典型喀斯特高原水庫浮游植物與環境因子的關系
2021-03-09 10:33:00葛秋詩張萍倪茂飛郭云王志康周慧張志威周業
生態環境學報 2021年1期
葛秋詩,張萍,倪茂飛,郭云,王志康,周慧,張志威,周業
貴州民族大學生態環境工程學院,貴州 貴陽 550025
浮游植物是水生生態系統的初級生產者,由于其群落結構對自身生存環境變化較為敏感,也可作為水環境特征的重要指示生物(胡芳等,2014;Lepisto et al.,2004;李然然等,2014)。因此,對水體浮游植物群落特征與環境因子間關系的研究十分必要。影響水體浮游植物密度及群落變化的環境因子具有時空差異,總體而言,水溫(tw)、透明度(SD)和營養物質作為自然水體生命活動的重要指標,通常與水生生物過程具有顯著的相關關系(鄧樂等,2019;鄒曦等,2017;邱小琮等,2012)。此外,其他因素如水利調節、化學計量比、溶解氧(DO)和pH等能夠顯示水生環境情況,因此可能極大地影響浮游植物密度與群落特征(李飛鵬等,2013;葛優等,2017;李磊等,2015)。
水庫是一種介于河流和湖泊間的半人工半自然水體,具有城市生活與工業供水等重要功能(林秋奇等,2001)。入庫水流在為水庫補給水源的同時,也為水庫提供營養來源。農業、城市和郊區徑流和生活廢水排放的營養鹽過量將導致浮游植物過量繁殖(楊朝霞等,2019;Howarth et al.,2002),從而對水庫水質及水廠后續處理產生惡劣影響。貴州省受喀斯特地貌影響,一方面,該地區受地質條件影響天然湖泊較少,需依靠人工建造的水庫以滿足城市生產生活用水(晏妮,2006),因而水庫水質與居民生活質量息息相關。另一方面,該地區水質具有偏堿性、鈣鎂含量高的特點(高廷進等,2014),白寧靜等(2019)研究認為,鈣離子是云貴高原地區湖泊中浮游植物群落構建的重要影響因子,說明喀斯特地區水體具有更加獨特的浮游植物群落特征。綜上所述,針對該地區水庫浮游植物群落特征與其相應的環境因子進行研究,具有十分重要的意義。
本文以典型喀斯特水庫——阿哈水庫為研究對象,通過對入庫支流、庫中心和南郊各采樣點不同季節(春、夏和秋季)的水體進行分析,探索了喀斯特水庫基礎水環境指標的量級、浮游植物群落組成、優勢種變化及其密度的時空分布,進一步揭示浮游植物與環境因子的耦合關系,為喀斯特高原水庫浮游植物研究以及水資源保護提供一定的科學依據。
1 材料與方法
1.1 采樣地概況及采樣點設置
阿哈水庫(106°39′E,26°33′N)位于長江流域烏江水系,屬于多入庫河流型水庫(高廷進等,2014),位于貴陽市南郊,由小車河上游的游魚河、白巖河、金鐘河、爛泥河以及蔡沖溝交匯而成。水庫匯水面積190 km2,正常蓄水位1110 m,對應水庫容積5.35×107m3,平均水深19 m;多年平均徑流量 3.24 m3?s?1,年均水輸送量 1.04×108m3。年均氣溫為15.3 ℃(萬國江等,2001),是貴陽市主要飲用水源地之一。
本研究在湖庫內共設7個采樣點,分別為游魚河(YY)、蔡沖溝(CC)、爛泥河(LN)、南郊水廠取水口(NJ)、水庫中心(KZ)、白巖河(BY)、金鐘河(JZ),分上、中、下三層取樣,其中蔡沖溝、爛泥河處較淺,取上下兩層。全年春、夏、秋三季共計取水樣 57份,冬季喀斯特地區浮游植物密度較少,且與秋季同屬枯水期(羅宜富等,2017),水文條件穩定,所以未將冬季納入研究范圍。采樣點的分布見圖1。
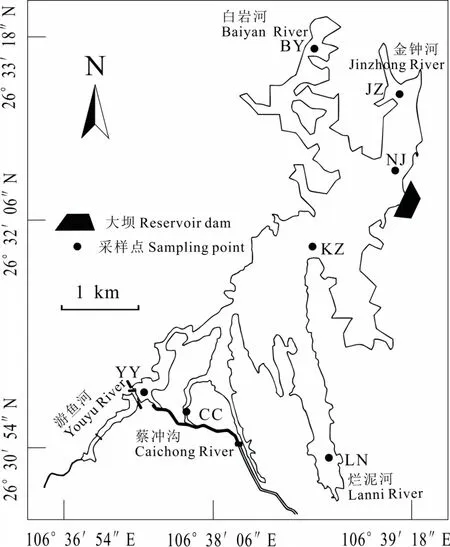
圖1 阿哈水庫采樣點分布Fig.1 Location of sampling sites in Aha Reservoir
1.2 樣品采集、處理與分析
1.2.1 野外采樣
樣品采集工作分別于2019年4、7、10月開展,共設置7個采樣點包含水庫主體及各支流,保證其地理位置具有空間代表性。3次采樣時間均為0:900—11:00,以避免浮游植物隨光照變化造成垂直移動現象。利用5 L有機玻璃采水器分3層(表層:0.5 m;中層:3 m;下層:5—8 m)取各點水樣,其中取1 L用于測定理化指標,另取500 mL現場加入7 mL左右魯哥氏液固定,利用25號浮游生物網于表層至0.5 m水深處作“∞”形緩慢循環拖動1—3 min,收集水樣并加入福爾馬林固定液使其濃度約為4%,作為浮游植物定性樣品。用于測定理化指標的水樣低溫運送回實驗室后,立即放入冰箱4 ℃保存并盡快測定,其余浮游植物指標水樣常溫下保存并及時測定。
1.2.2 樣品預處理
浮游植物定量樣品在實驗室靜置24 h,吸去上清液,并濃縮定容至30 mL定量瓶。將樣品充分搖勻后,吸取0.1 mL至計數框,利用400倍顯微鏡對浮游植物進行定量,每份樣品觀測兩次,結果取平均值。
1.2.3 實驗室分析
水庫tw和DO利用便攜式溶解氧儀(JPB-607A,雷磁,上海)進行測定,其中DO飽和度計算公式為:DO飽和度(%)=(溶解氧實測含量/實測條件下溶解氧的飽和含量)×100%,pH 利用便攜式 pH 計(PHB-4,雷磁,上海)測定,SD利用塞氏盤現場測定。總磷(TP)采用鉬酸銨分光光度法、鈣離子(Ca2+)濃度采用EDTA滴定法測定,方法參照《水和廢水分析監測方法》(國家環境保護局,1997)。總氮(TN)采用總有機碳分析儀(Vario TOC,Elementar,德國)的總氮(TNb)分析模塊測定,氨態氮(NH3-N)利用水楊酸鈉法通過全自動間斷化學分析儀(Cleverchem380,Dechem-Tech,德國)測定。氮磷比(N/P)為TN與TP的質量比值。浮游植物定性與定量分析依照《水和廢水分析監測方法》與《中國淡水藻類》(胡鴻鈞等,2006)所述方法進行。
1.3 統計分析
對不同季節測定的8種環境因子(tw、DO、SD、pH、TN、TP、Ca2+和NH3-N)與浮游植物密度(D)結果進行分析。浮游植物與環境因子的季節差異通過單因素方差分析(Analysis of Variance,One-way ANOVA)判斷;浮游植物密度與環境因子間關系通過一元線性回歸分析(Linear Regression Analysis)反映。
利用 Canoco 5軟件對浮游植物與環境因子進行排序分析,浮游植物優勢種類的篩選標準應滿足:該物種在各點出現的頻率>12.5%,且至少在一個點的相對密度>1%(Lopes et al.,2005;Muylaert et al.,2000)。首先,將除pH外的環境因子及浮游植物數據進行lg(x+1)轉換,并對浮游植物密度數據進行去趨勢對應分析(Detrended Correspondence Analysis,DCA)。DCA結果顯示,春、夏、秋的最大長度梯度依次為0.84、0.27、0.62,均小于3,因此采用基于線性的冗余分析(Redundancy Analysis,RDA)分析浮游植物與環境因子間的關系較為合適。
One-way ANOVA由IBM SPSS Statistics 20軟件完成,線性回歸分析及繪圖由Origin 2018軟件實現。
2 結果與分析
2.1 水體基礎環境因子
如表1所示,tw、DO、pH、SD、TN、NH3-N、和 Ca2+具有顯著的季節差異(One-way ANOVA,P<0.01),而各季節TP和N/P不具有顯著的統計性差異(P>0.05)。水庫tw在夏季最高為 (26.66±1.62) ℃,秋季最低 (17.00±0.84) ℃。DO為4.50—18.8 mg?L?1,且春季 (13.54±3.28) mg?L?1顯著高于其他季節[夏季(9.45±1.71) mg?L?1,秋 季(6.05±1.13) mg?L?1],春季水體 DO 飽和度最高(149%±34%),夏季次之 (118%±23%),秋季呈現缺氧狀態,飽和度最低 (63%±12%),這可能與浮游植物的光合作用有關。水體pH整體呈堿性,且在7.10—8.71范圍內。由于水體浮游植物活動頻繁,導致夏季SD值 (0.91±0.37) m低于春季 (1.08±0.49) m和秋季 (2.50±0.29) m。TN、NH3-N作為浮游植物的營養鹽,TN 夏季最高 (3.36±0.88) mg?L?1,NH3-N春季最高 (1.382±0.451) mg?L?1,均在秋季最低[TN:(2.09±0.21) mg?L?1,NH3-N:(0.055±0.042)mg?L?1]。受喀斯特地貌影響,Ca2+在 61.16—140.92 mg?L?1之間,其中秋季最高 (92.17±6.04) mg?L?1,夏季最低 (74.02±11.22) mg?L?1。

表1 各季節阿哈水庫的環境因子Table 1 Environmental indicators of Aha Reservoir in different seasons
空間上,春、夏季的營養鹽濃度(TN、TP、NH3-N)以游魚河最低,金鐘河最高,而Ca2+濃度則與之相反,為金鐘河低、游魚河高。秋季的金鐘河和南郊取水口TN、TP略高于其他位點,游魚河的NH3-N雖然高于其他位點的2—2.8倍,但僅為0.181 mg?L?1,Ca2+濃度各點差距較小,濃度最大的位點仍為游魚河(101.08 mg?L?1),即使是濃度最低的白巖河也達到 81.84 mg?L?1。總體而言,游魚河輸入營養鹽的量最少,金鐘河最多,其他各點對水庫的貢獻介于兩者之間,且由于金鐘河與南郊取水口距離最近,對水廠水質的影響不容忽視。
2.2 浮游植物群落組成及優勢種變化
通過對定性樣品的鏡檢,共鑒出8門80屬149種浮游植物,綠藻67種,為數量最多,硅藻42種次之,藍藻24種,甲藻5種,金藻4種,隱藻、裸藻各3種,黃藻1種。其中,不同時期的藍藻密度均占比最大,為64.7%—99.1%,其次是綠藻、硅藻、隱藻,密度占比分別為0.2%—22.8%、0.2%—11.0%、0—10.9%。金藻僅在春季具有數量優勢,最大密度占比為4.8%,甲藻、裸藻、黃藻數量較少,密度占比小于0.8%。
根據浮游植物出現頻率>12.5%和相對密度>1%的原則,篩選了22種浮游植物優勢種見表2。偽魚腥藻Pseudoanabaenasp.、水華束絲藻Aphanizomenonflos-aquae、曲殼藻Achnanthessp.為三季優勢種,三季各點出現頻率均為 100%;粗壯細鞘絲藻Leptolyngbyavalderiana為春、夏季優勢種,春、夏各點出現頻率分別為89%、100%;尖尾藍隱藻Chroomonasacuta、卵形隱藻Cryptomonas ovata和微球衣藻Chlamydomonasmicrosphaera為春、秋季優勢種,尖尾藍隱藻和卵形隱藻在春、秋各點出現頻率均為 100%,微球衣藻在春、秋各點出現頻率分別為 84%、95%;狹形纖維藻Ankistrodesmusangustus為夏、秋季優勢種,出現頻率均為100%;華美色金藻Chromulinaelegans、湖北小環藻Cyclotellahubeiana、尖針桿藻Synedra acus、庫津新月藻Closteriumkuetzingii為春季優勢種,出現頻率依次為89%、84%、37%、100%;兩頭針桿藻Synedraamphicephala為夏季優勢種,出現頻率為 100%;銅綠微囊藻Microcystis aeruginosa、點形平裂藻Merismopediapunctata、旋折平裂藻Merismopediaconvoluta、小空星藻Coelastrummicroporum、柱孢藻Cylindrospermumsp.、小球藻Chlorellavulgaris、嚙蝕隱藻Cryptomonaserosa、卷曲纖維藻Ankistrodesmus convolutus、顫絲藻Ulotrixoscillatoria為秋季優勢種,出現頻率依次為32%、68%、79%、79%、95%、95%、100%、100%、100%。

表2 阿哈水庫主要浮游植物優勢種名錄Table 2 Main phytoplankton species in Aha Reservoir
春季,華美色金藻、湖北小環藻及尖針桿藻在游魚河、蔡沖溝兩處的密度高于其他位點,而南郊取水口、庫中心、白巖河、爛泥河及金鐘河處的浮游植物以粗壯細鞘絲藻、偽魚腥藻、水華束絲藻、曲殼藻、尖尾藍隱藻、卵形隱藻微球衣藻和庫津新月藻為主;夏季蔡沖溝處為粗壯細鞘絲藻密度最大,游魚河、白巖河以兩頭針桿藻、曲殼藻為主,爛泥河、南郊取水口、庫中心、金鐘河以狹形纖維藻、偽魚腥藻、水華束絲藻、曲殼藻為主要種類;秋季各采樣點優勢浮游植物種類最多,不同采樣點的各類優勢種密度差別不大,可能是秋季水庫整體水質良好且混合均勻的緣故。
2.3 浮游植物密度時空分布
如圖2所示,浮游植物密度在時間尺度上存在顯著差異(One-way ANOVA,n=57,P<0.01),春、夏、秋季的浮游植物密度依次為2.55—21.10×105、3.13—43.92×105、6.79—17.45×104cells?L?1,呈夏季>春季>秋季的趨勢。春季浮游植物密度的最大點為南郊取水口(2.11×106cells·L?1),白巖河次之( 1.44×106cells?L?1),游魚河最低( 2.55×105cells?L?1);夏季浮游植物密度的最大點為庫中心(4.39×106cells?L?1),南郊取水口次之(3.38×106cells?L?1),蔡沖溝最低(3.13×105cells?L?1);秋季浮游植物密度的最大點為爛泥河(1.74×105cells?L?1),蔡沖溝次之(1.05×105cells?L?1),南郊取水口最低(6.79×104cells?L?1)。由此表明,春、夏兩季水庫中心區域易發生藻華。
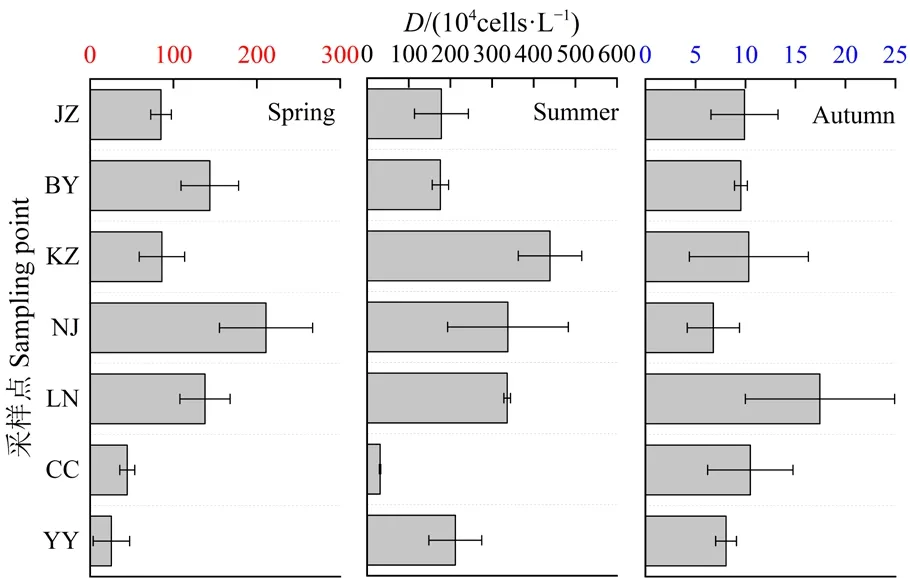
圖2 浮游植物密度(D)的時空分布Fig.2 Temporal and spatial distribution of phytoplankton density (D)
2.4 浮游植物密度與環境因子的線性回歸分析
不同季節浮游植物密度與環境因子間的線性回歸分析如圖3所示。春季浮游植物密度與眾多環境因子具有顯著相關關系,其與Ca2+濃度(r2=0.41,P<0.01)、N/P(r2=0.36,P<0.01)、pH(r2=0.26,P<0.05)以及 DO(r2=0.25,P<0.05)呈顯著的正相關,而與 SD(r2=0.38,P<0.01)呈顯著的負相關。夏季浮游植物密度僅與SD(r2=0.22,P<0.05)相關性較強,秋季浮游植物密度與所測環境因子均無顯著相關關系(P>0.05)。
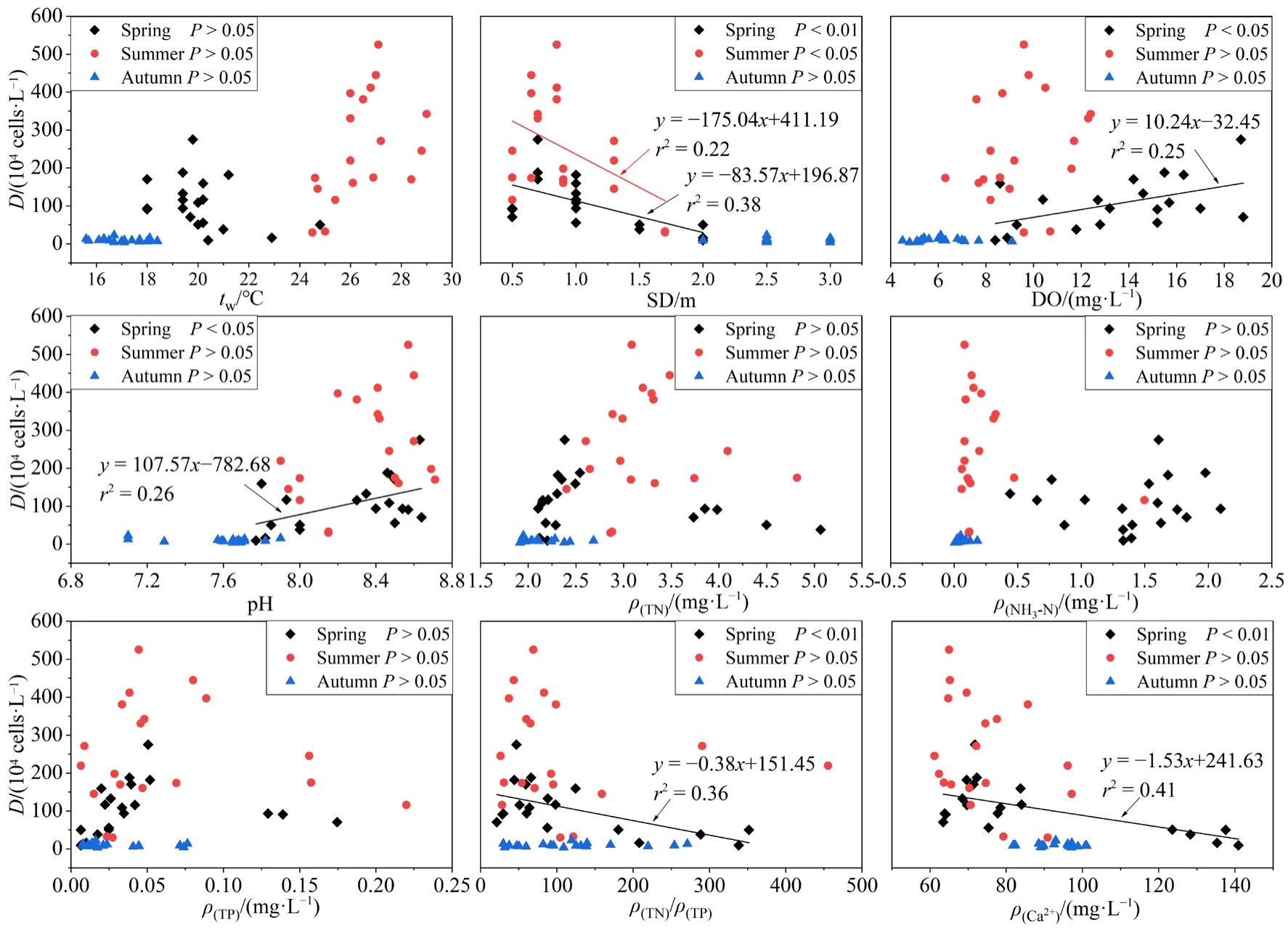
圖3 浮游植物密度(D)與環境因子之間的關系Fig.3 Relationship between phytoplankton density (D) and environmental factors
2.5 浮游植物與環境因子的RDA分析
RDA分析結果見表3及圖4,除秋季無環境因子入選外,春季軸Ⅰ和軸Ⅱ的特征值分別為 0.488和 0.1855,夏季軸Ⅰ和軸Ⅱ的特征值分別為0.445和0.1388,入選環境因子共解釋了春季76%、夏季68%的物種變化信息。前兩軸累計解釋了春季67.35%、夏季 58.38%的物種變化信息和春季88.7%、夏季 85.9%的物種-環境關系信息;春、夏季的兩個物種排序軸近似垂直,相關系數分別為?0.0451、?0.0149,環境因子軸與物種排序軸之間的相關系數春季為0.9672、0.8536,夏季為0.9527、0.8388,春夏季兩個環境排序軸的相關系數均為0,兩季節的Monte Carlo置換檢驗所有排序軸均達到顯著水平(P<0.05),說明排序結果可信(Ter Braak et al.,1988),排序軸與環境因子間的線性結合程度較好地反映了浮游植物與環境之間的關系。

表3 浮游植物與環境因子的RDA統計結果Table 3 Redundancy analysis results of phytoplankton and environmental factors
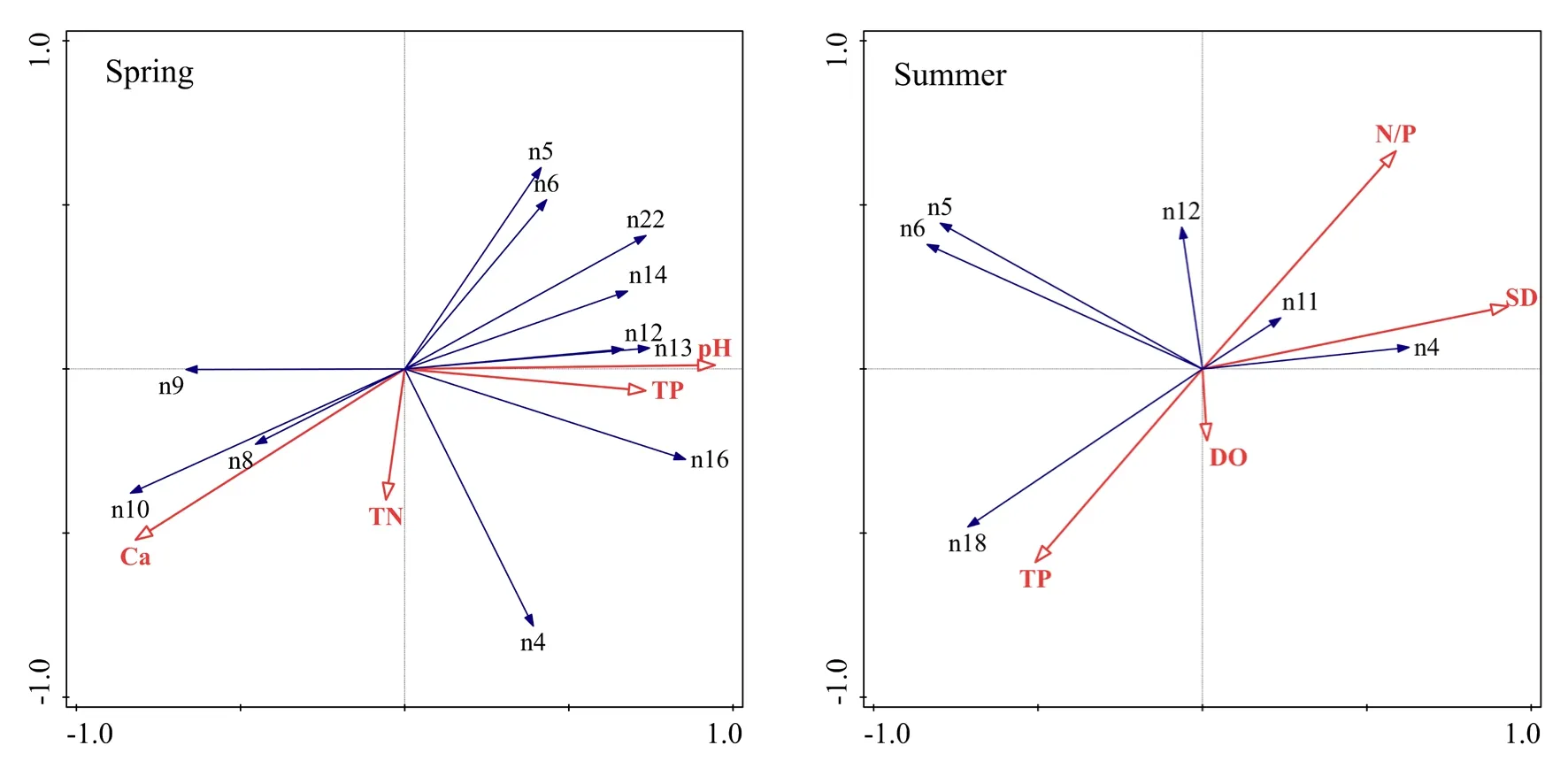
圖4 不同季節浮游植物-環境因子關系的RDA分析Fig.4 RDA of phytoplankton environmental factors in different seasons
利用向前引入法(Forward Selection)對環境因子進行篩選,將Monte Carlo置換檢驗結果為P<0.05的環境因子作為該季節的主要影響因子參與 RDA分析,春季的主要影響因子為pH、Ca2+濃度、TP、TN,夏季為SD、DO、N/P、TP,秋季浮游植物優勢種類組成較復雜,未有環境因子入選(P>0.05)。春季影響浮游植物分布的環境因子中,pH、Ca2+濃度、TP為第Ⅰ軸的主要因子,TN為第Ⅱ軸的主要因子。多數優勢種與pH、TP呈正相關,與Ca2+濃度呈負相關;其余優勢種,如華美色金藻(n8)、湖北小環藻(n9)、尖針桿藻(n10)與 Ca2+濃度呈正相關,與 pH、TP呈負相關;粗壯細鞘絲藻(n4)、華美色金藻(n8)、尖針桿藻(n10)與TN呈正相關,偽魚腥藻(n5)、水華束絲藻(n6)、卵形隱藻(n14)、庫津新月藻(n22)與TN呈負相關。夏季影響浮游植物分布的環境因子中,SD為第Ⅰ軸的主要因子,TP、N/P、DO為第Ⅱ軸的主要因子。粗壯細鞘絲藻(n4)、兩頭針桿藻(n11)與SD呈正相關,偽魚腥藻(n5)、水華束絲藻(n6)、狹形纖維藻(n18)與SD呈負相關;粗壯細鞘絲藻(n4)、兩頭針桿藻(n11)、曲殼藻(n12)與N/P呈正相關,與TP呈負相關;狹形纖維藻(n18)與 TP呈正相關,與 N/P呈負相關;狹形纖維藻(n18)與DO呈正相關,偽魚腥藻(n5)、水華束絲藻(n6)、兩頭針桿藻(n11)、曲殼藻(n12)與DO呈負相關。
總體而言,影響春季浮游植物群落結構的主要環境因子是pH、Ca2+濃度和TP,而夏季的主要環境因子為SD、TP和N/P。
3 討論
3.1 阿哈水庫浮游植物群落特征
采樣期間阿哈水庫浮游植物密度在 3.95—525.35×104cells?L?1之間(圖 2),春、夏季均為藻華暴發的高風險時期,浮游植物密度可達 106cells?L?1數量級,采樣期間浮游植物數量以藍藻、硅藻、綠藻、隱藻居多,屬于富營養型湖泊(龐清江等,2003)。另外,金藻也是春季的優勢種之一,在游魚河處密度最大,占總密度的1.88%,遠高于其他點位(<0.7%),金藻常生長于溫度較低、有機質含量少的清潔水體(胡鴻鈞等,2006),表明游魚河區域水體營養程度相對較低。與 2012—2013年對阿哈水庫浮游植物群落的調查結果(羅宜富等,2017)相比,甲藻與微囊藻數量均明顯減少,偽魚腥藻、水華束絲藻等絲狀藍藻成為水庫最大優勢種。
3.2 浮游植物密度與環境因子間的關系
根據線性回歸可知(圖3),影響浮游植物密度的環境因子,春季為 SD、DO、pH、Ca2+濃度和N/P,夏季為 SD。SD由水體泥沙顆粒物與浮游植物含量共同決定,SD隨著浮游植物密度的增長而降低(P<0.05),說明SD很大程度上是由浮游植物密度決定的,因此將SD看作浮游植物的被動影響因子。春季DO出現過飽和狀態且隨浮游植物密度的增加而升高(P<0.05),說明浮游植物光合作用釋放氧氣是水體DO偏高的重要因素之一,同時,浮游植物活動也將引起pH升高(Ouyang et al.,2018),再次驗證了春季浮游植物與 SD、DO、pH之間存在密切聯系。阿哈水庫Ca2+濃度偏高除了受喀斯特地貌的影響外,還有部分人為因素,游魚河、蔡沖溝、白巖河是煤礦廢水流入阿哈水庫的主要通道,酸性pH及鐵、錳等元素也會對水庫生態系統造成影響,需常在三處投放大量石灰進行治理(白薇揚等,2007),使得春季游魚河、蔡沖溝 Ca2+濃度高達140 mg?L?1左右,這勢必會對浮游植物生長造成影響。一方面,Ca2+可與磷形成共沉淀,從而減少水體磷含量(劉冠男等,2011),間接抑制浮游植物生長;另一方面,藍藻大量繁殖時會增強對Ca2+的吸收利用(史綿紅等,2009),使其濃度減小,因此春季浮游植物密度隨 Ca2+濃度增加而顯著減少(P<0.01)。營養鹽也是促進浮游植物生長的重要因素,根據羅宜富等(2017)的研究結果,阿哈水庫屬于磷限制水體,低N/P可能是內源或外源排放的磷濃度增加造成的,亦可能是藍藻水華導致氮濃度下降、磷濃度升高的結果(許海等,2011),從而使春季浮游植物密度隨 N/P增加呈現下降趨勢(P<0.05)。秋季浮游植物密度較低且各點位浮游植物組成無明顯差異,主要是整體tw、營養鹽濃度偏低導致。此外,采樣期間水庫大壩開閘放水,水流速率較大,也妨礙了浮游植物的正常生長,使其密度降低。
3.3 浮游植物群落結構與環境因子間的關系
根據RDA可知(圖4),影響春季浮游植物群落結構的主要環境因子是TP、pH和Ca2+濃度,而夏季的主要環境因子為TP、N/P和SD。高TP濃度的水體中,藍藻、綠藻、隱藻更易成為優勢種,金藻、硅藻更傾向生存于低TP濃度水體。高濃度Ca2+可抑制沉積物中的磷向水中釋放(劉冠男等,2011),間接抑制了藍藻、綠藻、隱藻的生長,促進了金藻和硅藻的繁殖。研究表明,大多數藍藻的最適pH在7—9之間,綠藻為7—10(許海等,2009),本研究結果顯示,藍藻、綠藻、隱藻更適于pH較高的水體生長,同時,浮游植物的光合作用也會導致pH進一步升高,從而鞏固其生長優勢。偽魚腥藻、水華束絲藻(藍藻)以及狹形纖維藻(綠藻)數量隨N/P的增加而減小,而兩頭針桿藻和曲殼藻(硅藻)與 N/P呈正相關,這是因為藍藻的生長環境需要高磷低氮,綠藻也是在高磷水體中更具優勢,而硅藻則傾向于高氮低磷的水體(石彭靈,2015;吳雅麗等,2013)。藍藻可通過偽空胞的組裝與破裂、碳水化合物的合成與消耗來調節其在水體中的垂直位置(Wallace et al.,2000;Pinilla,2006),因此SD降低使得藍藻上浮至水體表層,對中下層其他浮游植物生長造成影響,進而使藍藻成為夏季密度最大的優勢種。秋季水體優勢浮游植物組成最豐富,含有藍藻、綠藻、硅藻、隱藻共 16種浮游植物優勢種。藍藻的大量繁殖會分泌化感物質抑制其他藻類的生長(楊斌等,2007),其最適生長溫度為 25—30 ℃(Yamaguchi et al.,2000),秋季水溫均值僅為 17 ℃,在一定程度上抑制了藍藻的生長,為其他藻類提供生存機會。
4 總結
本研究通過對阿哈水庫3個季節的監測表明,春、夏季均為藻華暴發的高風險時期,浮游植物密度在 3.95—525.35×104cells?L?1之間,采樣期間浮游植物數量以藍藻、硅藻、綠藻、隱藻居多,絲狀藍藻(如偽魚腥藻、水華束絲藻)為水庫主要優勢種。各季節浮游植物與環境因子之間存在相互作用,春季與SD、DO、pH、Ca2+濃度和N/P關系密切,夏季僅與SD相關,秋季受水溫、營養鹽濃度以及水流速率影響較大。其中,pH、Ca2+和TP是影響春季浮游植物群落結構的主要環境因子,而夏季的主要環境因子為SD、TP和N/P,秋季受低溫影響較大。春季Ca2+濃度的變化對浮游植物密度及群落結構均有顯著影響。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
趣味(語文)(2020年3期)2020-07-27 01:42:46
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
