘導HSCs凋亡。Chen等[25]報道在肝纖維化進展中,miR-150-5p呈現出細胞特異性表達模式,即在肝細胞中表達升高,但是在HSCs中表達下降。過表達miR-150-5p可促進HSCs凋亡,而干擾素信號通路的介入可能是miR-150-5p誘導HSCs凋亡的機制之一。
在HSCs內促凋亡基因BAX和抗凋亡基因BCL2表達量的失衡,是致使HSCs凋亡的重要因素。Niu等[26]報道miR-1273g-3p在丙型肝炎病毒導致的肝纖維化病人的血清及肝組織中含量顯著升高,當向LX-2中轉染miR-1273g-3p模擬物時,細胞凋亡被明顯抑制,同時細胞增殖也明顯增加,而當miR-1273g-3p被抑制時,促凋亡基因BAX,BAD,cl-caspases3,cl-caspases9等表達水平升高,但是抗凋亡基因BCL-2表達水平下降,結果表明抑制miR-1273g-3p可促進HSCs凋亡進而阻止或逆轉肝纖維化進程。進一步的下游靶標探究發現miR-1273g-3p可能是通過PTEN/AKT信號通路來發揮抗纖維化作用,這一研究為肝纖維化的治療提供了新的途徑。
4.4 MiRNAs對HSCs衰老及相關信號通路的調節生物體內的多數細胞都要經歷分化、去分化、衰老、死亡等過程,其中衰老是多種刺激因素引起的細胞反應。當細胞進入衰老狀態時,其形態會發生較大變化,失去進入細胞周期的能力,并停滯再G0/G1期,增殖能力顯著降低,最終導致細胞永久性生長停滯[27],而當活化的HSCs停止增殖進入衰老狀態以后,ECM分泌明顯減少并且ECM降解酶的水平顯著增加。Kong等[28]研究表明,通過誘導HSCs衰老,肝纖維化病理進程明顯緩解。楊俊發等[29]在實驗中探究發現miR-145在小鼠肝纖維化組織以及活化的HSCs中表達均表現為下調趨勢,p53作為經典的細胞衰老調控基因,當其缺失時,HSCs衰老減少,最終導致肝纖維化惡化情況,而p53啟動子可以被ZEB1/2調節,因此楊等[29]研究發現miR-145可通過靶向ZEB2以p53依賴性方式促進活化的HSCs衰老,最終減輕肝纖維化進程,該發現可能為臨床肝纖維化的治療提供一種新的治療方案。
5 結論及展望
慢性病毒和代謝紊亂、酒精濫用等因素引起的進行性肝纖維化疾病平均每年可導致100萬人死于肝硬化,但針對抗纖維化的藥物迄今為止尚未被研究出世。活化的HSCs是分泌細胞外基質的肌成纖維細胞的主要細胞來源,而HSCs的活化、增殖和凋亡以及老化是肝纖維化發生和緩解的關鍵環節。因此,對HSCs進行細胞靶向藥物治療將成為治療肝纖維化的重要手段。大量miRNAs對HSCs的生物學功能調控的研究證實,部分miRNAs表達譜對HSCs的活化、增殖和凋亡及衰老等具有靶向調控作用,從而促進或減緩肝纖維化進程,表明miRNAs對于肝纖維化的預防、診斷、治療及預后等具有較大的潛力,并可能作為未來藥物治療的靶點,具有非常廣闊的研究前景。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34