長白落葉松HD-Zip基因對外源激素的響應模式1)
2021-03-06 09:00:58呂思毓安旆其王朋陽劉牧田呂汐祁明惠張磊
東北林業大學學報 2021年2期
關鍵詞:植物
呂思毓 安旆其 王朋陽 劉牧田 呂汐 祁明惠 張磊
(林木遺傳育種國家重點實驗室(東北林業大學),哈爾濱,150040)
同源異型域—亮氨酸拉鏈(HD-Zip)蛋白屬于植物特有的轉錄因子,參與調控植物的生長發育過程,對于植物組織分化也有重要意義。HD-Zip包含一個由61個氨基酸構成的高度保守的同源異型域HD(Homeodomain,HD)和緊隨其后的亮氨酸拉鏈(Leucine zipper,LZ)結構域[1],共包括4個亞家族。
HD-Zip基因家族在林木的研究范圍非常廣泛,例如龍眼(拉丁名稱)體胚發生過程中鑒定得到的HD-Zip基因家族成員可能參與調控根的發育及果實的形成[2];白樺(拉丁名稱)在應答鹽脅迫的過程中有7條HD-Zip基因的表達量有變化[3];小黑楊(拉丁名稱)中鑒定得到的63條HD-Zip基因在鹽脅迫條件下葉中有25個HD-Zip基因下調表達,21個基因上調表達,莖中有42個基因下調表達,11個基因上調表達,根中有26個基因下調表達,24個基因上調表達[4];茶樹(拉丁名稱)的HD-Zip家族轉錄因子IV亞族的CsHB1基因在不同非生物脅迫處理下均能誘導表達,且表達差異性明顯,證實該基因家族可以參與光信號轉導、非生物脅迫響應、胚胎發育、分生組織形成等多種過程[5]。Soderma et al.[6]利用激素突變體進一步研究發現,ATHB6和ATHB7的表達主要依賴于ABA途徑在轉錄水平上調節的,同時它們在體內的活性還受到絲氨酸/蘇氨酸磷酸酶(ABI1和ABI2)的調控。Komatsuda T, et al.[7]發現HAHB10參與GA的轉導途徑;Himmelbach et al.[8]對模式植物擬南芥ATHB6的啟動子表達分析后,發現ATHB6啟動子的活性隨著外源ABA濃度的升高而上升。由此猜測ATHB6可能是與特異的ABA信號通路匹配的主要開關。
植物激素生理效應復雜多樣,可以從影響細胞分裂、伸長、分化到影響植物發芽、生根、開花、結實、性別決定、休眠、脫落等。吲哚—3—乙酸(IAA)是一種用作刺激植物生長的激素,對植物抽枝或芽、苗等的頂部芽端形成有促進作用[9];赤霉素(GA3)可以促進植物的生長發育,提早成熟[10],提高產量和打破休眠[7],促進發芽、分蘗、抽苔,提高果實結果率[11];脫落酸(ABA)是一種抑制生長的植物激素,它的作用與IAA等生長類激素相反,對細胞的分裂與伸長起抑制作用,還能抑制胚芽鞘、嫩枝、根和胚軸等器官的伸長生長[12],一般來說,干旱、寒冷、高溫、鹽漬和水澇等逆境都能使植物體內ABA迅速增加,同時抗逆性增強[13]。通過對長白落葉松HD-Zip基因響應IAA、GA3和ABA這3種外源激素處理的應答模式分析可以為該基因家族在針葉樹種的主要功能研究奠定基礎,同時為長白落葉松分子育種遺傳改良提供重要的基因資源。
1 材料與方法
1.1 長白落葉松HD-Zip基因
根據實驗室前期基因挖掘的數據,獲得2條HD-Zip家族基因,分別命名為LoHDZ1和LoHDZ3,其分別屬于HD-Zip I亞家族HD-Zip II亞家族。
1.2 落葉松幼苗
試驗所用的材料取自于黑龍江省雞西種源的長白落葉松種子。選取飽滿表面有光澤的長白落葉松種子,用去離子水浸泡約1周,期間換水3~4次,之后播種于V(豁土)∶V(蛭石)∶V(珍珠巖)=5∶3∶2的營養土基質(本試驗用到的基質都為按此比例配置的營養土)中,覆膜保濕,植物生長培養條件為16 h光照,8 h黑暗,濕度為75%,溫度恒定為22 ℃。
1.3 落葉松HD-Zip轉錄因子基因的生物信息學分析
利用BioEdit軟件對長白落葉松的HD-Zip轉錄因子蛋白序列進行比對分析,利用ExPaSy在線軟件(http://web/expasy/org/protparam/)對HD-Zip基因的蛋白分子質量和等電點等理化屬性進行分析。用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)對2條落葉松HD-Zip轉錄因子蛋白序列的二級結構進行預測分析,并用SWISS MODEL程序對上述2條蛋白序列進行三級結構同源建模。
1.4 外源激素處理
待落葉松的針葉完全伸開后挑取長勢良好的小苗移栽至養苗盆于營養土基質中繼續培養至2~3個月后,用濃度為50 mg/L的IAA、GA3和ABA溶液以及水(對照)噴施落葉松苗,并分別于2、4、8、12、24、48、72、96 h取整株落葉松苗提取RNA并反轉錄為cDNA。ABA,IAA,GA33種激素處理濃度參照欒嘉豫[14]等人的報告。
1.5 RNA提取及cDNA合成
本試驗采用PureLinkTMPlant RNA Reagent試劑盒提取激素處理后的長白落葉松試驗樣品RNA,提取步驟按照RNA提取試劑盒的方法進行;采用反轉錄Prime Script RT reagent Kit Perfect Real Time(TaKaRa)試劑盒將長白落葉松的試驗樣品進行反轉錄得到cDNA。
1.6 qRT-PCR
利用Primer5軟件設計基因定量引物(表1),利用EXCEL處理數據,分析HD-Zip基因的表達量。

表1 基因定量引物
2 結果與分析
2.1 落葉松HD-Zip轉錄因子基因的理化性質
將獲得的2條落葉松HD-Zip轉錄因子基因序列,利用軟件工具等進行分析的結果表明:LoHDZ1基因全長為1 057 bp,CDS區域為343 bp,相對分子質量為12 439.37,其蛋白等電點為7.04,蛋白脂肪系數為72.34,蛋白疏水性平均系數為-0.682。LoHDZ3基因全長為996 bp,CDS區域為291 bp,相對分子質量為37 507.45,蛋白的等電點為4.69屬酸性范圍內,蛋白脂肪系數為66.31。
兩條基因蛋白脂肪系數均高于50,表明其蛋白熱穩定性較高。LoHDZ1,LoHDZ3蛋白疏水性平均系數為-0.824,表現為親水性。信號肽分析和蛋白跨膜分析結果表明,LoHDZ1和LoHDZ3蛋白不具信號肽,均為非分泌蛋白及非跨膜蛋白(圖1)。

a為LoHDZ3蛋白質的氨基酸組成分析;b為LoHDZ3蛋白質的信號肽組成;c為LoHDZ3蛋白質三級結構;d為LoHDZ3蛋白質二級結構,其中紅色為螺旋,藍色為無規卷曲,白色為β轉角。
2.2 激素脅迫對長白落葉松HD-Zip轉錄因子基因表達影響
2.2.1GA3處理下的LoHDZ1、LoHDZ3的表達量
在長白落葉松幼苗施用GA3處理之后,LoHDZ1與LoHDZ3基因的表達量變化趨勢基本抑制,LoHDZ1基因表達量均明顯低于對照,LoHDZ3基因表達量也存在不同程度的下調表達,尤其是在2、4 h基因的表達量明顯低于未處理時期,說明噴施GA3后LoHDZ1和LoHDZ3基因活性受到抑制(表2)。

表2 LoHDZ1以及LoHDZ3在GA3處理下表達量
2.2.2IAA處理下的LoHDZ1、LoHDZ3的表達量
長白落葉松幼苗中的LoHDZ1基因表達量在生長素IAA處理后的2、4、12、48 h明顯上調表達,LoHDZ3在處理后12 h內表達量略有增加,48 h達到最大,約為未處理的10倍左右。兩條基因的表達量在72 h之后與未處理相比發生明顯下調(表3)。
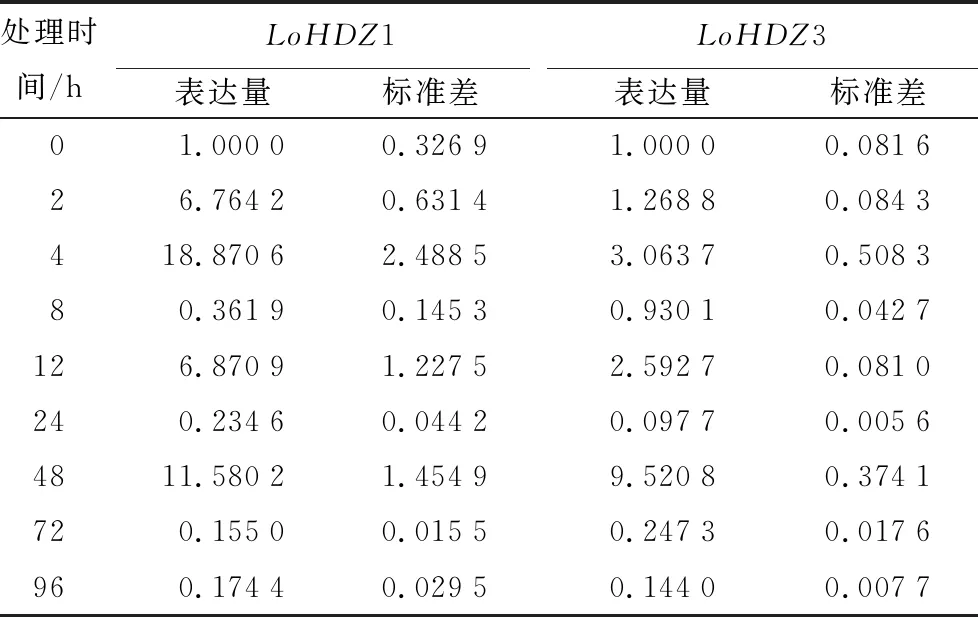
表3 LoHDZ1與LoHDZ3在IAA處理下表達量
2.2.3ABA處理下的LoHDZ1、LoHDZ3的表達量
經過ABA激素處理植株之后,LoHDZ1與LoHDZ3的基因表達量與未處理時相比大多為下調表達,LoHDZ1基因受到了極大程度的抑制,可推測該基因對于ABA激素的敏感度極高。即便是在處理后48 h表達量最大時也只有未處理時的1/5。LoHDZ3在2到12 h基因表達量明顯下降,在24 h時表達量為未處理時的1.5倍,72 h時表達量約為未處理時的1.3倍,可以初步推測LoHDZ3可以響應ABA處理,可能與植株抗逆性有一定關系(表4)。
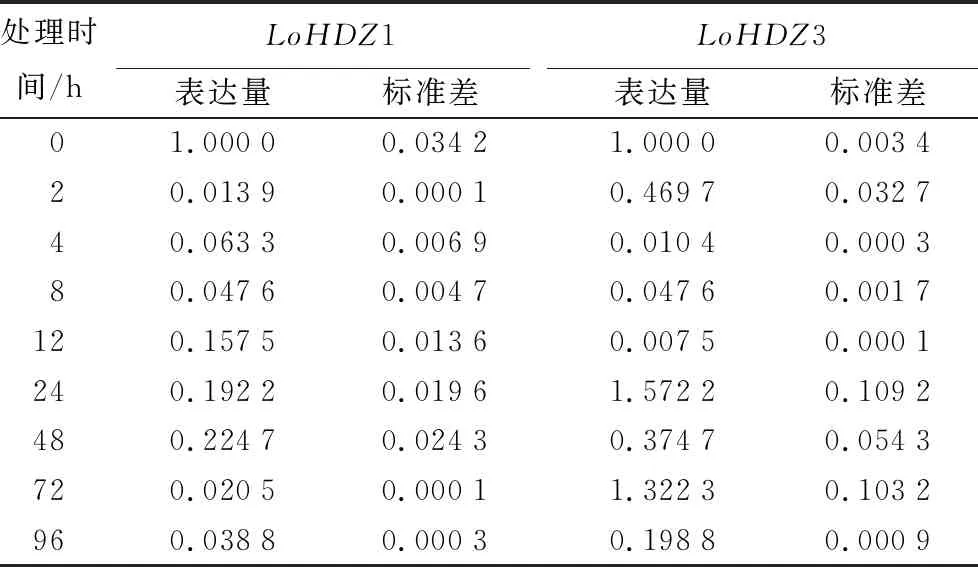
表4 LoHDZ1與LoHDZ3在ABA處理下表達量
3 結論與討論
經過3種不同類型的激素處理后,由于分別屬于HD-Zip I亞家族HD-Zip II亞家族,所以LoHDZ1與LoHDZ3基因的表達模式并不完全一致。從模式植物擬南芥以及水稻的研究來看,HD-Zip家族轉錄因子亞家族I和II基因表達受干旱、高鹽、ABA和冷害等環境的誘導,這兩類基因參與激素信號途徑,通過調控與激素途徑相關基因和下游基因互作來調控植物細胞擴增、分裂和分化,從而提高植物耐逆性[15],HD-Zip I亞家族主要在光信號轉導、非生物脅迫、葉片發育等方面發揮重要作用,而HD-Zip II蛋白的功能為介導植物對環境改變的應答并調節其發育過程[16]。HD-Zip I亞家族的相關研究表明其功能較為廣泛,例如番茄[17]、甘蔗[18]等植物中HD-Zip I亞家族基因在不同組織部位的表達特異性能證實該類基因可能參與調控植株不同組織器官生長發育的過程,特別是甘蔗GT1基因是HD-Zip I同源基因,在HD-Zip I家族蛋白的主要結構域上表現出高度的保守性,表明甘蔗GT1基因也可能參與了甘蔗關鍵發育過程的調控[18]。蘋果當中鑒定得到22條HD-ZipⅠ基因可以分別響應赤霉素、細胞分裂素、茉莉酸、乙烯和脫落酸的順式作用元件[19]。其中MdHZ1和MdHZ17可能與果實的后熟進程有較為緊密的聯系,可以作為候選基因進行果實后熟相關方面的深入研究。HD-Zip II轉錄因子控制激素調控發育的過程,缺失后將對植物生長素分布和響應產生影響[20]。例如油棕EgHOX1屬于HD-Zip II亞家族基因,其在胚胎發生誘導的早期組織培養階段檢測到高水平表達,但外源生長素對其誘導作用不明顯,而脫水和滲透脅迫對其誘導作用明顯[21]。桃樹中的HD-Zip II亞家族基因PpHB.G7在成熟果實中表達量高[22]。
在生長素IAA處理長白落葉松幼苗后,LoHDZ1的表達量在4 h內明顯上調表達,而LoHDZ3基因的表達量只是略有增加,說明其對生長類激素處理響應程度不同,LoHDZ1更容易被IAA調控表達。GA3處理后2條基因與對照相比表達量變化趨勢基本一致,因為赤霉素主要功能是促進植物生長,也能促進或控制內源生長素的合成與降解,避免生長素濃度過高及抑制時間過長致使植株生長減緩、停止甚至死亡。而ABA處理后LoHDZ1表達量較對照相比明顯下調,LoHDZ3在24和72 h時表達量升高,其余時期的表達量也均低于未處理時期。ABA對生長的作用與IAA、GA等相反,同時在植物對脅迫環境抗逆性中發揮重要作用。由此推斷LoHDZ1受到ABA的抑制,參與調控植物生長發育,不參與調控非生物脅迫過程,擬南芥第II亞類HD-Zip蛋白基因中有9個成員: ATHB2/HAT4,ATHB4,HAT1,HAT2,HAT3,HAT9,HAT14,HAT17和HAT22[23]。Steindler et al.[24]發現ATHB2的表達水平會影響生長素的信號途徑,表明其可能參與植物生長發育過程;過量表達擬南芥HAHB10后發現轉基因植株葉子的形狀、顏色,以及植株生長率、開花時間和生命周期都受到了影響,而用GA處理轉基因植株之后,植株形狀指標又會恢復原狀[25],表明HAHB10參與擬南芥生長發育過程。而LoHDZ3在用赤霉素處理后相對于對照組呈現明顯的差異性表達,推測其功能可能與大多數HD-Zip II亞家族基因一致,參與調控植物對脅迫的應答并調節生長發育部分過程。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
