分泌型煙草腺毛形態和葉面化學成分的比較
2021-02-25 07:47:32馬旭東王召軍閆筱筱張洪映
煙草科技 2021年1期
關鍵詞:煙草
馬旭東,王召軍,閆筱筱,崔 紅,張洪映
河南農業大學煙草學院,鄭州市文化路95號 450002
植物表皮毛是表皮細胞的特化結構,是植物表皮細胞的延伸,廣泛存在于植物的地上部分。植物表皮毛作為植物與環境間的天然屏障,能有效減輕病原菌、昆蟲等有害生物對植物的損害以及紫外線照射造成的損傷,有利于減少水分散失和熱量損耗,提高植物對自身溫度的調節能力[1-3]。擬南芥的表皮毛是一種特化的、無腺體的單細胞表皮毛[4-5]。與擬南芥不同的茄科、豆科、菊科、大麻科和唇形科等植物屬于多細胞表皮毛,這些物種的表皮毛類型豐富,根據其分泌腺的有無,可分為保護毛(非分泌型)和腺毛(分泌型)。如菊科蒿屬的黃花蒿表面有兩種表皮毛,包括腺毛和T型保護毛。青蒿素是黃花蒿在腺毛中合成、積累和分泌的一種倍半萜內酯,以青蒿素為基礎的聯合用藥(ACTs)被認為是治療瘧疾的一線藥物[6]。番茄屬植物表皮毛共有7種類型(Ⅰ~Ⅶ),其中Ⅰ、Ⅳ、Ⅵ和Ⅶ型屬于腺毛,Ⅱ、Ⅲ和Ⅴ型為保護毛[7-8]。茄科煙草屬植物的表皮毛發達,包括保護毛和腺毛,腺毛約占總表皮毛的85%,根據腺毛柄部長度,又分為長柄腺毛和短柄腺毛;煙草腺毛分泌物中不僅含有西柏烷二萜等重要的香氣前體物質,還具有抵抗蚜蟲的作用[9]。有研究表明,煙草分泌物西柏三烯二醇有抗癌作用,且可誘導愛潑斯坦-巴爾病毒早期抗原和蛋白質磷酸化[10]。煙草長柄腺毛中可產生含Cd/Ca的晶體并外溢,進而主動排除煙葉中的Cd[11-12]。短柄腺毛是葉面抗性蛋白的主要合成場所,在抑制孢子萌發、抵抗真菌侵染中具有重要作用[13]。而目前有關不同類型煙草腺毛分泌物的差異方面鮮見報道,為此,以不同腺毛類型的煙草品系TI1068和TI35為材料,分析其表皮毛的形態結構、主要葉面化學成分以及蚜蟲嗜好性,旨在揭示煙草腺毛分泌物的化學成分特點和功能,為煙草腺毛的改良提供依據。
1 材料與方法
1.1 材料
普通煙草TI1068和TI35是由美國農業部牛津煙草研究所(Oxford Tobacco Research Station,Oxford,North Carolina,USA)提供的,且是從美國本土以外收集到的普通煙草(N.tabacum)栽培品系。
1.2 煙草葉片表皮毛的形態比較
在煙株成熟期,選擇長勢一致的4棵煙株,選取葉長約10 cm的頂葉,在質量體積分數為0.2%的羅丹明B水溶液中浸染30 min,后用蒸餾水漂洗3次,沖洗未結合的染料,吸干表面水分。避開葉脈切取葉中部約5 mm×8 mm葉片,在超景深顯微鏡(VHX-5000,日本基恩士公司)下觀察葉片的表皮毛形態和數量,隨機選擇10個視野進行腺毛密度統計和分析。
1.3 葉面化學成分分析
在煙株成熟期,選擇長勢一致的4棵煙株,選取第10~12位(從下向上數)中部葉提取葉面化學物質。避開主脈位置,取直徑為10 cm的葉圓片20片,采用有機溶劑萃取的方法提取煙葉的葉面化學物質[14]。將葉圓片在二氯甲烷中依次浸提2 s,重復8次后加入1 mL內標(2.020 mg/mL蔗糖八乙酸酯和2.474 mg/mL正十七烷醇的混合液),旋轉蒸發儀濃縮,氮吹儀吹干。加入500 μL體積比為1 ∶1的N,N-二甲基甲酰胺(DMF)和N,O-雙三甲硅基三氟乙酰胺(BSTFA),在75 ℃水浴中進行60 min衍生化反應,加入N,O-雙乙酰胺和吡啶各125 μL,獲得待測液。利用GC/MS(色譜儀型號HP-5890,質譜儀型號vc-70SE,美國Agilent公司)進行化學成分的定性和定量分析,色譜參數條件參考文獻[15]的方法。
1.4 半定量RT-PCR分析
在煙株成熟期,選擇長勢一致的4株煙,取長約10 cm的頂葉,去除葉脈后混合煙樣。按照Invitrogen Trizol Reagent說明書提取總RNA,然后反轉錄合成cDNA。以L25作為內參基因,采用半定量RT-PCR方法分析西柏烷二萜合成基因GGPPS、CBTS和CYP71D16的表達量。在NCBI的Gene數據庫(http://www.ncbi.nlm.nih.gov)檢索GGPPS、CBTS和CYP71D16的序列。將獲得的每個基因家族序列進行序列比對,利用Primer 5.0在基因保守域設計引物,GGPPS-F:5′-TGTCGATTC ATGGGCTGCT-3′,GGPPS-R:5′-GTGGATCACAGG TGGGTCTTT-3′;CBTS-F:5′-CGACTTGCGAGGC AACAAGG-3′,CBTS-R:5′-GAGCGAATTCCATG TTCAATGAC-3′;CYP71D16-F:5′-TGTACTGCTA AGACTTATGA-3′,CYP71D16-R:5′-GGTTTCGA GTTCGTCCATT-3′;L25-F:5′-CCCCTCACCACA GAGTCTGC-3′,L25-R:5′-TTCTAACTCCTGTTGTT GTGGGAA-3′。PCR反應條件:95 ℃5 min;95 ℃40 s,60 ℃50 s,72 ℃30 s,28個循環;72 ℃5 min。
1.5 蚜蟲嗜好性比較
取TI1068和TI35大小相近的幼葉(15~20 cm)置于同一培養皿(直徑25 cm)中,為避免葉片在試驗過程中失水,用濕潤的脫脂棉將葉柄包裹,培養皿底部放置6層浸透無菌水的濾紙。將100頭蚜蟲等距放于葉片之間,不限制蚜蟲的自由活動。將培養皿放置于25 ℃恒溫人工氣候箱(RXZ-500,寧波江南儀器廠)12 h,每隔3 h統計葉片上的蚜蟲數量,設置3次重復,計算平均值。
2 結果與討論
2.1 煙草葉片表皮毛的形態比較
煙草TI1068和TI35葉片表皮毛的形態學觀察結果見圖1。TI1068和TI35同時具有長柄腺毛、短柄腺毛和保護毛3種類型表皮毛。TI1068的腺毛以長柄腺毛為主,腺毛頭部膨大,由多細胞組成,整個腺頭被分泌物包裹成球形或水滴形,含有極少量的保護毛(圖1A,B)。TI35的表皮毛以保護毛為主,頂部細胞與柄細胞結構無差異,TI35的長柄腺毛量甚少(圖1C,D)。TI1068和TI35的短柄腺毛形態無差異,柄細胞均為單細胞,腺毛頭部膨大呈圓球形,分泌物不及長柄腺毛多。因此,TI1068的腺毛分泌物遠多于TI35。
羅丹明B水溶液染色試驗結果(圖2)顯示,保護毛不能著色;長柄腺毛和短柄腺毛可以被羅丹明B水溶液染紅;總體來看,長柄腺毛的著色程度比短柄腺毛深。可見,長柄腺毛和短柄腺毛均有糖酯分泌,而長柄腺毛的糖酯含量或種類多于短柄腺毛。顯微鏡下統計葉面表皮毛的密度發現,TI1068總腺毛密度高于TI35,但未達到顯著水平;TI1068的長柄腺毛密度極顯著高于TI35,約為TI35的10倍;TI35保護毛密度極顯著高于TI1068,約為TI1068的7倍;TI1068和TI35的短柄腺毛數量無顯著差異。
2.2 煙草葉面化學成分分析

圖1 煙草TI1068和TI35葉片表皮毛的形態比較Fig.1 Morphological comparison of trichomes on leaf surface between TI1068 and TI35
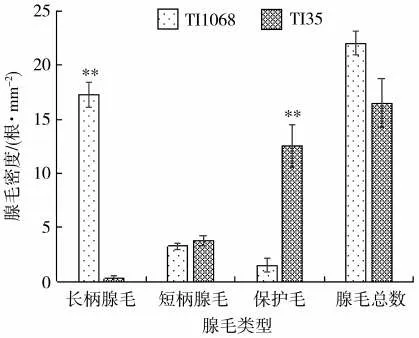
圖2 煙草TI1068和TI35葉片表皮毛密度的比較Fig.2 Comparison of trichome densities on leaf surface between TI1068 and TI35
西柏烷二萜、烷烴和蔗糖酯是煙草葉面分泌物的主要化學成分[16-17]。葉面化學成分分析結果(表1)顯示,TI1068和TI35的葉面分泌物成分相似,均含有西柏烷二萜、烷烴和蔗糖酯。TI1068中3種成分總量約是TI35的10倍,而各成分含量存在差異。TI1068:西柏烷二萜(66%)>蔗糖酯類(24%)>烷烴類(10%);TI35:烷烴類(94%)>西柏烷二萜(5%)>蔗糖酯類(1%)。
通過對TI1068和TI35葉面化學成分的比較發現,TI1068葉面成分主要是西柏烷二萜和蔗糖酯,而TI35主要是烷烴類,僅存在微量的西柏烷二萜和蔗糖酯Ⅳ。此外,TI1068和TI35的烷烴種類和總量無差異。考慮到TI1068和TI35的表皮毛類型差異,即TI1068以長柄腺毛為主,TI35以保護毛為主,短柄腺毛數量無差異,因此推測長柄腺毛是西柏烷二萜和蔗糖酯合成的主要場所,而烷烴類主要在短柄腺毛合成。腺毛分泌物對于煙草品質和抗性有重要影響[18]。有研究發現噴施茉莉酸甲酯可以促進長柄腺毛的發生,增加葉面分泌物含量,進而增強葉片對蚜蟲的驅避性[19-20];敲除煙草NtCycB2基因可以增加長柄腺毛的密度,提高西柏烷二萜含量,進而改善煙葉品質和抗性[21-22]。這些研究結果表明,不同腺毛類型對腺毛分泌物的貢獻不同,長柄腺毛是西柏烷二萜合成的主要場所,增加長柄腺毛密度是提高煙葉品質和抗性的重要途徑。
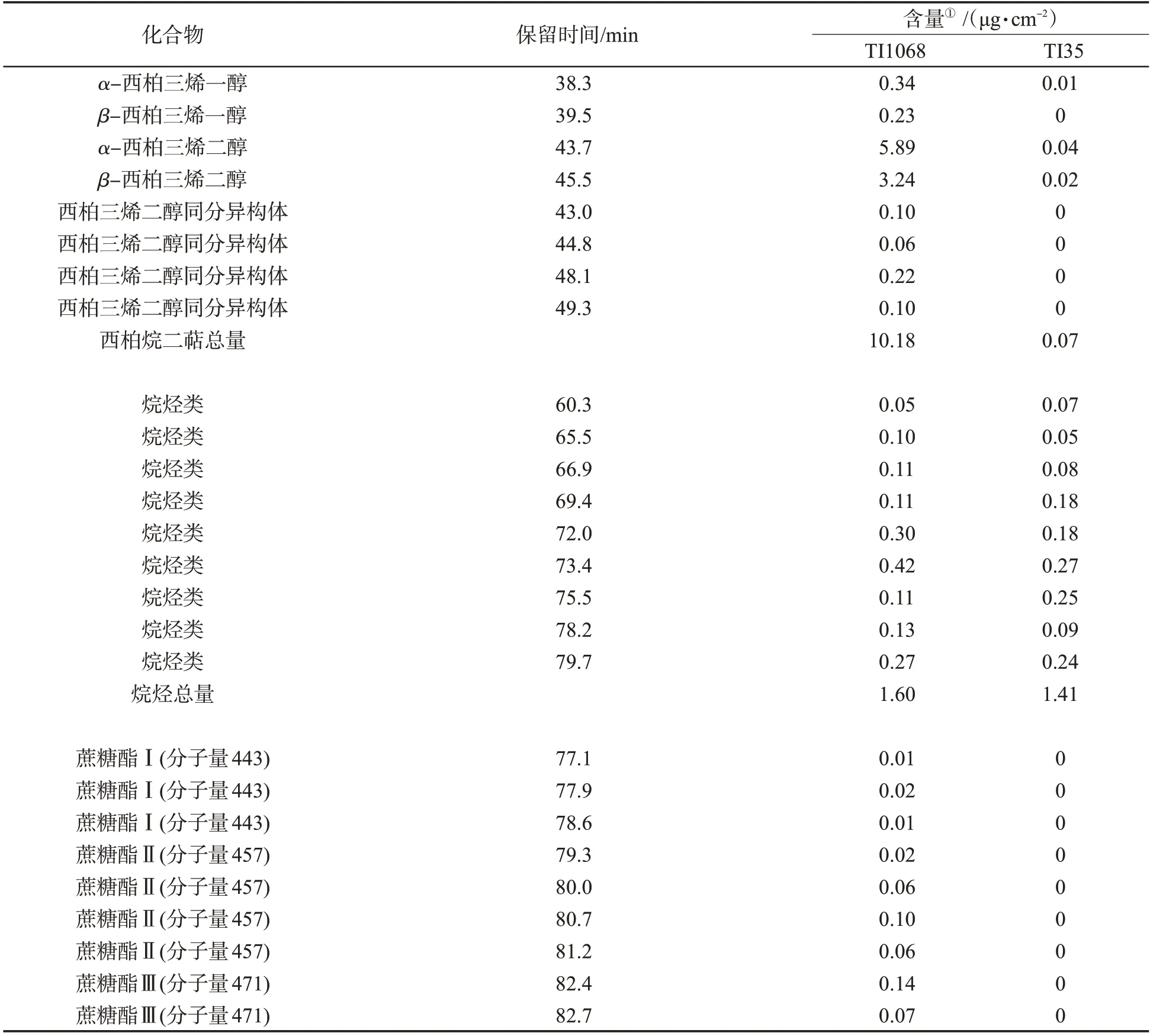
表1 煙草TI1068和TI35葉面化學成分比較Tab.1 Comparison of leaf surface chemical components between TI1068 and TI35

表1 (續)
2.3 煙草西柏烷二萜合成基因的表達分析
牻牛兒基牻牛兒基焦磷酸合成酶(Geranylgeranyl pyrophosphate synthase,GGPPS)是合成牻牛兒基牻牛兒基焦磷酸(GGPP)的關鍵酶[23],GGPP合成西柏烷二萜分為兩步:首先,GGPP在西柏三烯醇合酶(cembratrienol synthase,CBTS)的催化下發生環化反應,形成α-西柏三烯一醇和β-西柏三烯一醇;然后,西柏三烯一醇在細胞色素CYP450加氧酶(Cytochrome P450 hydroxylase,CYP450)的催化作用下,第6位碳發生羥化反應形成α-西柏三烯二醇和β-西柏三烯二醇,該酶的編碼基因為CYP71D16[24]。
采用半定量RT-PCR方法分析TI1068和TI35中西柏烷二萜合成基因的表達量結果見圖3。從圖3中可以看出,GGPPS、CBTS和CYP71D16在TI1068中的表達水平均高于TI35。這與表皮毛的形態比較和葉面化學成分分析結果一致,即長柄腺毛是西柏烷二萜合成的主要場所,TI1068具有高密度的長柄腺毛,而TI35僅有微量的長柄腺毛。

圖3 煙草TI1068和TI35西柏烷二萜合成基因的表達分析Fig.3 Expression analysis of genes related to diterpenoid biosynthesis in TI1068 and TI35
2.4 蚜蟲嗜好性比較

圖4 煙草TI1068和TI35的蚜蟲嗜好性比較Fig.4 Comparison of aphid preference between TI1068 and TI35
TI1068和TI35離體葉片上的蚜蟲數量見圖4。在各檢測時段TI35葉片的蚜蟲數量是TI1068的4~6倍。有研究表明,煙草腺毛可以特異地合成西柏烷二萜化合物,該化合物不僅具有抗真菌和抗蚜蟲活性,還可作為活性劑刺激煙草夜蛾產卵,其對于幼蟲的生長具有毒害作用[25-26];提高煙草腺毛中西柏三烯一醇含量可提高煙草的抗蚜蟲能力[27-28]。本研究中TI1068的蚜蟲嗜好性極顯著低于TI35,其原因可能是由于TI1068中西柏三烯一醇含量是TI35的57倍。
3 結論
①TI1068和TI35均有長柄腺毛、短柄腺毛和保護毛3種表皮毛類型;TI1068以長柄分泌型腺毛為主,含有少量保護毛,TI35以非分泌型表皮毛為主,含有微量的長柄腺毛;TI1068和TI35的短柄腺毛數量無顯著差異。②煙草TI1068和TI35葉面化學成分中均有西柏烷二萜、蔗糖酯和烷烴;TI1068葉面上3種物質含量約為TI35的10倍;TI1068葉面主要化學成分是西柏烷二萜和蔗糖酯類物質,而TI35主要是烷烴類,僅存在微量西柏烷二萜和糖酯Ⅳ。③西柏烷二萜合成基因GGPPS、CBTS和CYP71D16在TI1068中表達水平均高于TI35。④蚜蟲對TI1068的嗜好性極顯著低于TI35。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
