
雙殼綱軟體動物含碳量及年度碳儲量估算
——以河北昌黎黃金海岸國家級自然保護區海域為例
2021-02-16 08:33:06楊悅王妍朱浩然劉憲斌
海洋通報 2021年6期
楊悅,王妍,朱浩然,劉憲斌
(1.天津科技大學,天津 300457;2.天津市海洋環境保護與修復技術工程中心,天津 300457;3.天津市綠色發展決策行為分析實驗室,天津 300457;4.海洋資源化學與食品技術教育部重點實驗室,天津 300457)
伴隨著全球經濟發展進程的加快,化石能源燃燒等人類活動加劇,造成了全球氣候變暖等一系列難題,這主要是由于原先全球各碳庫維系的平衡受到擾動并發生了變化。以CO2為首的主要溫室氣體濃度上升,對氣溫升高的貢獻率達到70%,同時也是導致全球變暖的主要因素(陳瀛洲等,2014)。因此,亟須控制CO2的排放量或者降低其在大氣中的含量,這個過程僅僅依靠陸地植物的光合作用和微生物的新陳代謝作用是遠遠不夠的,這就需要開發海洋中的“藍碳”(劉芳明等,2019)。海洋中的碳匯普遍存在于海洋生態系統中,是最為經濟且污染程度幾乎為零的固碳方式,十分契合當今的低碳發展理念(彭巨光等,2014)。海洋生態系統中浮游植物的固碳方式是通過光合作用將海水中溶解的無機碳轉化為有機碳(何培民等,2015),濾食性雙殼綱軟體動物通過攝食浮游植物或有機顆粒物將碳轉化為自身的軟體組織,并以鈣化的方式形成外殼部分,實現對碳的固定和封存(Chauvaud et al,2003)。同時,伴隨著雙殼綱軟體動物被捕獲,大量的碳能夠直接從海水中去除,脫離海洋碳循環,從而對海洋生物固碳產生重要的積極作用(于佐安等,2020)。目前對于雙殼綱軟體動物的含碳量計算主要有兩種方式,一是測定攝食碳、排泄碳、呼吸碳和排糞碳求得生物固碳,計算公式為:生物固碳=攝食碳-排泄碳-呼吸碳-排糞碳(方磊等,2011)。二是利用儀器分別測定軟體動物外殼與軟體組織的含碳量,進而通過經驗估算的方式獲得總體含碳量。Branes(2017)通過對極地海域浮游植物的每日監測情況,根據碳通量及生物碳泵求得了底棲動物整體的固碳量;Rueda 等(2005)基于雙殼綱軟體動物生物固碳原理,構建了碳分配模型,求得了在不同攝食率等條件下的固碳效率。第一種方法的計算結果雖然準確,但是卻需要大量的連續監測數據作為支撐,所花費的經濟與時間成本較高;而第二種方法能根據對不同年齡雙殼綱軟體動物的含碳量進行估算,對于大面積海域或者大量樣品的估算有較大優勢。基于第二種方法,Tang等(2011)首次估算了1999—2008 年中國海水養殖貝類和藻類的平均固碳量(3.76 伊106t)。此后有學者對中國沿海養殖貝類固碳量進行了估算,如劉慧等(2011)研究了黃海沿岸三省的養殖貝類碳儲量;梁君等(2015)調查了東極海洋牧場貝類筏式養殖區的碳匯能力。但是,對于海洋保護區雙殼綱軟體動物的含碳量目前還鮮有報道。
昌黎黃金海岸國家級自然保護區(以下簡稱昌黎自然保護區)位于河北省秦皇島市昌黎縣,總面積208.5 km2,其中核心區70.50 km2,緩沖區138 km2。該保護區也是國家二級保護動物青島文昌魚的重要棲息地,水質與沉積環境良好。本文基于2019 年海域生態環境調查結果,對昌黎自然保護區雙殼綱軟體動物的含碳量進行了評估,并對該保護區碳匯能力及前景進行了初步探討,以期為提高海洋碳匯潛力、探索海洋“藍碳”遷移轉化途徑以及其他海域保護區碳儲量估算提供參考依據。
1 材料與方法
1.1 數據來源與方法
于2019 年春、夏兩季分別對昌黎自然保護區海域進行兩個航次的海域生態環境監測,共設置14 個監測站位(圖1)。樣品采集過程嚴格按照《海洋調查規范》 (GB/T 12763—2007)執行。使用0.09 m2的抓斗式采泥器采集大型底棲動物樣品,每個監測站位采集4 個平行樣品。樣品用5%甲醛固定,帶回實驗室進行鑒定、稱量、計數等。
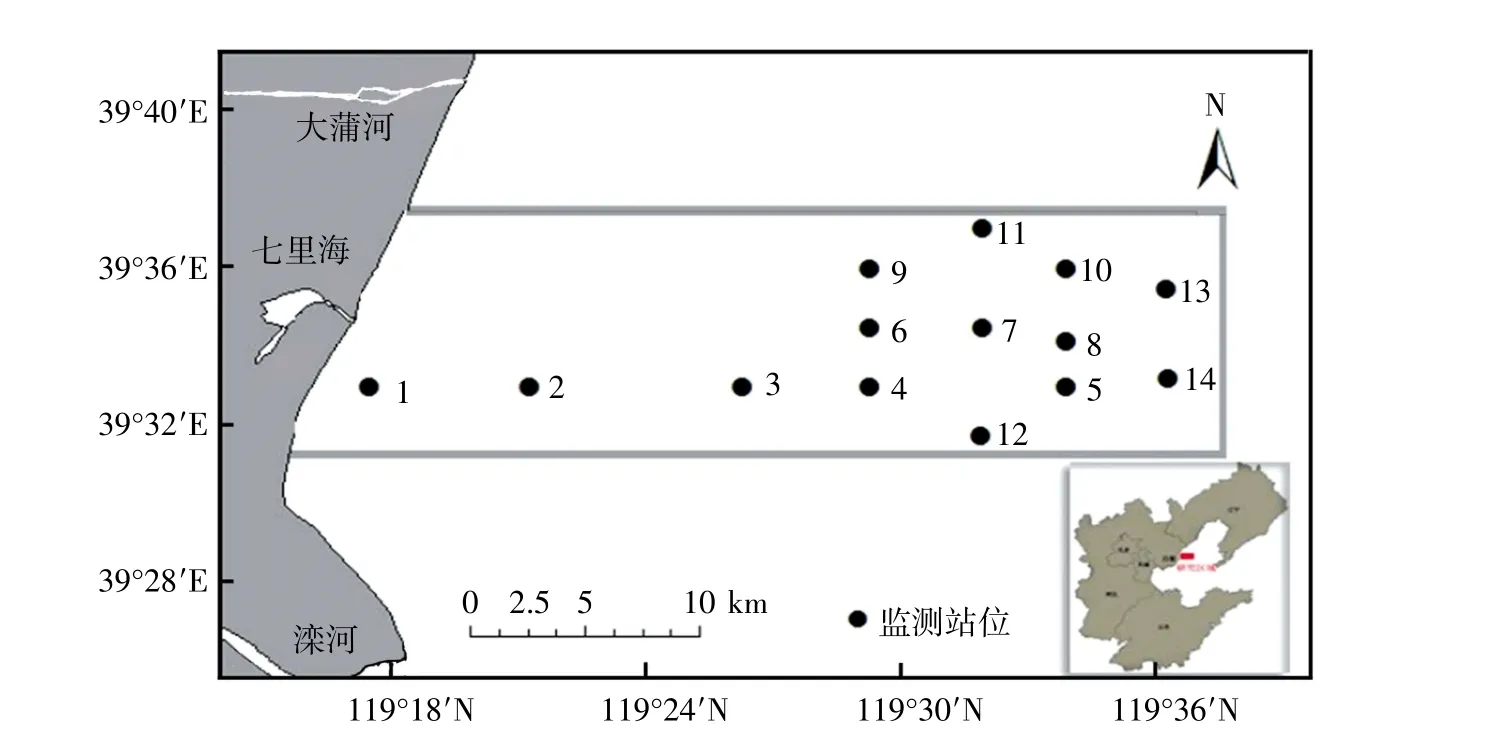
圖1 昌黎自然保護區海域監測站位示意圖
1.2. 雙殼綱軟體動物含碳量估算方法
不同種類的雙殼綱軟體動物,其含碳量也有所不同,根據文獻(Hawkins et al,1985)一般采用以下方法進行估算。
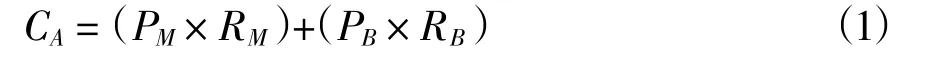
式中,CA代表雙殼綱軟體動物總含碳量;PM代表軟體部分產量;RM代表軟體部分含碳率;PB代表外殼部分產量;RB代表外殼部分含碳率;不同物種的干濕比和軟體部分與外殼部分含碳量不同(徐海龍等,2016)。本文選取干濕比和含碳量的平均值,同時按照雙殼綱軟體動物濕重的1/5 折算為干重(于子山等,2001),或按照已經報道的同科的生物干濕比進行轉換。軟體部分干重占全部濕重的4.31%,外殼干重占全部濕重的60.20%。含碳量按照軟體部分占個體總干重的42.21%~45.98%折算,外殼部分占個體總干重的11.44% ~ 12.01%折算(李昂等,2013)。
2 結果
2.1. 昌黎自然保護區海域雙殼綱軟體動物含碳量估算
在2019 年春、夏兩季的調查中共采集到雙殼綱軟體動物9 種,屬于4 目7 科(表1),其攝食方式均為濾食性。調查結果表明,春、夏兩季平均棲息密度為23.31 依5.46 ind./m2。基于對該區域監測的連續性,2017 年和2018 年春、夏兩季雙殼綱軟體動物平均棲息密度分為33.33 依2.02 ind./m2和25.92 依2.69 ind./m2。可見該區域的雙殼綱軟體動物棲息密度年際變化較小,維持在30 ind./m2左右。同時根據漁業資源利用與補充理論模式方程:當(砸垣鄖)躍(C+M)時,資源補充量有剩余;當(砸垣鄖)躍(C+M)時,資源補充量與捕撈和死亡持平。式中R 為補充群體初期生長量,G 為中后期生長量,C為捕撈力量,M 為自然死亡量(吳耀泉 等,1998)。由于研究區域為國家級自然保護區,所以捕撈等人為活動幾乎為零,因此從連續的調查結果來看,應為資源補充量和死亡持平狀態,所以雙殼綱軟體動物的年際含碳量變化可以忽略。
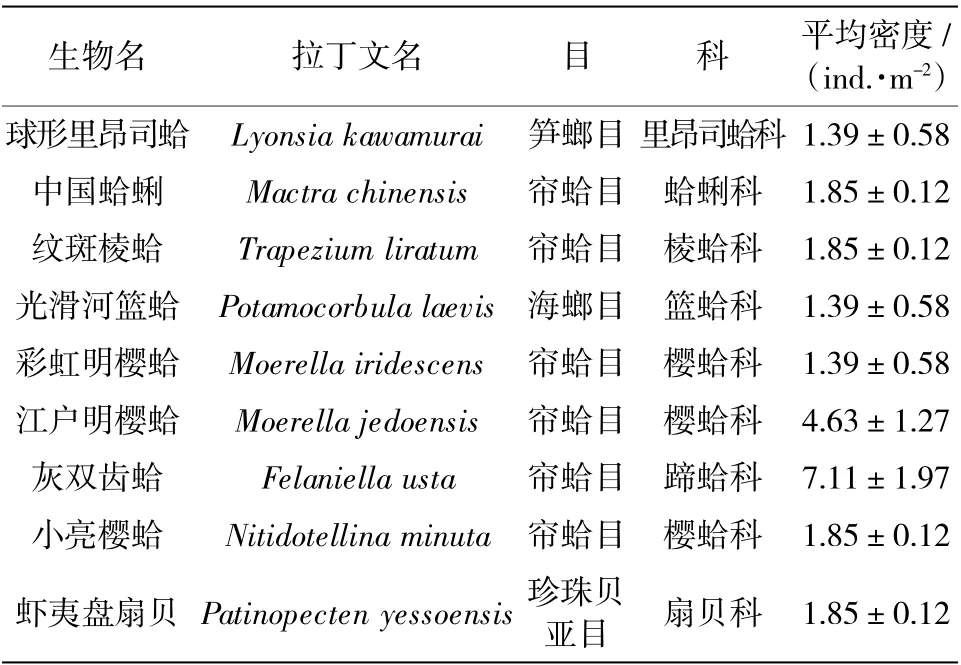
表1 昌黎自然保護區2019 年雙殼綱軟體動物平均密度
依照昌黎自然保護區海域面積(208.5 km2)來估算全部區域雙殼綱軟體動物的碳含量(表2)。結果表明,2019 年昌黎自然保護區雙殼綱軟體動物碳含量約為391.79~414.54 t,其中蝦夷盤扇貝碳含量占比最大,為71.50%。有資料表明,雙殼綱軟體動物每固定1 g 碳吸收的CO2量約為3.67 g(趙述華等,2019),故2019 年昌黎自然保護區海域雙殼綱軟體動物吸收的CO2總量為1436.57 ~1520.00 t。
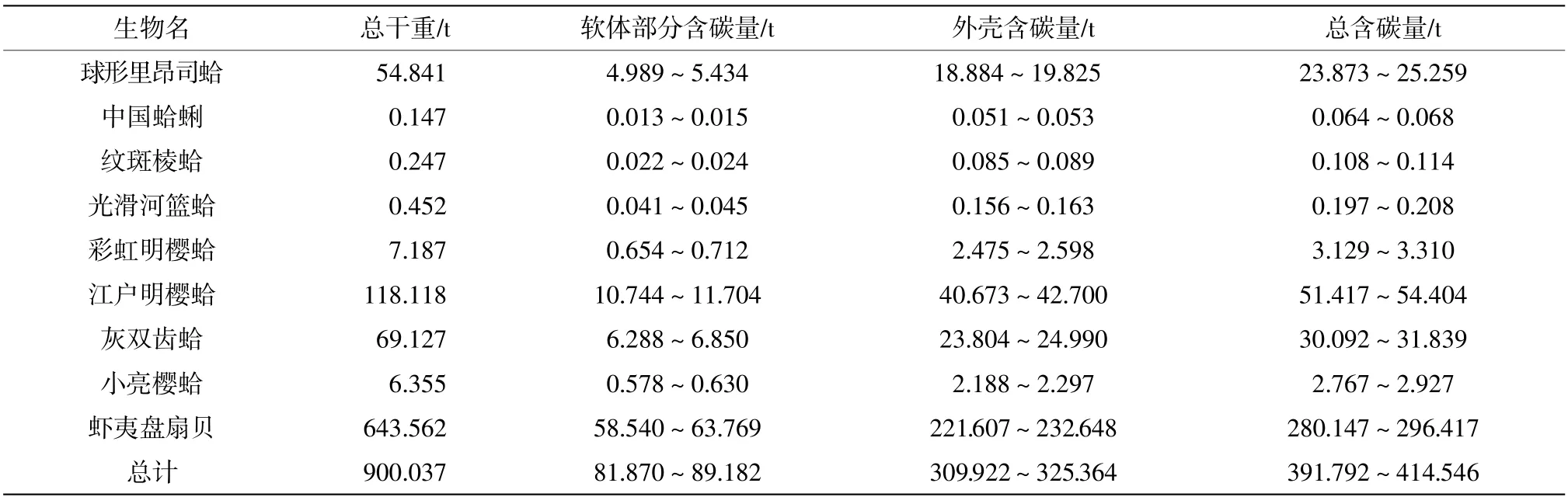
表2 昌黎自然保護區2019 年雙殼綱軟體動物含碳量
2.2. 昌黎自然保護區海域雙殼綱軟體動物碳匯經濟價值評估
海洋碳匯價值可理解為“吸收、固定、儲存CO2的海洋生物、非生物和其他海洋活動的總價值” (劉芳明等,2019)。按照海洋碳匯“五分類”模型,海洋碳匯總經濟價值應包含使用價值(直接使用價值、間接使用價值、選擇價值)和非使用價值(遺產價值、存在價值)。在本研究中主要考慮間接使用價值,即氣候調節價值(碳稅法與人工造林法)。
(1) 碳稅法。排放單位CO2征收的稅額作為CO2的價格,計算公式為:

Q 代表海洋生物固定CO2的物理量;r 為核算期半年內的碳稅平均值。
(2) 人工造林法。人工造林吸收單位CO2所耗費的成本,計算公式為:

Q 代表海洋生物固定CO2的物理量;C 代表人工造林吸收單位CO2所消耗成本。
為了使核算更加精確,取這兩種方法核算價格的平均值,作為最終海洋碳匯的氣候調節價值。
值得說明的是,在碳匯經濟價值核算中還應該考慮食用價值,即可食用海洋生物的年產量按照該生物市場批發價計算金額。但該區域屬于保護區,禁止人工捕撈活動,所以此項可忽略不計。
根據表3、表4 中的數據,可以推算出2019年昌黎自然保護區雙殼綱軟體動物實現的碳匯量相當于21.80~23.07 ha 的天然林地或23.47~24.83 ha的人工林地。同時,結合2019 年碳交易價格(寸永護等,2020),按照碳稅法和人工造林法來核算2019 年該區域雙殼綱軟體動物在氣候調節方面產生的碳匯經濟價值為146 677.6~155 179.6 元/a。
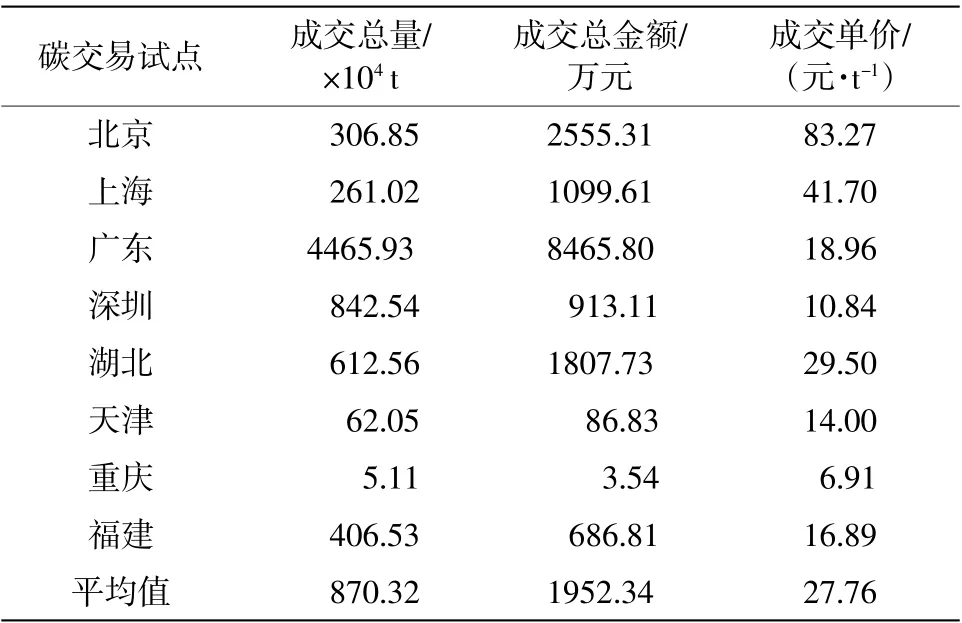
表3 2019 年全國碳排放交易試點成交情況

表4 天然林地與人工林地碳匯情況
3 討論
3.1 昌黎自然保護區海域與其他海域碳儲量比較
目前報道的軟體動物含碳量資料主要以海水養殖區域的貝類為主要研究對象,且在海水養殖的范疇中貝類除包括雙殼綱軟體動物外,還包括腹足綱軟體動物,如脈紅螺(Rapana venosa)、香螺(Neptunea cumingi Crosse) 等。根據資料報道,2002年全國海水養殖貝類的含碳量約為8.57 伊105t(方磊等,2011)。深圳近岸海域2014—2018 年貝類產量均值為1800 t/a,固碳量約為477.22 t/a(趙述華等,2019)。河北省2010 年海水養殖的貝類總含碳量約為2.74伊104t(李昂等,2013),約是本文核算的66 倍。溫州市海水養殖貝類2004—2014年年均固碳為5732.57 t(柯愛英等,2016),同樣遠超于本文的核算結果。其主要原因是資料中貝類均處于養殖區,產量大,且包括腹足綱軟體動物,而昌黎自然保護區海域的主要優勢種是青島文昌魚,雙殼綱軟體動物棲息密度較低,且在估算中沒有包括腹足綱軟體動物。雖然昌黎自然保護區海域周邊遍布貝類養殖區,但貝類移動范圍小,且該保護區地處近岸海域,無大型環流作用,因此外部輸入的貝類規模微乎其微,因而導致估算結果相比其他報道較低。另外在調查中發現的幾種雙殼綱軟體動物除中國蛤蜊、蝦夷盤扇貝外均為體積較小的物種,且中國蛤蜊與蝦夷盤扇貝均為幼體,導致估算含碳量時占比較小(Zhang et al,2005)。除此之外,在2017—2019 年的調查中,雙殼綱軟體動物的平均生物量均小于50 ind./m2,也反映出該區域雙殼綱軟體動物的存量較少(郭興然等,2019),導致其含碳量較低。在碳源的貢獻方面,硅藻是浮游植物中固碳能力較強的生物(孫軍,2011),且硅藻是該海域的主要優勢種,豐度最高值可以達到1.3伊107cells/m3(孫富強 等,2017),有效保障了碳源的供給,也說明了研究區域雙殼綱軟體動物棲息密度較小是導致含碳量較低的主要原因。另一方面,按照生態系統能量流動原理,雙殼綱浮游植物能直接獲取硅藻等浮游植物的能量,在整個生態系統中獲得的碳源較多,因此在碳儲量方面具有優勢。
3.2 雙殼綱軟體動物碳匯能力提升途徑及前景
雙殼綱軟體動物碳匯的途徑主要有三方面:一是鈣化作用(Calcification),利用海水中的碳酸氫根離子(HCO3-) 和鈣離子(Ca2+) 形成碳酸鈣(CaCO3);二是生物同化作用(Assimilation),通過濾食海水的有機顆粒物和浮游植物,把相當部分的有機碳同化到其軟體組織中(Yang et al,2005);三是生物沉積作用(Biodeposition),通過其自有的生活方式,將海水中的懸浮顆粒物以假糞便的形式輸入水體的底層,加快生物沉積過程,把部分有機碳或者無機碳封存在沉積物中(沈新強 等,2011)。從雙殼綱軟體動物的碳匯途徑可以看出,加快其攝食率可以提升其固碳量。目前,已經有許多學者研究了溫度、鹽度、pH 對于雙殼綱軟體動物攝食率的影響。資料表明,鹽度為20,pH=8.7時,文蛤等雙殼綱軟體動物的濾水率和攝食率最大(楊杰青 等,2016);水溫在16~28 益時濾水率和攝食率隨溫度升高而逐漸升高,同時體積質量與其濾水率和攝食率呈正相關關系(黃洋等,2013)。但在實際的海水中,pH 與鹽度在小范圍內變化不大,溫度則是隨季節變化和水深變化而變化,因此,需要在雙殼綱軟體動物濾水率和攝食率較高的夏季保證海水中浮游植物和有機顆粒物能充分滿足其需求,從而提高其固碳能力。
雙殼綱軟體動物在死亡或被其他生物捕食后,由于其特殊的結構,其外殼會沉降到海底或隨著潮汐作用被搬運到沙灘上,但外殼部分所固定的碳則不會很快進入地球化學循環中。據此可推測該區域的碳儲量要遠遠高于某一時期或現存的雙殼綱軟體動物含碳量,主要是因為在該海域的底部有著大量的雙殼綱軟體動物外殼。同時,雙殼綱軟體動物不僅可以有效地凈化水體,而且也是人們喜食的主要海產品,具有十分廣闊的生態環境價值和食用價值。它們一般生活在海底沉積物中,其棲埋深度一般與水體溫度有關,溫度高,棲埋淺;溫度低,棲埋深(Zhang et al,2009)。雙殼綱軟體動物很少移動,特別是幼苗基本不會主動移動,因此,對于定量估算某個海區的產量比較容易。同時,由于其有較強的固碳能力,能夠通過捕撈等方式較為直觀地“看到”其固碳量,因此,雙殼綱軟體動物在碳匯以及碳交易中占據著一定地位。
4 展望與不確定性分析
海洋系統是地球上最大的生態系統,其生物群落與海洋環境共同形成了地球上最大的碳匯——“藍碳”。有資料表明,藍碳的總碳匯量高達3.9 萬億噸,占全球總碳匯量的93%,約是大氣的50 多倍,人類活動產生的CO2中大約有30%被海洋所吸收(Chen et al,2009)。作為一個負責任的大國,中國已經向世界宣布要在2030 年達到CO2排放峰值,2060 年時實現“碳中和”。實現這一目標就必須要充分利用我國300 多萬平方千米的海洋國土(徐敬俊等,2018)。在海洋中,從初級生產者到頂級消費者的復雜食物網,使得海洋中碳的遷移、轉化途徑異常復雜。但食物網的每一層級中都存在有較強固碳能力的物種。例如,德國萊布尼茨海洋學研究所發現,大型底棲動物棘皮動物門中海星、海膽、蛇尾等,能夠直接從海水中吸收碳形成外骨骼,將碳封存在骨骼中,死后封存在碳骨骼中的碳會沉降到海底,脫離海洋地球碳循環,具有較強的碳匯作用(Giles et al,2006)。因此,碳在海洋生態系統的遷移轉化將成為今后的熱門研究方向,從而能夠利用特殊層級的物種有針對性地提升碳在海洋生態系統中的固定效果。同時,建立海洋生態系統的碳匯模型也是十分必要的。在獲得大量數據的基礎上,通過考慮海洋環流、水質、周邊環境等影響生物固碳的因素,進而擬合碳匯模型,既可以達到自然條件下預測碳儲量的目的,又可以根據預測結果進行人工干預從而獲得指定數量的碳匯。當然,這還需要進行大量的研究工作。
本研究在樣品采集過程中雖然嚴格按照國家標準并在專家的指導下進行,但是仍存在不確定性因素。如本研究在雙殼綱軟體動物含碳量的估算中,由于監測站位較少,且缺乏冬季與秋季的采樣結果,會導致結果出現偏差;另外,雖然在樣品采集時設置了平行樣品,但在分析中缺少時間上的縱向結果對比,也可能會產生誤差;同時,在估算模型中沒有考慮到雙殼綱軟體動物糞便沉降所固定的碳。因此,在今后的調查研究中,應設置更多的監測站位,并且在季節上要有連續性,從而保證結果的準確性。
致謝:感謝昌黎黃金海岸國家級自然保護區管理處對樣品采集提供的幫助,感謝課題組同學及老師對數據處理與資料查找提供的幫助。
